Medicinal & Aromatic Plants
Open Access
ISSN: 2167-0412
ISSN: 2167-0412
Research Article - (2016) Volume 5, Issue 2
Essential oils are used for their reputed health benefits; reversal of inflammation, oxidative damage, and infection are some of the therapeutic effects that essential oils are believed to possess. In many cases, there is scientific evidence to back these claims. However, the purported anti-aging effects of essential oils are not yet supported by the literature. We set out to evaluate the effects of 31 oils on telomere length, believed to play a role in cellular senescence. Cells were subjected to oils in the presence or absence of hydrogen peroxide, and changes in telomere length were measured using a PCR-based assay. Two oils which showed telomere-protective effects were further characterized using a cytogenetic assay, staining for telomeres. Significant variability was seen among these oils with respect to their ability to induce an apparent increase or decrease in telomere length, and to protect against peroxideinduced telomere damage. Expression analysis showed that telomere-protective oils did not increase telomerase (hTERT) gene transcription, but reduced TERF-1, a suppressor of telomere length. Though this mechanism is not yet understood, these data provide a base upon which the community can build, and represent the first formal scientific foray into the study of essential oils on the telomere. Conclusion: Certain essential oils, including those from basil and rosemary, appear to have telomere-protective effects, mediated through a reduction of the TERF-1 telomere-suppressive protein.
Keywords: Telomerase; TERF-1; TRF1; TERT; Natural product; Basil oil; Rosemary oil
Considerable research and development work has been invested in the evaluation of plant products as potential remedies for common ailments. One subset of the natural product market, the essential oils (EOs), is currently experiencing accelerated growth and significant attention. Responsible for many of the benefits attributed to plant extracts, EOs are complex mixtures of volatile natural compounds isolated from various parts of the parent plant (seed, leaf, root, et cetera). Analysis of EOs by chromatography and spectrometry assays show that they can contain 20-60 constituents in vastly differing concentrations and compositions. Generally, these consist of two or three major components at high concentrations (in the range of 20- 70% of the whole), with multiple minor components, some in trace amounts [1]. Recognized as the primary ingredients of EO, the major components are generally believed to be responsible for much of the observed biological effects [1]. Acting alone, and/or working in concert with other constituents of the EO however, minor components have been shown to possess important biological activities. Now it is widely believed that the combined effects of the EO components, potentially acting in synergy, provide the rationale for the use of EOs and combinations thereof (rather than isolated components) for their health-promoting benefits [2].
Data suggest that the therapeutic effects of EOs arise largely from their reported antimicrobial, anticancer, antioxidant, anti-mutagenic, anti-inflammatory and anti-allergic properties, amongst others [1,3-6]; the research to provide actual mechanisms of action and scientific support for these products is severely lacking, although much anecdotal evidence exists for the usefulness of these products. One such claim, commonly touted by natural product proponents, is that EOs help to combat the effects of aging (wrinkles, organ failure, muscle weakness, et cetera).
The telomere theory of aging posits that, as somatic cells continue to divide over the life of an organism, their telomeres, the protective sequences at the end of each chromosome, gradually shorten [7,8]. Briefly, this is due to the inability of the DNA replication machinery to fully replicate the ends of linear chromosomal DNA, such that a small amount of sequence is lost with each round of replication (that is, each time a cell divides). After a specified number of cell divisions (known as the “Hayflick limit”) [9], sufficient chromosome material is lost that a cell’s replicative potential is exceeded and it senesces. This shortcoming can be overcome through the actions of the telomerase protein/RNA complex, which allows chromosome ends to be replicated without loss of genetic material. Stem cells, such as those responsible for replacing skin cells and blood cells, necessarily express telomerase and are not bound by this replicative limit. Telomerase activation is also a hallmark of cancer, as tumor cells exhibit limitless replicative potential [10]. This enzyme, however, does not act alone; several proteins either interact with telomerase, or regulate its action in one way or another. One such protein, TERF-1 (also reported as TRF-1), is a DNA-binding protein with specificity to telomeric DNA, which acts to inhibit telomere lengthening by acting in cis (blocking binding sites for telomerase) [11]. As telomeres lengthen due to the actions of telomerase, more binding sites for TERF-1 are generated, until a threshold is reached; at which point telomerase can no longer act. As telomeres eventually shorten, this threshold is no longer met, and telomerase is once more able to bind and extend chromosome ends, creating equilibrium. In this way, telomere length is maintained.
While telomere erosion is one means by which cells are believed to senesce, it is not thought to be the only pathway. Prolonged oxidative stress, whether through inflammation or exposure to reactive oxygen species (ROS)-generating chemicals, is also believed to play a role. While oxidative stress has many biological effects, it is known to cause direct damage to DNA, through the formation of several types of base lesions (modifications of the building blocks of DNA) [12]. If these damaged bases are not, or cannot be, repaired, this damage is made permanent and can lead to mutations, DNA breakage or cell death. If these lesions occur in the telomeric regions of DNA, telomere shortening can result, even in the presence of telomerase, as the telomerase enzyme cannot perform its function if damaged bases are present, and cannot counteract wholesale loss of DNA due to incomplete or improper DNA repair [8,13].
The present study was undertaken to determine the effect of a large subset of popular essential oils on telomere maintenance in the presence and absence of oxidative stress, in an effort to apply a measure of scientific rigor to the anti-aging claims made regarding these oils. The intention is to provide a base set of data, upon which additional studies can be built in the future by members of the nutraceutical and natural product community; to our knowledge no other study has evaluated such a large number of EOs for their effect on telomeres, and differences in cell origin, assay type, and assay procedure make it difficult to compare results from different studies with confidence.
Essential oils
Thirty one essential oils (peppermint [Mentha piperita], bergamot [Citrus Bergama], wintergreen [Gaultheria procumbens], grapefruit [Citrus x parasisi], basil [Ocimum basilicum], rosemary [Rosmarinus officinalis CT cineol], orange [Citrus sinensis], melaleuca [Melaleuca alternifolia], clove [Syzygium aromaticum], lemon [Citrus limon], balsam fir [Abies balsamea], birch [Betula alba], chamomile [Chamaemelum nobile], cinnamon [Cinnamomum verum], cypress [Cupressus sempervirens], eucalyptus globulus [Eucalyptus globulus], eucalyptus radiata [Eucalyptus radiata], frankincense [Boswellia carteri], ginger [Zingiber officinale], helichrysum [Helichrysum italicum], lavender [Lavandula angustifolia], lemongrass [Cymbopogon flexuosus], marjoram [Origanum majorana], myrtle [Myrtus communis], oregano [Origanum compactum], sandalwood [Santalum album], spruce [Picea mariani], tangerine [Citrus reticulate], thyme [Thymus vulgaris], vetiver [Vetiveria zizanoides], ylang ylang [Cananga odoratac]) were sourced through Améo, a subsidiary of Zija International (Lehi, UT, USA). Oils were diluted into cell culture medium (see below) before being applied to cells.
Cell culture
K562 cells (CCL-243, ATCC, and Manassas, VA, USA) were grown in RPMI media supplemented with 10% fetal bovine serum, and 1% Penicillin/Streptomycin solution in a humidified incubator with 5% CO2. This cell line was chosen based on a literature search as having telomeres of a sufficient length (long enough to detect, and well within the power of the assay to detect increases and decreases) to measure consistently in the PCR-based assay described below [14]. Cells were seeded at 6 × 105 cells per well (2 mL total, after addition of all test agents) in 6-well tissue culture plates for all studies, and allowed to equilibrate for at least one hour before exposure to treatment.
Treatment with test agents
Stock solutions (0.1%) of EOs were made immediately before treatment of cells, by performing a 1:1000 dilution of oil in cell culture medium. 100 μL of EO stock was added to treated wells, for a final concentration of 0.005%. A 50 mM stock solution of control antioxidant (phenyl-alpha-TERT-butyl nitrone, PBN) was made in water, and aliquots were kept at -20°C. 40 uL of PBN stock was added to appropriate test wells, for a final concentration of 1mM. A 10 mM hydrogen peroxide solution in cell culture medium was made immediately prior to treatment of cells from a 3% stock. 40 uL of this solution was added to appropriate treatment wells after all other additions, for a final concentration of 0.2 mM. Cells were exposed to test agents for 48 hours, at which point cells were centrifuged for 5 minutes at 500 × g to remove growth medium and washed once in PBS. The resultant cell pellet was used immediately for downstream applications, or stored at -80°C until needed (except in the case of PNA-FISH assays, for which the cell pellet was always used immediately after collection).
Nucleic acid extraction
To extract and purify genomic DNA for PCR-based telomere length studies, the Quick-gDNA Miniprep Kit (Zymo Research, Inc., Irvine, CA, USA) was used, according to the manufacturer’s instructions. RNA extraction for downstream qRT-PCR was carried out using the PureLink RNA Mini Kit (Thermo-Fisher Scientific, Grand Island, NY, USA).
PCR-based telomere length assay
The quantitative PCR (qPCR) procedure outlined by O’Callaghan and Fenech [15] is used as a determinant for telomere length, using 2 wells per DNA sample, 4 wells per treatment group. qPCR is carried out using the 7500 Fast Real-Time PCR System (Thermo-Fisher Scientific). Relative telomere content is then calculated as the average ratio of fluorescent signal in each treatment group to fluorescent signal in the untreated group.
Measurement of gene expression
Total RNA was converted to cDNA using the SuperScript VILO cDNA Synthesis Kit (Thermo-Fisher Scientific) and 1 μg input RNA. This first-strand cDNA was used directly in quantitative reverse transcription and PCR (qRT-PCR) for human TERT (probe/primer set Hs00972650_m1, Thermo-Fisher Scientific), or TERF-1 (probe/primer set Hs00819517_mH, Thermo-Fisher Scientific). GAPDH was used as a reference control (4326317E, Thermo-Fisher Scientific), as laid out in the TaqMan Fast Advanced Master Mix manual (Thermo-Fisher Scientific). PCR is carried out using the 7500 Fast Real-Time PCR System (Thermo-Fisher Scientific). Expression levels of TERT following treatment are normalized to control untreated cells, calculated using the ΔΔCT method.
Metaphase spread preparation and qualitative PNA-FISH telomere assay
For qualitative verification of the data collected from the PCRbased telomere assay, a telomere-specific peptide-nucleic acid probe was employed for fluorescent in-situ hybridization (PNA-FISH). The Telomere PNA FISH Kit/FITC (Dako Inc., Carpinteria, CA, USA) was used according to the manufacturer’s instructions, including the recommended protocol for metaphase spread preparation, with the following exceptions: Cells were synchronized with colcemid for 19 hours, to ensure a sufficient population of metaphase nuclei. Four fixation steps were performed to remove all traces of cytoplasm, rather than the three recommended. ProLong Diamond Antifade Mountant with DAPI (Thermo-Fisher Scientific) was used as the antifade reagent, and slides were allowed to cure for 24 hours following antifade application. Slides were visualized on a Nikon A1 fluorescent confocal microscope, set to detect FITC (peak excitation/emission 490/525 nm) and DAPI (peak excitation/emission 360/460 nm), at the University of Utah HSC Cell Imaging Research Core.
Effect of essential oils on telomere length
Thirty-one EOs were tested for their effect on telomere maintenance, in the presence or absence of H2O2, using a PCR-based assay described earlier. A concentration of 0.005% was used for each EO, which we have determined to be minimally cytotoxic over the time course studied. The results of this study are represented graphically in Figure 1. EOs are ordered according to the apparent telomere length following treatment with EO alone. Error bars represent standard error among replicate samples, performed over several days. It is evident that there is a deleterious effect of hydrogen peroxide on telomere maintenance in these cells, reducing apparent telomere length to approximately 40% of controls. This may be due to direct oxidative damage to the DNA, or to improper repair of peroxide-mediated lesions. It is of interest to note that some essential oils cause apparent increases in telomere length over those seen in untreated cells, when administered alone. In many cases, and especially following treatment with basil, clove or rosemary oils, the telomere-shortening effects of hydrogen peroxide are mitigated by EO treatment, leading to telomeres 60-80% of the size of those in untreated control cells. These data prompted us to verify the telomereprotecting effects of these oils in additional assays.
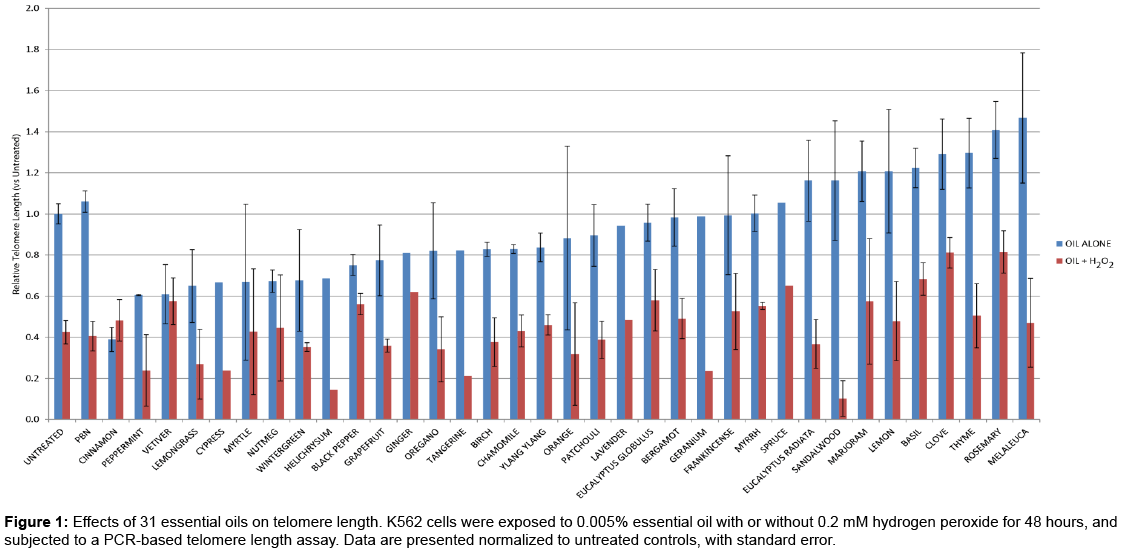
Figure 1: Effects of 31 essential oils on telomere length. K562 cells were exposed to 0.005% essential oil with or without 0.2 mM hydrogen peroxide for 48 hours, and subjected to a PCR-based telomere length assay. Data are presented normalized to untreated controls, with standard error.
To support the PCR-based telomere length study, a parallel study was performed, assaying two of the best H2O2-mitigating EOs (basil and rosemary) for their effect on telomeric DNA directly, using FISH. In this assay, a fluorescent telomere-specific peptide nucleic acid probe was employed, which binds to telomeric DNA sequences. Compacted metaphase chromosomes were extracted from cells treated with these EOs, and visualized using DAPI, a general DNA stain, as well as the fluorescent telomere probe, which is expected to bind at the ends of these chromosomes. The results of this assay are shown in Figure 2. While these data were not quantitative, it is apparent that, upon treatment with hydrogen peroxide, the telomere-specific signal is much reduced. In the presence of antioxidant or either of the two essential oils tested, this signal is restored. This corroborates the earlier quantitative findings and suggests that the effect of these oils on telomeres is indeed a repeatable phenomenon.
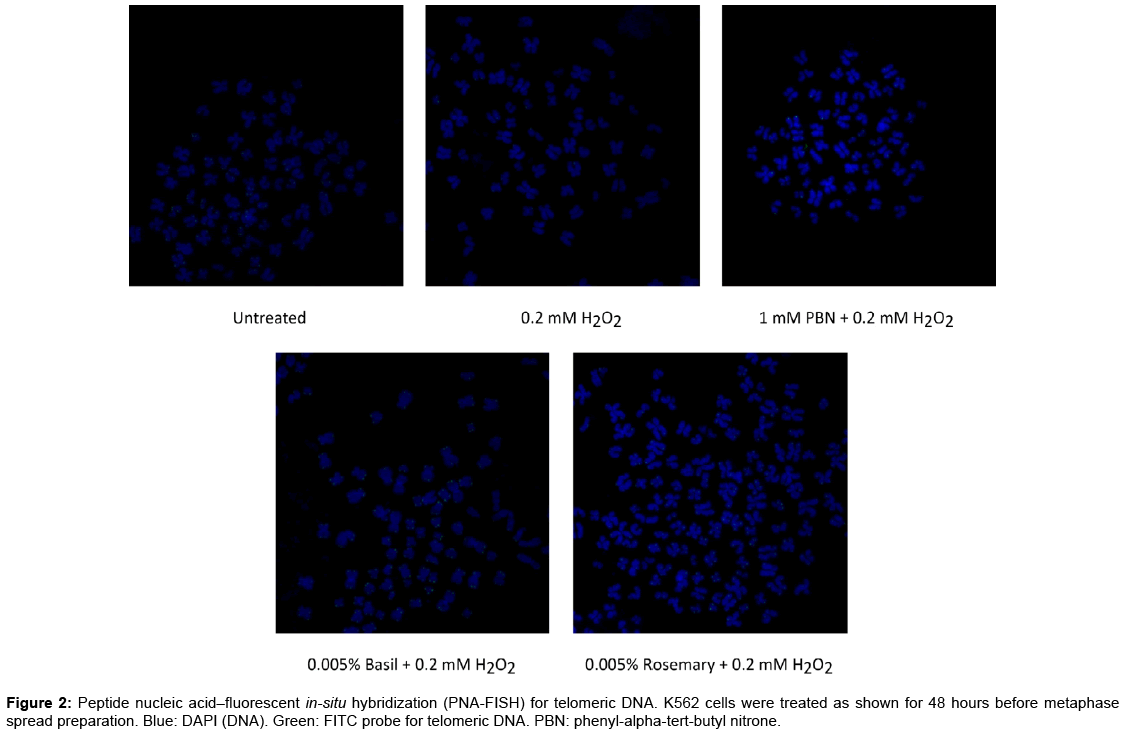
Figure 2: Peptide nucleic acid–fluorescent in-situ hybridization (PNA-FISH) for telomeric DNA. K562 cells were treated as shown for 48 hours before metaphase spread preparation. Blue: DAPI (DNA). Green: FITC probe for telomeric DNA. PBN: phenyl-alpha-tert-butyl nitrone.
Effect of EOs on telomerase and telomerase regulation
In order to determine if the effect of these telomere-preserving EOs on telomere length is due to changes in expression of the telomerase enzyme, qRT-PCR was performed, with the human TERT gene as a target. Cells were once again treated with either basil or rosemary oil, in the presence or absence of hydrogen peroxide, under the same conditions as the telomere length assays described above. Expression data are shown in Figure 3A, normalized to untreated controls. In apparent opposition to the telomere length data, TERT levels were decreased following treatment with these EOs, and increased following exposure to hydrogen peroxide. It appears that the EOs and PBN (a control antioxidant) mitigate the effect of peroxide on TERT to some extent, suggesting that this change in gene expression is related to oxidative stress. While this does not support the data seen earlier, this should perhaps not come as a surprise; K562 cells are immortalized, and known to express the telomerase enzyme. Increasing the expression of telomerase, then, may not result in an increase in telomere length, assuming that telomerase is not a limiting factor in these cells. These data can be rationalized in the context of oxidative stress, however. The TERT gene is controlled in part by oxidative stress, and is thought to play a protective role unrelated to telomere length maintenance in the face of ROS, which cause an increase in TERT gene expression [16,17]. When antioxidant-like molecules such as these EOs or PBN are present, this effect may be reduced or reversed, explaining the data we present here; TERT gene expression is increased by peroxide without a subsequent lengthening of telomeres, as telomerase is likely present in these cells in high enough amounts that additional enzyme would not be expected to impact telomere length. The shortening of telomeres seen may then be related to oxidative damage, or another unrelated mechanism. Of note is the fact that certain EOs have been shown in independent studies to upregulate members of the AP-1 transcription factor complex [18,19]. AP-1 overexpression is known to negatively regulate TERT [20] suggesting yet another mechanism by which these EOs may be downregulating telomerase expression.
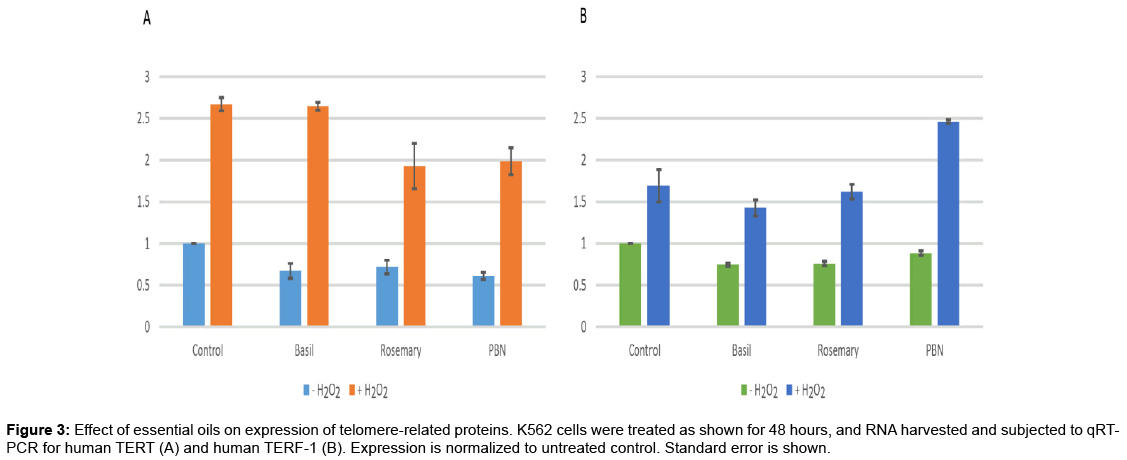
Figure 3: Effect of essential oils on expression of telomere-related proteins. K562 cells were treated as shown for 48 hours, and RNA harvested and subjected to qRTPCR for human TERT (A) and human TERF-1 (B). Expression is normalized to untreated control. Standard error is shown.
The TERF-1 gene, a negative regulator of telomere length [11], was also studied for its response to EO and oxidative stress. Using the same cDNA synthesized in the TERT assay above, quantitative PCR was performed for TERF-1, and the expression data are represented in Figure 3B. In this case, a similar effect was seen as that on TERT: reduction of TERF-1 expression as a consequence of EO treatment, and an increase in expression following oxidative stress. The mechanism which mediates this effect is not clear at this time; it is known that TERF-1 interacts with KEAP-1 [21], a protein involved in the oxidative stress response (which also binds to Nrf-2, an activator of genes bearing the antioxidant response element, ARE, in their promoters). How this TERF-1 / KEAP-1 interaction is related to oxidative stress, however, is as yet unknown. Regardless of the mechanism of action, the effect of these two essential oils on TERF-1 expression may partially explain the results seen in our earlier telomere length assays; EOs reduce the amount of TERF-1 available to bind to telomeres, which tips the equilibrium in favor of telomerase and telomere lengthening. Similarly, the upregulation of TERF-1 by peroxide treatment could move the equilibrium in the other direction and lead to shortened telomeres. While these outcomes are potentially related to oxidative damage (and protection from oxidative damage by EOs), it is interesting to note that PBN, a known antioxidant, does not mitigate the TERF-1 upregulation by hydrogen peroxide; when both PBN and peroxide are present, TERF-1 expression is higher than in cells treated with peroxide alone. This suggests that the impact of EOs on telomere length may not be due solely to their antioxidant properties.
To our knowledge, very few controlled scientific studies have been performed linking essential oils to effects on the telomere, based on a search of PubMed entries; anecdotal and unsupported claims that EOs affect telomerase or telomere length abound, however. One rare scientific study from a group in China showed that pine needle EO was able to downregulate telomerase expression in a hepatocellular (liver) tumor cell line [22], which supports the data we have presented here, but evidence otherwise is sparse, and often presented outside of the confines of peer review. Here, for the first time, we have demonstrated the effect of 31 EOs on telomere length, and have found that there are significant differences among them; some, such as cinnamon and peppermint, appear to have a negative effect on telomere length, although they bear other health benefits. Others, including basil and rosemary EOs, lead to telomere elongation, and seem to protect cells against the loss of telomeric DNA induced by oxidative stress. While this may be due in part to the antioxidant properties of these oils, absorbing ROS directly, the data we present indicate that there are other factors at play as well. While these telomere-protecting oils do not induce telomerase production (expression of the TERT gene is inhibited following treatment with these EOs), a telomere-regulating protein, TERF-1, is downregulated, which may be partially responsible for the increased telomere length seen. While unexpected, this presents a novel mechanism of action for certain essential oils that has not been reported or even proposed to this point. In addition, this suggests that these oils can maintain telomere length without increasing telomerase expression. Since activation of telomerase is one of the “hallmarks of cancer”, a series of changes that must occur in order for a healthy cell to transform and become tumorigenic, this finding suggests that cellular senescence can be combatted by systemic, long-term supplementation with EOs, without increasing cancer risk.
The work described here is by no means exhaustive; the mechanism through which TERT and TERF-1 are abrogated by these telomerelengthening EOs is not yet understood, and there may be other factors contributing to this effect that we have not elucidated in this study. This work represents a foundation in understanding the role that EOs may play in telomere modulation and reversal of senescence, which we hope will be built upon by the natural products community. It is clear that these effects are not common to all essential oils, indicating that attention must be paid to the desired outcome when selecting an oil (or combination of oils) for supplementation. As natural products research expands, and the public learns more about the health benefits of essential oils, we believe that these data will be an invaluable addition to the nutraceutical community knowledge base.
After evaluation of 31 commonly-used therapeutic essential oils, we have shown here that certain of these, including oils from basil (Ocimum basilicum) and rosemary Rosmarinus officinalis CT cineol are capable of increasing the apparent length of telomeres in cell culture, when administered at low (subtoxic) doses. This effect was demonstrated using a previously-described PCR-based telomere length assay, and verified by qualitative cytogenetic analysis. Gene expression analysis revealed that these oils did not cause a concomitant increase in the expression of hTERT, the gene coding for the catalytic subunit of the telomerase protein complex, but instead downregulated the TERF-1 telomere length suppressor. We conclude that essential oils may have potential use as a supplement to impact cellular senescence, once this mechanism has been fully elucidated.
Wasatch Scientific Services (Murray, UT, USA), an independent contract research organization, carried out the work described in this manuscript, under guidance of the author. Wasatch Scientific Services was also employed in the preparation of this manuscript.
The specific essential oils utilized in this study were sourced through Améo, a subsidiary of Zija International, of which the author is an employee. The data presented here were collected as part of this company’s characterization of its product portfolio, and are made public here in the hope that the broader community will benefit from their dissemination. Wasatch Scientific Services was paid for their laboratory and manuscript preparation services, but is not otherwise affiliated with Zija International or its subsidiaries.