Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Research Article - (2012) Volume 1, Issue 7
To evaluate the protective effects of Vitamin C and Vitamin E against Mercuric chloride -induced reproductive toxicity and to study the mechanisms underlying these effects. Male Wistar rats were orally administered mercuric chloride (10 mg/100 gm b.wt), (10 mg/100 gm b.wt) of mercuric chloride + (40 mg/100 gm b.wt) of Vitamin C and (10 mg/100 gm b.wt) of mercuric chloride + (20 mg/100 gm b.wt) of Vitamin E were administered orally to male albino rats for 30 days. Treatments with either Vitamin C or Vitamin E resulted in a significant protection of reproductive function. Both vitamins reduced the extent of mercuric chloride-induced reproductive toxicity, as evidenced by decrease in sperm abnormality and increase in sperm motility. Mercuric chloride-induced alterations in testis lipid peroxidation (MDA) were also markedly improved by test vitamins, and mercuric chloride -induced alterations in the testis antioxidation defense system were profoundly prevented by vitamins. In groups, where mercuric chloride was combined with either Vitamin C or Vitamin E, antioxidation enzymes like superoxide dismutase (SOD), reduced glutathione (GSH), and catalase (CAT) were significantly elevated compared to the mercuric chloride-treated group. The results provide further insight into the mechanisms of mercuric chloride-induced reproductive toxicity and confirm the antioxidant potential of both Vitamin C and Vitamin E.
<Keywords: Mercuric chloride; Testicular toxicity; Protection; Male rats; Testis; Oxidative stress; Antioxidants; Vitamin C; Vitamin E
Mercury is a naturally occurring metal, which can exist in several forms. Metallic mercury is a shiny, silver-white, odorless liquid, which forms a colorless, odorless gas if heated. Mercury combines with other elements such as chlorine, sulfur, or oxygen to form inorganic mercury compounds or salts, which are usually white solids. Mercury also combines with carbon to make organic mercury compounds, the most common of which is methyl mercury [1].
Historically, mercuric chloride (HgCl2) pollution has posed a serious health hazard to humans. Mercuric chloride (HgCl2) from industries surrounding the area was discharged directly into the bay, producing elevated levels in fish [1]. The extreme morbidity and mortality in humans who consumed these fish clearly demonstrated the relationship between HgCl2 ingestion and human health. Adverse effects of HgCl2 on fish-eating mammals have also been reported [2], suggesting a link between environmental HgCl2 and wildlife health. Although strict regulations now limit the discharge of HgCl2 directly into bodies of water, indirect contamination continues. Augmented output of airborne HgCl2 from anthropogenic sources, such as coalfired utilities and the burning of municipal wastes are likely to be responsible for observed increases in atmospheric levels [3]. Although controversial, reports have indicated that exposure to HgCl2 might impair male reproductive health. For example, several reports have indicated that HgCl2 ingestion can cause reduced spermatogenesis and steroidogenesis in rodents [4].
Inorganic mercury-induced testicular damage is a known fact in experimental animals [5]. In humans, mercury induces loss of libido [6], hypospermia, astheno-spermia and teratospermia [7]. Owing to the relative spermiotoxicity of HgCl2, almost all patients under chemotherapy will show temporary or permanent azoospermia. The damage to both spermatogenesis and testicular endocrine function can be temporary or permanent base on the applied dose of HgCl2 [8]. Testicular damage is also developed in various animal species [9,10]. Within days of HgCl2 injection, animals develop severe testicular damage, which is characterized by spermatogenic damages, germ cell apoptosis, Leyding cell dysfunction and testicular steroidogenic disorder [10].
Many environmental contaminants have been reported to disturb the pro-oxidant or antioxidant balance of the cells thereby inducing oxidative stress [11]. Oxidative stress in biological systems originates as the result of an imbalance between the generation of oxidizing species and cellular antioxidant defences [11,12]. The radical chain reaction of lipid peroxidation appears to be a continuous physiological process. The process, if out of control, can alter essential cell functions to cell death [13]. Oxidative stress has been shown to mediate the HgCl2 -induced reproductive toxicity [14].
A major contributor to non-enzymatic protection against lipid peroxidation (LP) is vitamin E and vitamin C, a known free radical scavenger [12]. Vitamin E as a lipid soluble, chain breaking antioxidant [13] plays a major role against oxidative stress, and prevents the production of lipid peroxides by scavenging free radicals in biological membranes [15]. Vitamin C, an endogenous water-soluble natural antioxidant, chain breaking antioxidant and is found in both intra cellulary and extra cellularly [16]. It prevent lipid peroxidation due to peroxy radicals, protects against DNA damage by hydrogen peroxide radical. Vitamin E and Vitamin C have been widely investigated due to its action against oxidative stress [17], its protective role on biological membranes [18].
The administration of antioxidants such as vitamin E [19], or vitamin C [14] has been reported to prevent different HgCl2-associated side effects. On the other hand, the mechanism of HgCl2-induced testicular toxicity is not quite understood yet and protective remedies against HgCl2-induced testicular damage are scarce. This investigation was set to evaluate the protective effects of Vitamin C and Vitamin E against HgCl2-induced testicular toxicity in male rats and to study the mechanisms underlying these effects.
Chemicals
Mercuric chloride, Vitamin C, Vitamin E, Thiobarbituric acid, reduced glutathione, 5,5-dithiobis (2- nitrobenzoic acid), Folin’s reagent, epinephrine, SOD enzyme, H2O2 and bovine serum albumin were obtained from Sigma Chemical Co., Mumbai, India. All other chemicals were obtained from local commercial suppliers.
Experimental animals
Twelve - week-old adult Wister albino male rats weighing between 175 - 225g, along with supplies of their standard diet, were obtained from animal house centre of “Jawaharlal Nehru Institute of Post Graduate Medical Educations and Research” (JIPMER), Pondicherry. The study design was approved “Animal ethic committee” J. J. College of Arts and Science, Pudukottai, Tamil nadu, India.
Treatment regime
Animals were divided into six groups of six rats each for 30 days. Group I was received distilled water and served as the control. Group II was administered mercuric chloride (10 mg/100 gm. b.wt). Group III received mercuric chloride (10 mg/100 gm b.wt) and Vitamin C (40 mg/100 gm b.wt), Group IV was administered mercuric chloride (10 mg/100 gm. b.wt) and Vitamin E (30 mg/100 gm. b.wt), Group V was received (40 mg/100 gm b.wt) of Vitamin C and Group VI was administered (30 mg/100 gm. b.wt) of Vitamin E.
Sample preparation
Following diethyl ether anesthesia, blood was collected from the retro-orbital plexus [20]. Following sacrifice, testes and epididymis were removed and weighed. The organ weight/body weight ratio x 100 was calculated and expressed as a relative organ weight. For histopathological examination, one testis was immediately fixed in 10% buffered formalin. For biochemical determination, another testis was homogenized in ice-cold KCl4 (150 mM). The ratio of tissue weight to homogenization buffer was 1:10. From the latter, suitable dilutions for determination of the levels of GSH, LP product MDA, total proteins, and activities of SOD and CAT were prepared in suitable different buffers. To obtain serum, blood was collected in centrifuge tubes and centrifuged in a refrigerated centrifuge (4°C) at 3000 r.p.m. for 20 minutes.
Sperm motility, sperm count and abnormality test
After removing each epididymis, it was immediately weighed, minced in 5ml of physiological saline and then incubated at 37°C for 30 minutes to allow sperms to swim from the epididymal tubules. After mixing, one drop was placed onto a warm microscope slide and cover slip was placed over the droplet. The percentage of motile sperm was recorded with a phase contrast microscope at 400X magnification. The cover slip was removed and sperms were allowed to air dry and then stained with 1% eosin and examined for morphology abnormalities. Three hundred sperm from different fields were examined with a phase contrast microscope. Total sperm number was determined by using a Neubauer hemocytometer by a method of Yokoi et al. [21]. The total number of sperm per gram of epididymis was then calculated.
Biochemical assays and histopathology
The GSH content of testis homogenate was determined using the method described by Van Dooran et al. [22]. The GSH determination method is based on the reaction of Ellman’s reagent 5, 5-dithiobis (2- nitrobenzoic acid) (DTNB) with the thiol group of GSH at pH 8.0 to produce 5-thiol-2- nitrobenzoate, which is yellow. Malondialdehyde (MDA) is the most abundant individual aldehyde resulting from lipid peroxidation (LP) breakdown in biological systems and is used as an indirect index of LP [23]. Determination of MDA in biological materials, as described in Uchiyama and Mihara [24], is based on its reaction with thiobarbituric acid (TBA) to form a pink complex with absorption maximum at 535 nm. The activity of SOD enzyme in testis homogenate was determined according to the method described by Sun and Zigman [25]. This method is based on the ability of SOD to inhibit the auto-oxidation of epinephrine to adrenochrome and other derivatives at alkaline pH. These derivatives can easily be monitored in the near-UV region of the absorption spectrum. CAT activity was determined by measuring the exponential disappearance of H2O2 at 240 nm and expressed in units/mg of protein as described by Aebi [26]. The total protein content of testis was determined according to the Lowry method as modified by Peterson [27]. Absorbance was recorded using Shimadzu recording spectrophotometer (UV-160) in all measurements. For the histological examinations, pieces of testis were fixed in 10% neutral phosphate-buffered formalin and hydrated tissue sections, 5μm in thickness, were stained with Hematoxylin and Eosin. The sections were examined under a Nikon light microscope.
Testosterone levels
Serum was collected by allowing trunk blood to clot overnight at 4°C. Interstitial fluid was collected into a centrifuge tube by perforating the tunica albuginea at the distal pole of the testis several times with a needle and centrifuging at 54 × g at 0°C for 15 min [28]. Samples were stored at −20°C. For each rat, testosterone levels in trunk blood and in the interstitial fluid of the testes were measured using fully automated chemiluminescence analyser at Vivek laboratories, Nagarkovil, Tamil nadu, India.
Statistical analysis
The results were expressed as Mean ± SD, and statistical analysis was performed using Student “t” test.
Effects of HgCl2 on the weights of reproductive organs
Administration of HgCl2 individually treated rats showed significantly (p<0.05) decreased relative weights of the testes (Figure 1a) and epididymis (Figure 1b). However, No significant changes were observed in the relative weights of testes and epididymis in animal groups treated with a combination of Vitamin C or Vitamin E and HgCl2 treatment (Figure 1a and Figure 1b).
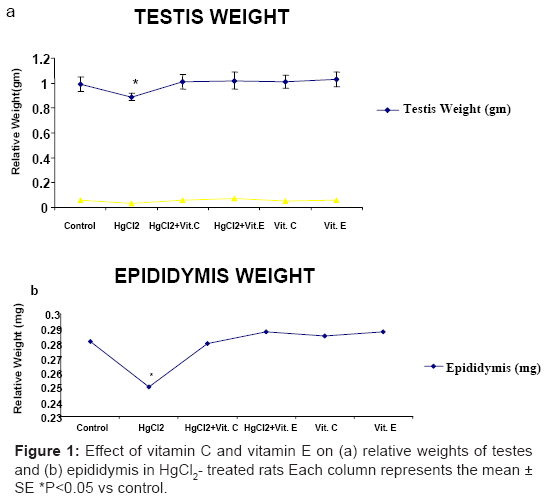
Figure 1: Effect of vitamin C and vitamin E on (a) relative weights of testes and (b) epididymis in HgCl2- treated rats Each column represents the mean ± SE *P<0.05 vs control.
Protective effect of Vitamin C and Vitamin E on sperm parameters
The number of sperm per gm of epididymis was significantly reduced (P<0.01) in the HgCl2 a lonely treated rats (Figure 2a). A combination of HgCl2 with Vitamin C had no effect on this parameter; similar results were obtained in HgCl2 treatment with vitamin E. The effect of HgCl2 on sperm motility and abnormality is shown in (Figure 2b). The results indicate that sperm motility was reduced (P<0.001) and sperm abnormality was increased (P<0.05) in HgCl2 treated rats (Figure 2c). While the treatment with Vitamin C or Vitamin E was found to protect against the changes of HgCl2 induced parameters.
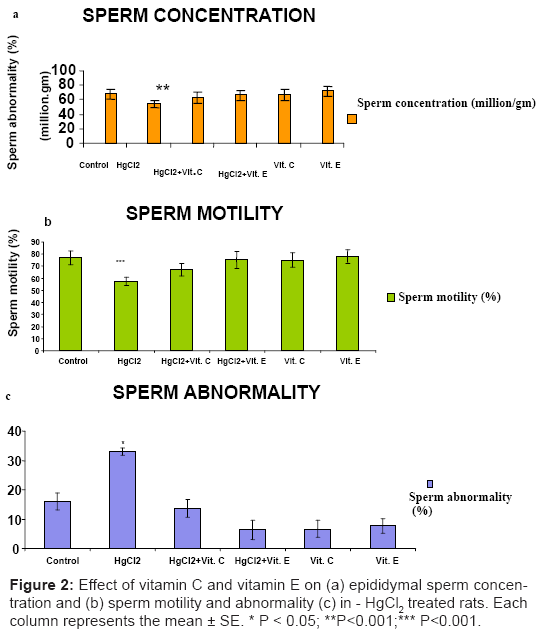
Figure 2: Effect of vitamin C and vitamin E on (a) epididymal sperm concentration and (b) sperm motility and abnormality (c) in - HgCl2 treated rats. Each column represents the mean ± SE. * P < 0.05; **P<0.001;*** P<0.001.
Significantly declined (P<0.05) in testicular content of reduced GSH (Figure 3a), activities of CAT and SOD (Figure 4a and Figure 4b) in HgCl2 – treated rat, while significantly increased (P<0.05) in the level of MDA was recorded after HgCl2 treatment (Figure 3b). These markers of oxidative stress did not differ significantly from control level, when Vitamin C or Vitamin E was administered with HgCl2.
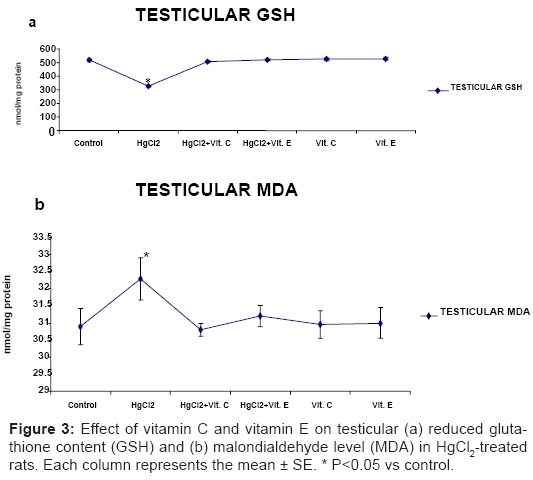
Figure 3: Effect of vitamin C and vitamin E on testicular (a) reduced glutathione content (GSH) and (b) malondialdehyde level (MDA) in HgCl2-treated rats. Each column represents the mean ± SE. * P<0.05 vs control.
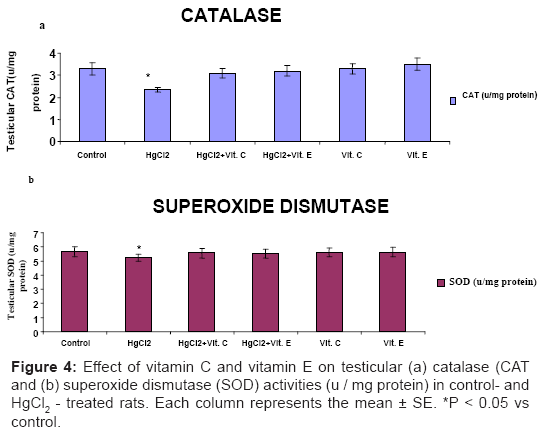
Figure 4: Effect of vitamin C and vitamin E on testicular (a) catalase (CAT and (b) superoxide dismutase (SOD) activities (u / mg protein) in control- and HgCl2 - treated rats. Each column represents the mean ± SE. *P < 0.05 vs control.
Testosterone level
Serum concentration of testosterone significantly (P < 0.01) decreased in the HgCl2 treated rats than control groups (Figure 5). Similarly, testicular testosterone also significantly decreased (P < 0.01) in HgCl2 treated animals as compared to control rats (Figure 5). However, did not evoke any significant changes in the concentration of serum testosterone and testicular testosterone in the combination of HgCl2 vitamin C or vitamin E treated animals and same results were measured in vitamin C or vitamin E individually treated rats for serum testosterone and testicular testosterone (Figure 5).
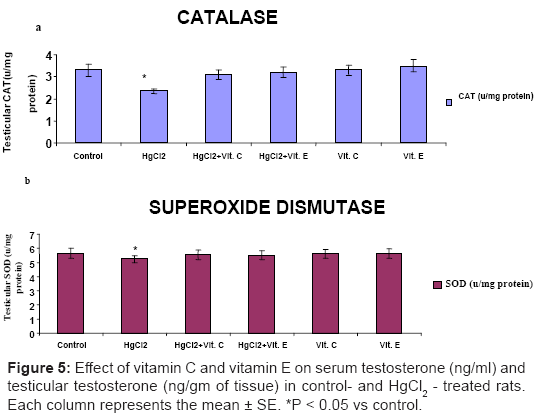
Figure 5: Effect of vitamin C and vitamin E on serum testosterone (ng/ml) and testicular testosterone (ng/gm of tissue) in control- and HgCl2 - treated rats. Each column represents the mean ± SE. *P < 0.05 vs control.
Histopathological effect of Vitamin C and Vitamin E
Severe degeneration of seminiferous tubules, depletions in germ cells and reduction in Leydig cells were clearly seen in the HgCl2-treated group. Debris from the degeneration of cellular components was seen in the lumen. Congestion of blood vessels was also observed between tubules. Animals treated with Vitamin C or Vitamin E showed normal testicular morphology and spermatogenesis with slight degeneration of spermatids and spermatozoa (Figure 6).
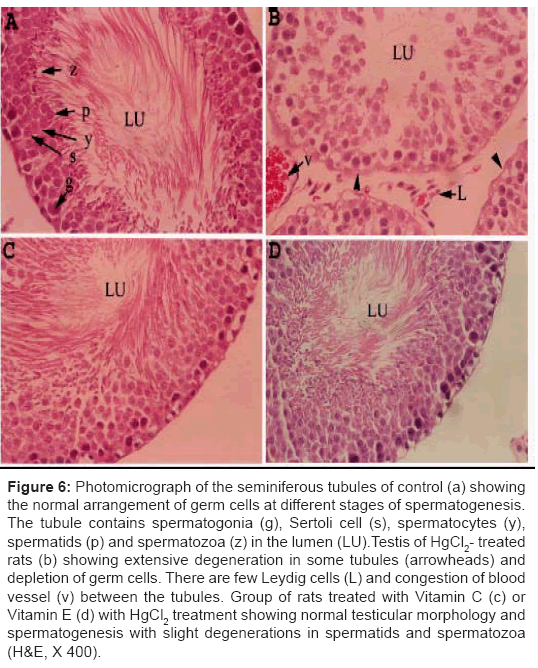
Figure 6: Photomicrograph of the seminiferous tubules of control (a) showing the normal arrangement of germ cells at different stages of spermatogenesis. The tubule contains spermatogonia (g), Sertoli cell (s), spermatocytes (y), spermatids (p) and spermatozoa (z) in the lumen (LU).Testis of HgCl2- treated rats (b) showing extensive degeneration in some tubules (arrowheads) and depletion of germ cells. There are few Leydig cells (L) and congestion of blood vessel (v) between the tubules. Group of rats treated with Vitamin C (c) or Vitamin E (d) with HgCl2 treatment showing normal testicular morphology and spermatogenesis with slight degenerations in spermatids and spermatozoa (H&E, X 400).
The protective effect of vitamin C and vitamin E on testicular toxicity of HgCl2, were investigated in male albino rats. To our knowledge, the present study planned to evaluates the protective effect of vitamin C and vitamin E against testicular damage induced by HgCl2 toxicity in experimental animals. In this report, a single dose of HgCl2 (10 mg/100 gm b wt) induced severe reproductive toxicity in adult male rats. Treatment with HgCl2 has been shown to cause testicular damage in various animal species and in humans [9,10]. The HgCl2-induced testicular damage in animals is commonly associated with spermatogenic damage, germ cell apoptosis, Leydig cell dysfunction and testicular steroidogenic disorder [29].
B. The present result showed a significant reduction in the relative weight of testes and epididymis as well as a decreased in the quality of epididymal sperm (sperm count, motility and morphology) after treatment with HgCl2 [30]. The results of this study suggest that a diet containing low levels of HgCl2 can lead to elevated concentrations of HgCl2 in the testes, decreased levels of testosterone in serum and testicular interstitial fluid, a decreased in cauda epididymal sperm storage [1]. This HgCl2-induced testicular damage was also confirmed by histopathological lesions [31]. Many investigators have reported a correlation between MDA and HgCl2-induced complications [14,32]. In this study, HgCl2-treated animals showed an elevation in testicular MDA level versus the control group. A possible explanation for the increased MDA concentration might be the decreased formation of antioxidants in HgCl2-induced tissues, which in view of the augmented activity of reactive oxygen species (ROS) in HgCl2 treated tissues, potentially explains the resulting increase in MDA production. GSH concentration and activities of CAT and SOD in the testes were also lower in the HgCl2-treated animals relative to control animals. It has been suggested that HgCl2 generates free radicals by interacting with DNA [33], which interferes with antioxidant defense system and results in the tissue injury. Therefore, over production of free radicals and hence oxidative stress may account, at least in part, for testicular injury associated with HgCl2 treatment. Recently, much attention has been focused on the protective effects of antioxidants and naturally-occurring substances against HgCl2-induced nephrotoxicity [14,32].
However, little is known about vitamin C and vitamin E act as protective agents against HgCl2-induced testicular toxicity. Administration of vitamin C or vitamin E with HgCl2 treatment clearly restored the testicular damage and quality of sperm, in addition to retaining the control values of oxidative stress markers. Accumulating evidence suggests that the protective effect of vitamins against oxidative damage could be attributed to its anti-oxidative properties [34]. This prevention of the HgCl2-induced oxidative stress damage in rat with vitamin C and vitamin E supports the hypothesis that part of the mechanism of testis damage is attributed to the overproduction of free radicals. (Figure 6): Photomicrograph of the seminiferous tubules of control (a) showing the normal arrangement of germ cells at different stages of spermatogenesis. The tubule contains spermatogonia (g), Sertoli cell (s), spermatocytes (y), spermatids (p) and spermatozoa (z) in the lumen (LU). Testis of HgCl2- treated rats (b) showing extensive degeneration in some tubules (arrowheads) and depletion of germ cells. There are few Leydig cells (L) and congestion of blood vessel (v) between the tubules. Group of rat treated with vitamin C (c) or vitamin E (d) and HgCl2 showing normal testicular morphology and spermatogenesis with slight degenerations in spermatids and spermatozoa (H&E, X 400).
Administration of vitamin C or vitamin E with HgCl2 treatment also attenuated testicular damage induced by HgCl2 treatment as shown by the normal sperm count, normal sperm morphology and low histopathological changes in comparison to the HgCl2 -treated animals. The protective effect of vitamin C or vitamin E is accompanied by normalization of antioxidant activity in testis and associated decrease of MDA. Vitamin C, which would neutralize H2O2 and would protect the plasma membrane from lipid peroxidation [14,35], Vitamin E has been shown to lower lipid peroxidation and increased glutathione content in blood of rats [36]. Indeed, the protective effect of Vitamin E against HgCl2 toxicity might be mediated not only by its potent antioxidant properties but through its androgenic activities as well. This activity was reflected by the increase of testis weight and serum testosterone levels [37].
In conclusion, we showed that oxidative stress contributes to the testicular toxicity induced by HgCl2 in male rats. Either vitamin C or vitamins E were shown to have a potent protective effect on HgCl2 - induced testicular damage and oxidative stress in rats. The protective effect of vitamin C or vitamin E may be due to their antioxidant properties.
Authors are wishing to thank the Management of J. J. College of Arts and Science, Pudukottai, Tamil nadu, India for providing the facilities.