Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Research Article - (2015) Volume 4, Issue 5
The object of this study was to investigate the effect of serum antibody of peptide tyrosine tyrosine on activities of digestive enzymes in rats. A total of sixty sprague-dawley (SD) rats weighing 87.77 ± 1.61 g were randomly allocated to 1 of 4 dose treatment:0 (control), 50 μL, 100 μL, or 200 μL peptide tyrosine tyrosine antiserum during the 21-d experimental period. The activity of amylases and lipases in the pancreas and duodenum as well as gastric pepsin was measured at the end of the study. The results showed that the activity of lipases in the pancreas and duodenum increased with increasing peptide tyrosine tyrosine antiserum level, but obvious tendency of pancreatic lipases activity between all treatments was not observed(P>0.05). The lipase activity in the duodenum showed a significant increase (P<0.05) in the groups treated with 100 μL or 200 μL of peptide tyrosine tyrosine antiserum compared with the control group. The amylase activity in the pancreas and duodenum increased with the increasing peptide tyrosine tyrosine antiserum dose, and the group treated with 200 μL of peptide tyrosine tyrosine antiserum was significantly higher (P<0.05) than the other groups. The amylase activity in the duodenum showed no significant difference (P>0.05) among groups, but the group treated with 200 μL of PYY antiserum showed the highest activity. In conclusion, the injection of peptide tyrosine tyrosine antiserum improved nutrition utilization rate and activity of digestive enzymes in a dose dependent manner in rats.
<Keywords: Peptide tyrosine; Tyrosine; Antiserum; Digestive enzymes; Rat
The gut hormone peptide tyrosine tyrosine (PYY) is a 36-amino acid peptide and two main endogenous forms PYY1–36 and PYY3–36, have been identified, with PYY3–36 being the predominant circulating form [1-3]. Peptide tyrosine tyrosine is released from specialized enteroendocrine cells called L-cells, which are found predominantly within the distal GI tract [4,5]. PYY may be secreted by the stomach and pancreatic endocrine cells, while it is also found in the central nervous system (hypothalamus, brain stem, medulla, pons and spinal cord) and the peripheral nervous system neurons [6,7]. Peptide tyrosine tyrosine is secreted alongside hormone glucagon-like peptide 1, and it is released after eating, circulates in the blood and functions by binding to receptors in the brain. The binding of these receptors then causes a decreased appetite and makes people feel full after eating. Moreover, peptide tyrosine tyrosine had important effects on the function of the stomach and intestine to slow down the movement of food through the digestive tract.
Peptide tyrosine tyrosine exerts its action through neuropeptide Y (NPY) receptors to inhibit gastric motility and increase water and electrolyte absorption in the colon [8]. Expression of Y2 receptor is the most abundant PYY receptor on pancreatic cancer cell. This indicated that peptide tyrosine tyrosine may also affect pancreatic secretion [9]. In the pancreatic, PYY occurs in islet cells of the mouse pancreas, most of which are glucagon cells, and that PYY inhibits stimulated insulin and glucagon secretion in vivo in the mouse.[10]. This indicated that it played an important role in the proliferation and differentiation during the development of the pancreatic [7]. Peptide tyrosine tyrosine can regulate the function of digestive system by slowing the gastric emptying; hence, it increases efficiency of digestion and nutrient absorption after a meal. Peptide tyrosine tyrosine also plays a very important role in energy homeostasis by regulating food intake via the neuroendocrine system (brain-gut axis). Previous study indicated that PYY may affect directly the hypothalamic arcuate nucleus of its neurons after secreted by the intestinal endocrine cells by G protein-coupled way and have an ingibitory effect on the ingestion of animals; Or else PYY may affect the feed intake of animal in the way of negative feedback. That means the negative feedback of PYY can block the ventrolateral thalamic nucleus under the excitement and express of NPY caused by ingestion [1]. PYY-knockout rats, on the other hand, are resistant to obesity, but have higher fat mass and lower glucose tolerance when fed a high-fat diet, compared to control rats. High PYY concentrations cause a decrease in appetite and food intake. Moreover, high PYY concentrations are associated with diseases where there is dramatic weight loss, such as anorexia nervosa, coeliac disease, inflammatory bowel disease (Crohn’s disease and ulcerative colitis) and some cancers. Low PYY concentrations are associated with an increase in appetite and food intake. There has been some research into using PYY as a medication for obesity, aiming to decrease the appetite of people who are overweight.
PYY is the satiety factor physiologically inhibiting food intake in animals, and it regulates a variety of gastrointestinal hormones [11]. As shown by both in vitro and in vivo studies, PYY inhibits the secretion of insulin. PYY can also reduce gastric acid secretion and slow down gastric emptying. In addition, a relationship between PYY and regulating factors associated with feeding has been observed in animals. However, future investigation is needed. Moreover, active immunization against PYY decreases its level in serum, but the resulting effects on the hormones and enzymes related to feeding in animals still remain largely unexplored. Therefore, the objective of this study was to investigate effects of passive immunization against PYY on activities of digestive enzymesin rats.
Animal management
PYY antiserum was derived from rabbits and prepared before the experiment. Sprague–Dawley (SD) rats were obtained from the Laboratory Animal Center of the Chinese Academy of Sciences (Hunan, China). The rats were housed in a pathogen-free house and had free access to food and drinking water. Feed intake was recorded.
Study design
Sixty SD rats weighing about 70 g were randomly assigned to three active treated groups and one control group, with five replicates in each group and three rats for each replicate. The rats were injected with 50 (group 1), 100 (group 2), 200 (group 3) μL of PYY antiserum or 0.9% physiological saline (control) respectively. The rats were treated once a week for three weeks and the experiment lasted for 21 days.
Digestion experiment
The digestion experiment began on the day of 21. The digestion trial consisted of a 3-day adaptation period followed by a 7-day feces collection period. During the digestion trial, the rats had free access to food. The feces were collected twice daily (8 AM and 8 PM) and stored at -20°C. The feces were thawed, mixed well, dried in a 60-70°C oven, and ground to pass a 40 mesh screen for analysis of dry matter, crude protein, and gross energy.
Sample collection
At the end of the trial, one rat from each replicate was sacrificed, and the stomach, duodenum, pancreas were removed and weighed immediately. The samples were snap frozen in liquid nitrogen and stored at -20°C until further analysis.
Measurement of digestibility of nutrients
Measurement of original moisture: The original sample was dried in a 65°C oven for 8-12 hours, and the weight was taken after leaving the sample at room temperature for resurgence for 24 hours. After weighing the sample, it was dried in a 65°C oven for 2 hours and left at room temperature for resurgence for 24 hours before achieving the weight again. The procedure was repeated till the difference between two weights was less than 0.5 g, and the lowest value of the weights was used for the calculation of original moisture.
Original moisture = (initial weight-dried weight) / initial weight × 100%
Measurement of dry matter: The air-dried sample was dried in a 105 ± 2°C oven till the stable weight was achieved. The bound moisture was measured and the dry matter was calculated by the following formula. Determination of dry matter content of the drying method, reference GB/T 6433-2006 [12].
Water = (W1-W2) / (W1-W0) ×100%
*W1 (g) is the total weight of the sample and the container before the sample was dried in a 105±2°C oven; W2 (g) is the total weight of the sample and the container after the sample was dried in a 105±2? oven; W0 (g) is the weight of container.
Total water =[A+(1—A)×B]×100%
*A is the measured original moisture (%); B is the bound moisture of dried sample (%).
Measurement of crude protein: Crude protein were measured using FOSS Kjeltec 2300 (Foss, Denmark). Determination Kjeldahl method crude protein content, reference GB/T 6432-1994 [13].
Energy measurement: The Energy was measured by using HXR- 6000 precision automatic calorimeter.
Measurement of enzyme activity: The tissue of stomach, pancreas, or duodenum was homogenized in chilled 0.9% physiological saline to obtain a 10% (for pepsin and amylase) or 20% (for lipase) homogenate (w/v) which was then centrifuged at 2500 rpm for 10 min. The resulting supernatant was used for analysis of stomach pepsin activity with an ELISA assay kit (Nanjing Jiancheng Bioengineering Institute) according to the manufacturer’s instructions.
The lipase activity in the pancreas and duodenum was analyzed according to the Kaumas brilliant blue method using a lipase kit (Nanjing Jiancheng Bioengineering Institute) according to the manufacturer’s instructions
The amylase activity in the pancreas and duodenum was determined using an amylase assay kit (Nanjing Jiancheng Bioengineering Institute) according to the manufacturer’s instructions. The protein content in the tissue was also measured with an assay kit (Nanjing Jiancheng Bioengineering Institute) using Coomassie brilliant blue method.
Statistical analysis
Data were processed using Excel and analyzed by SPSS 11.5. Tukey test was used for multiple comparisons. The significance level was set at 0.05. The results were expressed as mean ± standard deviation (X ± SD).
Effects of PYY antiserum on nutrition utilization rate
As shown in Figure 1, the difference of utilization ratio of dry matter among groups was not significant (P>0.05), with the highest in the control group and the lowest in the group 2. Utilization ratio of crude protein was improved by 76.65 % (P<0.05) in group 3 and numerically increased (P>0.05) by 24.4% in group 1 and 38.13 % in group 2 when compared with the control. Utilization ratio of energy was enhanced (P<0.05) by 161.11% in group 3 and numerically increased (P>0.05) by 46.88% in group 1 and 19.90 % in group 2 relative to the control.
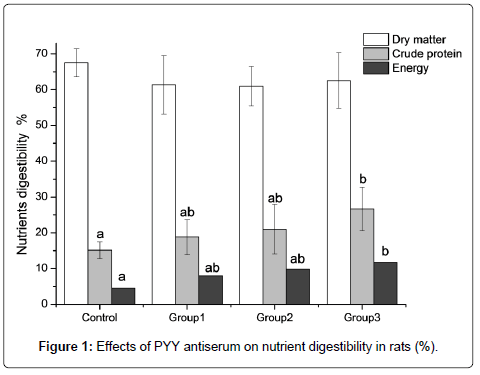
Figure 1: Effects of PYY antiserum on nutrient digestibility in rats (%).
Effects of PYY antiserum on activity of digestive enzymes
The activity of pancreatic lipases increased numerically with the increasing dose of PYY antiserum, but the difference among groups was not significant (P>0.05). Compared with the control group, pancreatic lipases activity increased by 7.80%, 27.96% and 38.58% in groups 1, 2, and 3, respectively. The activity of lipases in the duodenumincreased with the increasing dose of PYY antiserum, and group 3 had a significant (P<0.05) increase compared with the control and group 1. Lipase activity in the duodenum of group 3 increased by 14.31%, 10.21% and 2.37% compared to the control, group 1 and group 2, respectively. Group 2 had an 11.67% increase (P<0.05) of lipase activity in the duodenum compared with the control group (Figure 2).
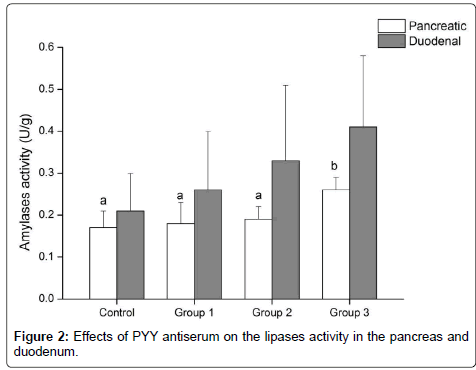
Figure 2: Effects of PYY antiserum on the lipases activity in the pancreas and duodenum.
As shown in Figure 3 the activity of amylases in the pancreas and duodenum increased as the dose of PYY antiserum increased. The activity of pancreatic amylases of group 3 increased (P<0.05) by 52.94%, 44.44% and 36.84% compared with the control, group 1 and group 2, respectively. The activity of amylases in the duodenum did not show a significant difference among groups (P>0.05), but group 3 had the highest activity, with an increase of 96.24%, 75.69% and 24.24% compared with the control, group 1 and group 2, respectively (Figure 3).
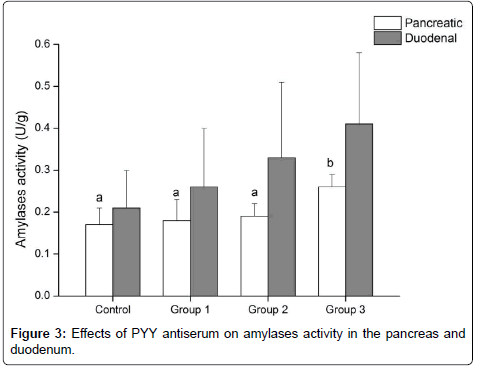
Figure 3: Effects of PYY antiserum on amylases activity in the pancreas and duodenum.
As shown in table 1, the activity of gastric pepsin increased as the dose of PYY antiserum increased, but the difference among groups was not significant (P>0.05). After the injection of PYY antiserum, the protein content in the pancreatic and gastric tissues was not significantly different among groups (P>0.05), and the group 3 has the lowest protein content in the pancreatic tissue. The difference of the protein content in the duodenum was significant among groups, with the control, groups 1 and 2 being 16.13%, 11.29% and 8.06%, respectively, lower (P<0.05) than group 3 (Figure 4).
| Pepsin | |
|---|---|
| Control group | 52.25 ± 14.02 |
| Group 1 | 58.12 ± 10.47 |
| Group 2 | 63.27 ± 10.64 |
| Group 3 | 65.26 ± 5.71 |
Table 1: Effects of PYY antiserum on pepsin activity (U/g).
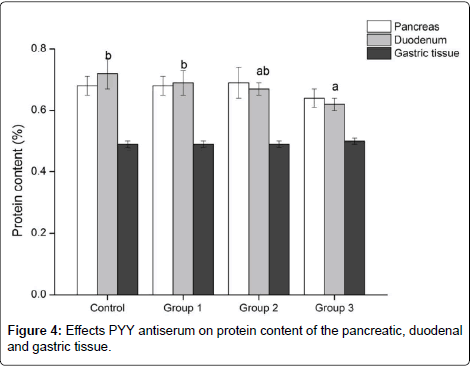
Figure 4: Effects PYY antiserum on protein content of the pancreatic, duodenal and gastric tissue.
Effects of PYY antiserum on nutrition utilization rate
PYY is released from the gut into the circulation in a nutrientdependent manner. PYY levels are low in the fasting state, rapidly increase in response to food intake, reach a peak at 1–2 h after a meal and then remain elevated for several hours [14]. PYY3–36 has additional metabolic beneficial effects on energy expenditure and fuel partitioning. PYY regulates body weight by reducing food intake and increasing energy expenditure [15]. Several studies suggested a role of endogenous PYY in satiety regulation in humans [5,16]. And some benefits were observed in experimental subjects in reducing hunger and promoting weight loss [2]. This would help explain the weight-loss experienced with high-protein diet. Blockade of endogenous PYY by administration of PYY antiserum resulted in increased food intake in rats that had undergone jejuno-intestinal bypass [17-18].
The effects of the injection of PYY antiserum on the growth and endocrine in rats have been proved to some degree, but the effect of PYY antiserum on the nutrient absorptivity in rats are not well studied [19]. Our study showed that dry matter digestibility was not affected by PYY in a dose dependent manner, and the difference among groups was not significant. Digestibility of crude protein increased with the increasing dose of PYY antiserum, and the difference between the treated groups and the control group was significant. Digestibility of energy also increased with the increasing dose of PYY antiserum in rats, which indicates that the injection of PYY antiserum affected the metabolic rate, facilitated the metabolism of food, and improved the growth of animals.
Effects of passive immunization against PYY on the digestive enzymes
Exocrine pancreas (mainly pancreatic juice) plays an important role in digestion and absorption of food. Irregular secretion of pancreatic juice leads to abnormal digestion and absorption, affects the growth of animals, and harms the production [20]. Pancreatic juice is maintained at a normal level under the neuronal and homeostatic regulation. It is the colorless and odorless alkaline solution, mainly including water, inorganic and organic chemicals [21]. The organic chemicals are mostly composed of multiple digestive enzymes, including pancreatic amylases, lipases, proteases and intestinal kinases. PYY regulates growth, digestion and absorption in the gut and pancreas, and its physiological function in the immunization and nutrient absorption are through decreasing cell factors and releasing amylases. Although the mechanism is still unclear, the glandular level of PYY is believed to be associated with many cell transcription factors. PYY stimulates the secretion of pancreatin, CCK and insulin, and inhibits the secretion of glucagon, somatostatin and pancreatic polypeptide [22]. PYY inhibits endocrine and exocrine secretion of the pancreas, reduces the pancreatic blood flow, suppresses the growth of the pancreas stimulated by pancreatin, and inhibits the basal pancreatic secretion and the secretion of pancreatic juice stimulated by vagus nerve [23]. As shown in Figures 2 and 3, the activity of pancreatic lipases, pancreatic amylases and duodenal amylases increased with the increasing dose of PYY antiserum, which is consistent with previous studies.
Pepsin hydrolyzes the protein in the food, and its primary role is to hydrolyze protein and peptide containing phenylalanine or tyrosine, and the decomposed products are mainly peptone, with polypeptide or amino acids as minor products. Pepsin only functions in the acidic environment, and the optimistic pH is 2. The activity of protease decreases as the pH increases, and the protease is irreversibly denatured if the pH rises to more than 6. Therefore, stomach proteases and stomach acid are closely related. Previous research has shown that intravenous injection of 50 pmol/kg/h of GLP-1 or PYY in dogs suppressed the secretion of stomach acid, and completely inhibited secretion of stomach acid if injected with both drugs. Studies also showed that taking 10 pmol/kg of PYY and 0.05 CU/kg of secretin hourly inhibited the secretion of stomach acid stimulated by pentagastrin in human [24-25].
PYY suppresses the secretion of stomach acid through the regulation of central nervous system. Research showed that PYY regulates the secretion of stomach acid via vagus nerve, and most of them are through vagus nerve at the pyloric part of stomach [26]. PYY and Y2 receptor agonists regulate the secretion of stomach acid via dorsal motor nucleus of vagus nerve in the in vitro and in vivo experiments [27,28].
As shown in Table 1 the activity of pepsin decreased with the increased dose of PYY antiserum. A possible mechanism could be that the effects of inhibiting the secretion of stomach acid by PYY reduced with the passive immunization of PYY, which caused the raising of pH in the stomach, and thus decreased the activity of pepsin. However, further investigation is still needed.
The injection of PYY antiserum affected digestibility of crude protein and energy, which increased with the increasing dose of PYY antiserum. However, PYY antiserum had no obvious effect on the digestion of dry matter. Injection of PYY antiserum improved the activity of pancreatic lipases, pancreatic amylases, duodenal lipases and duodenal amylases in a dose-response manner. But the activity of gastric pepsin decreased with the increasing dose of PYY antiserum. The injection of PYY antiserum had no effects on protein content in pancreas and gastric tissue, and the injection of high dose of PYY antiserum (200 μl/rat) decreased the protein content in duodenum.
Immunizing rats with the PYY antiserum could improve the nutrient utilization rate and digestive enzyme activity, and the effects increased with the increasing dose of PYY antiserum. The possible mechanism of this effect is that PYY antiserum specifically neutralizes the endogenous PYY thus decreases its biological effects in rats.
This research was jointly supported by grants from the State Key Laboratory of Animal Nutrition (2004DA125184F1305), National Key Technology Research and Development Program of the Ministry of Science and Technology of China (2012BAD39B00).