Medicinal & Aromatic Plants
Open Access
ISSN: 2167-0412
ISSN: 2167-0412
Research Article - (2015) Volume 4, Issue 1
Insulin resistance is a serious challenge in diabetes treatment. Around the world, several plants are used in the treatment of diabetes and experimental studies clearly demonstrated that insulin resistance can be reduced by some antidiabetic plants. More than 100 plants have been reported to improve insulin sensitivity. The aim of this study was to review the effect of three popular medicinal plants that have generated broad interest during the past decade and used traditionally in the management of diabetes on insulin resistance. These plants are: Momordica charantia, Camellia sinensis and Cinnamomum species. We demonstrated that these plants effectively reduced insulin resistance in animal models involving different molecular mechanisms. However, clinical investigation remains needed to clearly affirm the effect of these plants on insulin resistance.
Keywords: Medicinal plant; Ethnopharmacology; Mechanism of action; Diabetes
Insulin resistance, a reduced biological effect of endogenous or exogenous insulin, is a common biochemical entity which is associated, either directly or indirectly, with a range of non-communicable human diseases. Insulin resistance may be entirely genetically determined (as in rare syndromes of severe insulin resistance) or acquired, either during intrauterine development or during adolescence and adult life [1-4]. Relative insulin resistance is also a transient feature of a number of physiological states in human [5].
The clinical impact of insulin resistance ranges from subclinical hyperinsulinaemia to major life-limiting disturbances of carbohydrate and lipid metabolism [6-9]. The main clinical concern derives from the association between impaired insulin action and the development of vascular disease [10-13]. Microvascular disease is a complication of type 2 diabetes mellitus, in which insulin resistance is a prominent feature. Atherosclerotic macrovascular disease, on the other hand, has a more complex association with insulin resistance that extends beyond hyperglycaemia [14].
The Ebers Papyrus written in approximately 1550 B.C. provides the earliest documentation about the use of plants in the treatment of conditions associated with diabetes. In India, the early Ayurvedic texts such as the Sushruta Samhita and the Charaka Samhita written in the 4th to 5th century B.C [15-18] describe the use of about 760 and 500 species of medicinal plants, respectively, including those prescribed for conditions such as glycosuria, polyphagia, and polyuria associated with diabetes [19-25]. In China, Ben Jing, written in about 104 B.C., contains detailed descriptions of 252 species with reference to those used to treat diabetes. It is easier to track the traditional and modern uses of species in the treatment of diabetes in cultures with a strong written culture like those in India, China, and the Middle East as well as in South America and Africa, where much less documentation is available [26,27].
Plants thought to play a role in the treatment of diabetes are taken as food or as medication. Reviews remain a classic reference work for those involved in using ethnobotanical data to study the potential use of plants in the treatment of diabetes. It has been based on a search of information about antidiabetic plants up to the year 1994 available in the computer database NAPRALERT at the College of Pharmacy, University of Illinois, Chicago [28-33]. Furthermore, studies identified about 1200 species of plants from 725 genera representing 183 families [34-36].
The insulin resistance syndrome, also known as the metabolic syndrome, is a common pathophysiological condition which is implicated in the development of type 2 diabetes, atherosclerosis, dyslipidemia, and hypertension [37-40]. The leading cause of mortality in people with the metabolic syndrome is cardiovascular disease (CVD). The close association between the metabolic syndrome and CVD may be due to the dyslipidemia [41-43]. The dyslipidemia observed in the metabolic syndrome is characterized in part by high plasma triglycerides and low high-density lipoprotein (HDL)- cholesterol concentrations [44-46]. Although we have made major strides in treating the dyslipidemia associated with type 2 diabetes with both lipid-lowering and anti-diabetic drugs, there is continued controversy on what constitute the optimal diet in treating this disorder [47-49]. Other than unsaturated fats and fiber, little is known about what nutrients may be beneficial in treating metabolic dyslipidemia [50,51]. There is growing evidence that medicinal plants may be used to treat metabolic dyslipidemia [52-55]. Animal studies have shown that green tea intake can modulate lipid metabolism in a number of diabetic animal models [56-58]. Furthermore, human intervention studies have shown that fiber plants consumption reduces lipid levels; however, the effects were found to be mild [59,60]. One possible way to obtain a more noticeable benefit would be to use an extract containing a higher amount of the active ingredient, namely green tea catechins [61-64]. Today, herbal green tea leaf extracts enriched with catechins are commercially available and are widely used in the treatment of many ailments, including CVD and cancer [65-69].
Insulin is the leading hormone that regulates blood glucose and fat metabolism. Insulin resistance is a condition in which circulating insulin decreases in responsiveness to target tissues, namely skeletal muscle, adipose tissue, and the liver. In Type 2 diabetes, the sensitivity of insulin decreases by β-cell dysfunction, insulin receptor mutation, and obesity factors [70-72]. Insulin sensitivity may be affected by many circulating lipids, which included hypertriglyceridemia and increased FFAs. Raised plasma FFA level is an important inductor of both peripheral and hepatic insulin resistance because it inhibits insulin signaling [73-77]. In addition, hypertriglyceridemia is also an important maker of insulin resistance. In the literature, more than 300 publications are related to the effect of plants on insulin resistance [78-80]. We chose three examples to illustrate the potentialities of plants in the management of insulin resistance [81-83].
Momordica charantia (Bitter melon)
The fructose fed rats model is a model that mimics the human metabolic syndrome in many aspects, including hypertriglyceridemia, insulin resistance, and compensatory hyperinsulinemia [84-86]. Insulin resistance was developed by the administration of a high-fructose diet. The treatment with bitter melon significantly ameliorated insulin resistance [84,85]. Moreover, the result of suppressing the elevated plasma TG and TC by bitter melon was consistent with previous studies in different animal models. Bitter melon has no dose-dependent effect in FFA, TG, and TC [87]. One possible explanation is due to the complex ingredients. Nevertheless, bitter melon and ROS suppress the fructose diet-induced increases in the concentrations of FFA, TG, and TC [85,87]. Bitter melon, traditionally used as an anti-diabetic herb, is effective to improve insulin resistance in a rat model of type 2 diabetes possibly by decreasing blood glucose through the regulation of PPARsmediated pathway. PPARs are ligand-activated transcription factors belonging to the nuclear receptor super family, and PPAR ligands include fatty acids and eicosanoids. PPARγ plays a key role in glucose homeostasis and differentiation of fat cells. PPARγ ligand activates PPARγ [88]. TZD, a synthetic PPARγ ligand, significantly increased insulin sensitivity. It has been suggested that PPARγ ligand up-regulated the expression of genes involved in glucose uptake and lipid storage in adipocytes. In addition, bitter melon increased the mRNA expression of PPARγ in adipose tissue and the observed down-regulation of leptin expression [89]. Therefore, it is possible that bitter melon behaved similar to several PPARs ligands. Furthermore, a significant decrease in the concentrations of TG and FFA by administration of bitter melon is possibly mediated by the increased expression of PPARs. It is apparent that insulin sensitivity is regulated by adipocytokine, a wide group of bioactive proteins produced by adipose tissues. PPARγ regulated genes including leptin and adiponectin involved in glucose and lipid metabolism. Exogenous leptin administration may improve insulin resistance [90]. Therefore, bitter melon at low dose significantly inhibited the abnormal increase of leptin expression in white adipose tissue. Bitter melon also prevented fructose-induced abnormalities in leptin levels and improved insulin resistance. Adiponectin plays a protective role against insulin resistance. Administration of adiponectin to diabetic mice enhances insulin sensitivity and attenuates plasma NEFA levels and glucose excursions. Reports have demonstrated an increase in plasma adiponectin levels in response to treatment with PPARγ agonists. Thiozodindiones (TZDs) affect adiponectin levels through the activation of its promoter activity according. Insulin resistance is thought to be the core of the metabolic syndrome (MS) [91]. The fructose fed rats model is a model that mimics the human MS in many aspects, including hypertriglyceridemia, insulin resistance, and compensatory hyperinsulinemia. PPARγ is mainly present in adipose tissue. Bitter melon induced a significant increase in levels of adiponectin and the expression of PPARγ, perhaps mediated by this increase, the reversal of the entire deleterious metabolic indices [92].
Recent studies in humans have shown that plasma adiponectin regulates insulin receptor tyrosine phosphorylation. Knock-out mice under a fat/carbohydrate-rich diet develop insulin resistance and display the impaired insulin receptor substrate (IRS) associated PI3K activity in their muscles. The molecular mechanism by which adiponectin-mediated enhances insulin sensitivity appears to be related to the activation of AMPK and PPARα, inducing an increase in fatty acid oxidation in both muscle and liver, and an increase of muscle glucose uptake. Therefore, due to an increase of IRS-phosphorylation and adiponectin level, bitter melon stimulates insulin sensitivity. Further study should be confirmed on PI3K pathway to analyze glucose uptake and the insulin-stimulated Glut-4 . Moreover, AMPK plays an important role in regulation of cellular energy levels. AMPK activation in muscle increases glucose uptake and fatty acid oxidation. The biological function of bitter melon is associated with the increased activity of AMPK. Therefore, bitter melon might be a good metabolic regulator (resulting in lowering TG level) and insulin sensitizer. Further studies are needed to shed light on the molecular mechanism involved in the p38 mitogen-activated protein kinase (p38MAPK) pathway activation of cell surface GLUT4. All of these findings point to at least two pathways which are implicated in bitter melon improved insulin resistance, and increased muscle glucose uptake resulting in a decrease of glucose levels [93].
However it was clearly demonstrated that bitter melon improved the expression and protein synthesis of GLUT4 in skeletal muscle, and prevented insulin resistance and hyperglycemia. Several approaches like pharmacological therapeutics, such as PPARγ have been shown to increase GLUT4 gene expression or plasma membrane translocation, contributing to improving glucose homeostasis. It was suggested that the effects of PPARγ on glucose homeostasis are caused by the induction of signaling molecules in adipose tissue, which indirectly would cause an improvement in muscle glucose disposal [94]. Similar effects of bitter melon on PPARγ mRNA to those observed in the previous study were shown in high-fat fed mice. Thus, it is possible that the notable improvements in glycemic control of bitter melon to increase the mRNA expression of GLUT4 in skeletal muscle might be attributable to be through PPARγ-mediated pathway. The increasing efficiency of glucose uptake can suppress elevated plasma glucose. Therefore, an increased GLUT4 protein in the plasma membrane of skeletal muscle seems to be responsible for the favorable impact on insulin sensitivity and GLUT4 mRNA level did not correlate well with protein content. In some tissues, post-transcriptional modulation may occur, leading to discrepant changes in the mRNA and protein content. However, the beneficial effects of two different extracts of bitter melon in rats fed a high-fructose diet thereby producing evidence of the role of changes in expression of PPARγ and GLUT4. The exact molecular mechanism involved in the enhancement of GLUT4 expression and protein by bitter melon remains to be elucidated [85].
Camellia sinensis (Green tea):
The polyphenols of green tea, namely catechins, have been reported to have multiple beneficial health effects, including a role in the treatment of hyperlipidemia and diabetes However, the molecular mechanisms that mediate these health benefits remain essentially unknown [95]. Studies were designed to investigate the effects of a green tea catechin-enriched extract on lipid and glucose homeostasis and decipher its probable mode of action by using the fructose-fed hypertriglyceridemic insulin-resistant golden Syrian hamsters as animal model. This model seems to be very appropriate because of the similarities between hamster and humans with respect to the major features of lipid metabolism and atherosclerosis. Hamsters fed the high-fructose diet for 2 weeks developed hypertriglyceridemia and hyperinsulinemia, while plasma glucose levels remained essentially unchanged [96]. The higher plasma glucose levels during the GTT also indicated the establishment of insulin resistance in the fructose-fed group. In addition serum adiponectin levels decreased after 7 weeks of fructose feeding. Adiponectin is an adipose-specific plasma hormone that is thought to be linked to the development of type 2 diabetes and obesity. It has also been shown to possess anti-atherogenic properties. The effect of a commercially available green tea EGCG-enriched extract was tested [97].
Interestingly, supplementation of the green tea extract for 4 weeks reversed the effects on all of the above parameters to near-baseline control values. Plasma TG values decreased dose-dependently, while cholesterol levels did not change. Furthermore, insulin, adiponectin, and apoB levels also reversed to levels comparable to the baseline control group. Plasma glucose levels also reduced during the GTT to levels comparable to the chow-fed group. Oral administration of a green tea extract is capable of improving glucose and lipid metabolism in an insulin-resistant hamster model induced by a high-fructose diet [98]. This confirms a number of other investigations comparing the effects of various types of tea extracts (1%, w/v; 3-4 weeks) on sucrose-induced hyperlipidemic male Sprague–Dawley rats. All teas, including green tea, normalized sucrose-induced hypertriglyceridemia and hepatic TG content. Wu et al. have examined the effects of green tea supplementation (0.5%, w/v; 12 weeks) on lipids and glucose homeostasis using a fructose-induced hypertriglyceridemic, insulinresistant rat model [98]. Green tea supplementation was shown to significantly improve the fructose-induced hypertriglyceridemia and glucose intolerance. For adiponectin, the results confirm a study in which the intake of tea was shown to increase plasma adiponectin levels in patients with type 2 diabetes [99]. There is strong evidence to support that fat diversion from adipose to non-adipose tissue, such as liver and muscle, which is not adapted to TG storage, leads to insulin resistance and type 2 diabetes. The clinical condition has been described as a lipotoxic disease. When TG over-accumulates in nonadipose tissues, fatty acids enter deleterious non-oxidative pathways such as ceramide and diacylglycerol production, and compromises cellular function. Green tea showed that TG content in both liver and heart muscle, but not skeletal muscle or adipose tissue, were reduced after extract supplementation in various animal models of insulin resistance [100]. The finding provides for a plausible link as to how green tea may improve insulin resistance. In addition, green tea would reduce body and tissue weight. A study indicated that tea catechins suppressed adipocyte differentiation in 3T3-L1 cells and a decrease of body weight gains in rats consuming tea has been also reported [101].
Since tea extract was shown to mediate responses on blood and tissue lipids, glucose metabolism, and body weight, the involvement of peroxisome proliferator-activated receptor was studied. PPARs are ligand-dependent transcription factors of which four isotypes (i.e. α, β, γ, δ) have been identified. They have been implicated in the control of energy homeostasis and obesity-related metabolic diseases. Once activated by a ligand, they control a variety of genes in several metabolic pathways. Much of the functions of PPARs have been associated with genes which participate in various aspects of lipid metabolism such as fatty acid uptake, fatty acid oxidation, lipolysis, and lipoprotein assembly and transport and glucose metabolism such as glucose production and disposal. Thus, the therapeutic potential of PPAR ligands has been realized with the advent of synthetic ligands for both PPARα (fibrates) and PPARγ (thiazolidinediones, also known as glitazones) in the treatment of the metabolic syndrome. Tea catechins may also represent PPAR ligands of the natural type. Interestingly, both hepatic PPARα and PPARγ protein expression increased dramatically upon green tea extract supplementation relative to fructose-fed control. This suggests that tea catechins act as PPAR ligands with a dual alpha/gamma agonistic effect. This finding may provide a molecular mechanism for the beneficial effects seen on blood lipid/glucose levels, reversal in hepatic and myocardium lipotoxicity, and reduction in body weight and liver/adipose tissue in diabetes [102,103].
Cinnamomum species
The current pharmacological treatments including use of thiazolidinediones (TZDs) for the treatment of insulin resistance have undesirable side effects such as weight gain, fluid retention, the worsening of coronary heart disease and an increased risk of myocardial infarctions [103]. However, medicinal plants and purified compounds are expected to have a similar degree of efficacy without the troublesome side effects associated with conventional drug treatment. Moreover, beneficial effects of cinnamon polyphenols including flavonoids have been reported in humans, rats and cell culture studies. However, the anti-diabetic activity of characteristic chemical constituents of Cinnamomum osmophloeum (C. osmophloeum), is not well defined. Only one kaempferol diglycoside kaempferitrin was reported to activate the insulin signaling pathway and stimulate secretion of adiponectin in 3T3-L1 adipocytes. Therefore, the insulin-like effects of two flavonol glycosides from C. osmophloeum leaves (CO–1 and CO–2), which are structurally different from previously reported kaempferitrin, were investigated on three of the proteins (adiponectin, IRβ and GLUT4) involved in insulin signal transduction pathway using mouse 3T3-L1 adipocytes [104].
Adiponectin is an adipocytokine secreted solely by adipose tissue and has been shown to have both anti-diabetic and anti-atherogenic properties. It has attracted much attention because of its decrease in the circulating levels causing to the development of obesity, insulin resistance, type 2 diabetes and atherosclerosis. Therefore, adiponectin is regarded as being a crucial tool for the diagnoses of type 2 diabetes. Effective C. zeylanicum dose of 0.2 mg/ml increased glucose uptake, but concurrently inhibit adiponectin secretion from the 3T3-L1 adipocytes [105]. The flavonol glycoside CO-1 was isolated from the leaves of an endemic tree species C. osmophloeum, therefore its adiponectin secretion property may different from those of C. zeylanicum. Furthermore, it should be noted that the well-known anti-diabetic drugs TZDs shown to have insulin-like activities through increase adiponectin secretion in 3T3-L1 adipocytes. Thus, CO-1 has in line effect of these known anti-diabetic drugs in terms of adiponectin secretion. It is previously reported that CO-1 with anti-inflammatory function through the down-regulation of pro-inflammatory mediators such as nitric oxide, and cytokines tumor necrosis factor (TNF)-α and interleukin (IL)-12 in macrophage cells. Further, obesity and type 2 diabetes is associated with increased levels of adipokines including TNF-α and decreased levels of adiponectin [106]. The insulin signaling pathway leading to the cellular uptake of glucose begins with the binding of insulin to its receptor (IR); in the sequence, ligand binding to the IR induces autophosphorylation of specific tyrosine residues, which then activates the substrate protein tyrosine kinase of the IR. The data show that compounds CO-1 at 5 μM and CO-2 at 20 μM significantly enhances the tyrosine phosphorylation level of IRβ and suggests a beneficial role of these compounds in insulin signaling. Further, low concentrations of CO-2 did not exhibit a significant effect on IRβ levels, which is in agreement with a previous report that there is not a significant difference in IRβ levels between rats fed with cinnamon and the control rats fed with saline. GLUT4 is the major glucose transporting molecule in muscle and adipose tissue. It is known that insulin promotes translocation of GLUT4 from intracellular compartment to plasma membrane [107].
Although, insulin-stimulated adiponectin level and insulin receptor phosphorylation and promote GLUT4 translocation, however, previous reports suggest that adiponectin and other two markers are not necessarily responded similarly to the tested compounds. Apart from insulin, PPAR-γ ligands, a number of hormones including testosterone, prolactin, glucocorticoids and growth hormone, inflammation and oxidative stress in adipose tissue, a number of pharmacological agents (angiotensin receptor type 1 blockers, ACE inhibitors, and cannabinoid receptor antagonists) also regulate the level of adiponectin. These regulators do not stimulate insulin receptor phosphorylation and GLUT4 translocation. Furthermore, the mechanism of insulin regulating adiponectin level is not still clear. It is believed that adiponectin level is raised with the feedback mechanism activated when high level of insulin is present to induce insulin resistance [108]. An example of adiponectin regulated separately to insulin receptor phosphorylation and GLUT4 translocation is seen when the adipocytes are stimulated with vanadium. When stimulated with vanadium, cellular content of adiponectin from adipocytes was increased, but this up-regulation is not accompanied with increased level of phosphorylated IR. This result indicated that the signaling cascade involved in adiponectin accumulation does not depend on the activation of IR. In other words, level of adiponectin could be up-regulated independently of insulin receptor. Addition of PI3-K inhibitor results on further increase of adiponectin. This lack of an inhibitory effect suggests that PI3-K is not positively involved in mediating insulin induced adiponectin accumulation [109]. In a clinical study, oral administration of cinnamon (Cinnamomum cassia) with 2g cinnamon powder filled in capsules (4 capsules/day/person) for 40 days on non-insulin dependent diabetes mellitus patients showed that at initial level fasting blood glucose of experimental group was 148.73 ± 3.69 mg/ dl which reduced to 134.0 ± 3.12 mg/dl after 20th day and further reduced to 120.66 ± 4.70 mg/dl at the completion of the study. Post prandial blood sugar also reduced significantly from 187.66 ± 3.29 mg/dl (0 day) to 172.93 ± 3.51 mg/dl (20th day) and again reduced to 163.6 ± 5.09 mg/dl after 40 days of intervention in the experimental group [107,110].
In this study a critical analysis of the effect of Momordica charantia, Camellia sinensis and Cinnamon species on insulin resistance has been performed. These plants have been clearly shown to exhibit antihyperglycemic effect on different animal models. In addition, in order to elucidate the mechanism of action involved in their pharmacological activity, many studies have been performed and a clear effect on insulin resistance has been demonstrated (Figure 1).
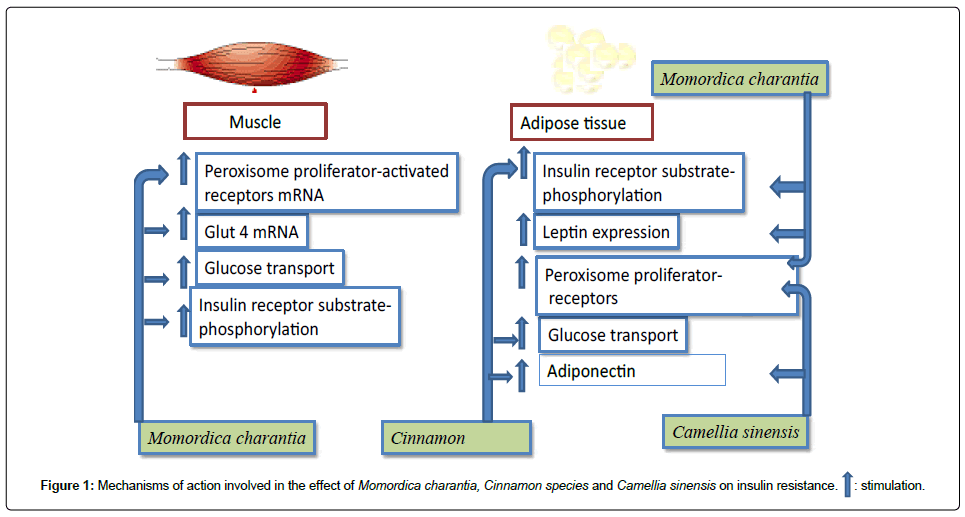
Figure 1: Mechanisms of action involved in the effect of Momordica charantia, Cinnamon species and Camellia sinensis on insulin resistance. : stimulation.
: stimulation.
It appears from the cited examples that plants can effectively reduce insulin resistance throughout different mechanisms. However, little is known about the effect of these plants under human insulin resistance. In addition, phytochemical investigation is extremely needed to identify and safely uses of phytotherapy in a large scale.
This work has been supported by the “MESRSFC: Appel à projets dans les domaines prioritaires de la recherche scientifique et du développement technologique (Call for projects in the priority areas of scientific research and technological development).”