Fisheries and Aquaculture Journal
Open Access
ISSN: 2150-3508
ISSN: 2150-3508
Research Article - (2010) Volume 1, Issue 1
This study was undertaken to define the effects of temperature on the energy and protein partial utilisation efficiencies of juvenile Barramundi. The experiment used a factorial design with four temperatures (25ºC, 29ºC, 32ºC, and 36ºC) and three ration levels (low, moderate, satiety) to examine the response of Barramundi to varying digestible energy (DE) and digestible protein (DP) intake. Energy and protein deposition with varying intakes at most temperatures were linear, though aberrations occurred at 36ºC relative to the other temperatures. The coefficients of DE utilisation were relatively consistent at 0.56 ± 0.02 (mean ± SEM) between 25ºC and 32ºC, though at 36ºC this declined to 0.42 ± 0.04. Similarly the maintenance DE demand for the fish was relatively constant across the range 25ºC to 32 ºC (~40 kJ DE/metabolic body weight (MBW)/d), but at 36ºC dramatically increased to around 110 kJ DE/MBW/d. The coefficients of DP utilisation were also relatively consistent at 0.51 ± 0.02 between 25ºC and 32ºC, though at 36ºC this declined to 0.28 ± 0.12. Similarly, the maintenance DP demand at 36ºC dramatically increased from around 0.5 g DP/PBW/d to 1.5 g DP/PBW/d. These results demonstrate that at high temperatures Barramundi protein demand and utilisation is significantly compromised and this affects their ability to efficiently convert dietary protein to tissue growth.
Keywords: Asian seabass, heat, temperature, stress, energetics, bioenergetics.
The production of Barramundi (Lates calcarifer), also known as Asian seabass, in Australia occurs in water temperatures ranging from 18°C to 36°C [1]. Growth rates of Barramundi are optimal at around 30°C – 32°C. Above this temperature, the growth rate plateaus before declining rapidly at temperatures exceeding 35°C. Death occurs at around 38˚C – 40°C and larger fish appear to be more sensitive to higher temperatures than smaller fish [2]. Symptoms of thermal stress above 35°C include slower growth rates, increased mortality, increased level of cataract formation, reduced rates of protein synthesis and increased levels of endogenous protein turnover [2-4]. Temperature stress has been shown to have a significant effect on the total maintenance energy and protein losses of Barramundi during periods of starvation and on the relationship between animal size and energy or protein demands for maintenance [5].
One way to examine the implications of thermal stress on fish is to model its effects using a factorial bioenergetic approach [5,6]. Bioenergetic factorial models are an empirical model form that compartmentalizes the energy flows into either somatic (growth) or non-somatic (maintenance) components. In simplistic terms, the total energy demand model can be generally summarised as:
Total Energy Demand (kJ/fish/d) = ME * (Liveweight)b + GE * Energy gain
Where ME and GE are parameters describing the efficiency of energy utilisation for maintenance and growth respectively and b is the metabolic weight exponent of the animal [5,6]. The total protein demand can also be estimated in a similar fashion, but with the energy terms substituted with protein terms [5,6]. These models typically rely on a suite of empirically determined parameters (i.e. coefficients and exponents) relating to energy/protein flux, with the mathematical linking of these parameters forming the basis of the working bioenergetic model. However, like most simplistic models the determination and subsequent use of the model parameters are based on a range of assumptions. One of the key assumptions of many of the bioenergetic models being used in fish nutrition is that the partial efficiencies of energy and protein utilisation above maintenance demand are not affected by temperature.
Temperature has been shown to have a profound effect on many of these model parameters. The demands for maintenance increase with temperature and as such alter the ME coefficient [5]. While an exponent of ~0.80 appears to be typical for Barramundi and most other fish species when fish are reared within their normal thermal range [5,6], it has been observed that above normal temperatures this exponent dramatically increases and an exponent of 0.80 is no longer applicable. The growth potential (Energy gain) of the fish is also clearly affected by temperature [2,5].
A number of studies have determined coefficients of energy utilisation that typically range between 0.61 and 0.76 [5,6]. Similarly, coefficients for protein utilisation have also been determined, but the range in these values appears to be much greater, with values between 0.23 and 0.55 being reported [5-7]. There is a range of factors that probably affect these utilisation coefficients (e.g. diet protein amino acid composition, diet protein-energy-balance, among others). However, we hypothesise that under conditions of high temperature stress, the utilisation of protein and energy by Barramundi will alter, consistent with deterioration in the rates of protein synthesis and increased rates of protein turnover. This will most likely lead to a decrease in the coefficients of protein and energy utilisation, as the animals enter a heat stress state.
Therefore, the aim of this study was to examine the effect of water temperature on the partial efficiencies of energy and protein utilisation in juvenile Barramundi. In order to study these effects we subjected groups of Barramundi to four experimental temperatures which bracketed the lower, optimal and upper thermal temperature range of this species (i.e. 25°C, 29°C, 32°C or 36°C) and fed them a range of feed ration levels. The effect of temperature on the capacity of Barramundi to convert dietary energy and protein into tissue protein and energy stores and the underlying assumptions about the effect of temperature on the application of bioenergetic factorial models is discussed.
Experimental feeds
An experimental diet was extruded using an APV 19:25 laboratory-scale twin-screw extruder fitted with a 2.4 mm diameter pellet die. Following extrusion, the pellets were oven dried at 60C for approximately 12 h. After drying and whilst the pellets were still warm, they were vacuum infused with fish oil (Table 1). The formulation, composition and source of ingredients for the experimental diet are presented in Table 1.

Table 1: Nutrient composition of the experimental diet.
Fish handling and trial preparation
This study used hatchery-reared Barramundi (Lates calcarifer) that were pre-grown to an acclimation weight of around 15 g in indoor 1000L heated seawater tanks (29˚C). The experiment was run in two blocks (i.e. events) designed to include 4 temperatures (25°C, 29°C, 32°C or 36°C) x 3 ration levels (30% of apparent satiation, 60% of apparent satiation or apparent satiation) duplicated within each blocking event (i.e. 24 tanks per block). For each blocking event, an acclimation period was used where the pre-grown fish were split into four temperature groups of two tanks containing approximately 100 fish each. Each holding tank was supplied with pre-heated and continuously aerated seawater at a flow rate of 4 L/min. The temperature of each holding tank was adjusted or maintained using a computer (PLC) that controlled the mixing of 20°C and 40°C seawater in a mixing chamber prior to the water being distributed to each tank. One group of tanks was maintained at 29°C consistent with the original temperature at which the fish were grown, while the other groups of tanks had their water temperature altered by 2°C every 72 h over an eight day period to a maximum temperature of 36°C or to a minimum of 25°C consistent with those methods used in Bermudes et al [2]. Prior to all weighing procedures, fish were sedated using isoeugenol at 0.002 mL/L (supplied as AQUI-S™) and individually weighed to 0.1 g. The fish were then allowed to regain consciousness and equilibrium before being placed within their designated experimental tank.
Growth assessment
Following the acclimation period, 18 fish were each randomly allocated to six tanks within their respective temperature block Mean ± SD initial weights for block 1 and block 2 were 14.5 ± 0.53 g/fish and 13.1± 0.26 g/fish, respectively with an overall initial weight of 13.8 ± 0.81 g/fish. Temperatures for each treatment within each block were determined using data loggers that recorded temperature every hour for the duration of the trial. The recorded mean±SD temperature in each treatment array was: Block 1; 25.4± 0.06°C, 29.3± 0.02°C, 32.7± 0.07°C, 36.4± 0.07°C and Block 2; 26.2± 0.26°C, 29.5± 0.11°C, 32.9± 0.04°C, 36.5± 0.17°C. Block 1 was run for 24 days and Block 2 was run for 22 days. At the end of each blocking period all fish were individually weighed to determine the mean weight of fish from each tank. The mean value for each tank was used as the experimental replicate unit.
Feed intake assessment
Treatments allocated to the satiety ration were fed to apparent satiety once daily between 0900 and 1100 hours for the duration of the experiment. Fish fed to satiation were fed in three sessions over a 1-hour period with apparent satiety determined by a loss in feeding activity. The average daily amount of feed consumed by the satiated group was subsequently used to calculate sub-satietal rations for groups of fish allocated to the 30% or 60% ration treatments. Any uneaten feed was removed from each tank one hour later and the uneaten portion dried and weighed to allow the determination of daily feed intake based on correction factors for leaching losses sustained over an equivalent period [9].
Sample processing and chemical analysis
All chemical analyses were carried out by NATA (National Association of Testing Authorities) accredited analytical service providers (Chemistry Centre, East Perth, WA, Australia; Animal Health Laboratories, South Perth, WA, Australia; SARDI Pig and Poultry Production Institute, Roseworthy, SA, Australia). Whole fish samples (minimum of three fish per tank, starved for a minimum period of 24 h prior to sampling) were analysed for dry matter, protein (as nitrogen) and energy content. Dry matter was calculated by gravimetric analysis following oven drying at 105°C for 24. Protein levels were calculated from the determination of total nitrogen by LECO auto-analyser, based on N x 6.25. Gross energy was determined by adiabatic bomb calorimetry of the samples.
Diet samples were analysed for dry matter, nitrogen, total lipids, ash, amino acids and energy content. All analyses were performed in accordance with the methods specified by AOAC International (2005). Amino acid analysis involved the samples being hydrolysed at 110°C for 24hr in 6M HCl with 0.05% Phenol. Cystine was derived during hydrolysis by the addition of 0.05% 3-3-dithiodipropoinic acid. The acid hydrolysis destroyed tryptophan making it unable to be determined. Separation was by HPLC on a Hypersil AA-ODS 5m column using an 1100 series Hewlett Packard HPLC system. Total lipid content of the diets was determined gravimetrically following extraction of the lipids using chloroform:methanol (1:1). The proximate composition of the diet was as reported in Table 1 [2].
Protein and energy utilisation analysis
Protein (N x 6.25) and energy (E) utilisation in Barramundi was determined based on the gain in nitrogen or energy content of the tissue stores over the period of the experiment relative to the respective consumption of digestible nitrogen or digestible energy over the same period. Digestible nitrogen and energy intakes were determined by multiplying total feed intake values by the apparent digestibility coefficients of nitrogen or energy determined for a previously used experimental diet [2] at each respective temperature. Protein or energy gain and protein or energy intake values were calculated based on the amount gained or consumed per unit of body weight. To provide some independence of size effects, modeling of the protein and energy utilization data was done after referring intake and consumption data to energy (MBW) and protein body-weight (PBW) exponents determined for Barramundi at each of the experimental temperatures. These scaling exponents are derived from equations presented in Table 2.

Table 2: Performance of juvenile barramundi reared at different temperatures and fed different rations.
Statistical analysis
All values presented in tables and figures are means unless otherwise specified. All data were analysed for homogeneity of variances using Cochran’s test. Effects of temperature and ration were analysed using a 2-factor multiple analysis of variance procedure (MANOVA; Statistica, Statsoft™, Tulsa, OA, USA). Where appropriate, significance of differences were examined using a Fishers LSD test based on critical ranges being set at P < 0.05. Regression coefficients and constants were also calculated using Statistica software while figures were plotted using MS Excel (Version XP, Microsoft Corporation, USA).
Growth and feed utilisation
The growth and feed intake observed of the fish in this study showed a clear significant effect of both temperature and ration (Table 2). Satietal feed intake by fish was lowest at 25°C, though only marginally so than those fish fed at 36°C. Feed intake was highest at 32°C.
Growth was highest in those satietal fed fish at 32°C, though this was not significantly better than those at 29°C. When feed intake was kept constant, there were some significant differences in gain achieved among the different temperatures. Those fish kept at 29°C had the lowest feed conversion, gaining more with the same level of intake than those fish at the other temperatures.
Energy utilisation
The relationship between DE intake and energy deposition at each temperature was fitted to a separate linear function (Figure 1; Equations 1 – 4a). There was some indication that data for the 36°C treatment could be fitted to a logarithmic response (i.e. Equation 4b), but as it was our intention to compare the partial utilisation efficiency of energy at each temperature we restricted our examination to that using a linear response. The partial efficiency of utilisation of DE as defined by the regression coefficient was observed to increase with temperature before peaking at 32°C then dramatically declining at 36°C (Figure 3a; Equation 5). The maintenance demands for DE, as defined by the x-intercept at zero energy gain for each temperature was relatively constant below 32°C ranging between approximately 34-39 kJ/MBW/d (Figure 3b; Equation 6). However, use of a linear function to describe the maintenance DE demand at 36˚C resulted in a positive y-intercept, which returned a non-sensical value for maintenance DE demand. To circumvent this problem, the logarithmic function described by Equation 4b was used to identify the x-intercept or maintenance DE demand at this temperature (Figure 1; Equation 4b). This value was calculated as 110kJ/MBW/d.
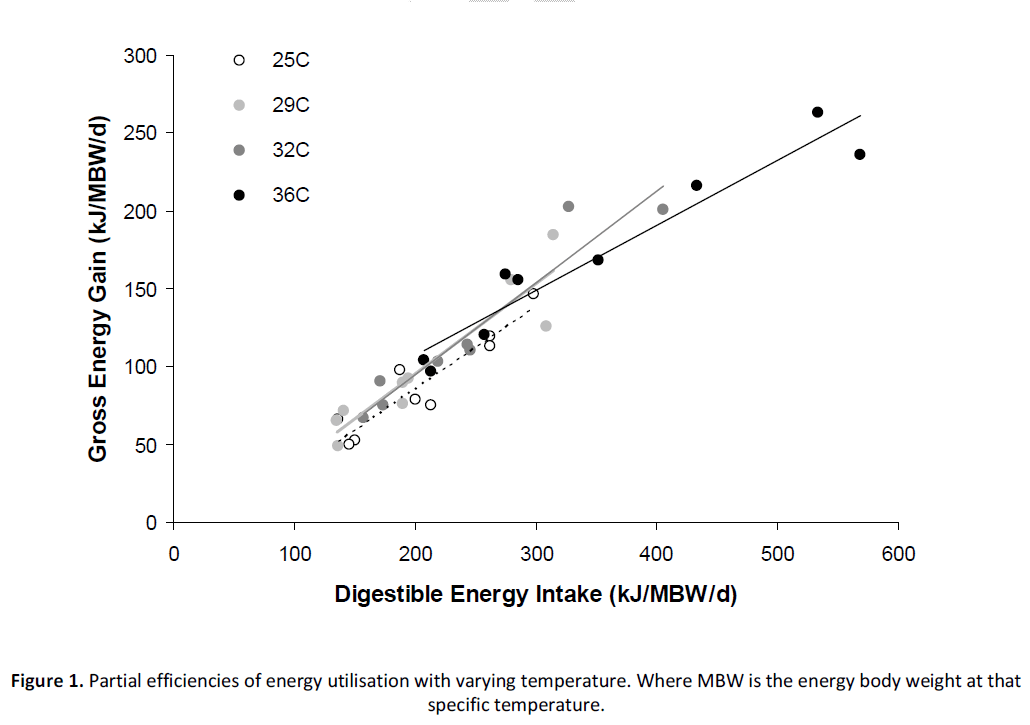
Figure 1: Partial efficiencies of energy utilisation with varying temperature. Where MBW is the energy body weight at that specific temperature.
Eq 1. Energy gain @ 25°C (kJ/ kg0.80/d) = 0.529(±0.076) * DE intake – 20.358(±16.240), R2 = 0.87
Eq 2. Energy gain @ 29°C (kJ/ kg0.79/d) = 0.576(±0.087) * DE intake – 20.018(±19.153), R2 = 0.86
Eq 3. Energy gain @ 32°C (kJ/ kg0.82/d) = 0.590(±0.074) * DE intake – 22.981(±18.887), R2 = 0.91
Eq 4a. Energy gain @ 36°C (kJ/ kg0.91/d) = 0.418(±0.049) * DE intake + 23.513(±17.130), R2 = 0.93
Eq 4b. Energy gain @ 36°C (kJ/ kg0.91/d) = 152.2 * Ln(DE intake) – 711.86, R2 = 0.95
Eq 5. DE utilisation coefficient = 0.0005*temp3 + 0.0442*temp2 – 1.223*temp + 11.691, R2 = 1.00
Eq 6. Maintenance DE demand (kJ/MBW/d) = 0.185*temp3 – 15.561*temp2 + 434.4*temp + 3983.7, R2 = 1.00
Protein utilisation
The relationship between protein intake and protein deposition at each temperature was also fitted to linear functions (Figure 2; Equations 7 – 10a). As with energy utilisation, there was an indication that protein retention followed a logarithmic response at 36°C (i.e. Equation 10b). As for DE utilisation, the logarithmic function was used to calculate maintenance DP demand at this temperature. The partial efficiency (coefficient) of utilisation of DP was 0.55 at 25°C and declined to approximately 0.49 at 29˚C and 32°C. The coefficient declined to 0.28 at a temperature of 36˚C (Figure 4a; Equation 11). When plotted against temperature, the maintenance demands for DP, as defined by the x-intercept at zero protein gain are lowest at 32°C, with DP demand elevated at lower or higher temperatures, particularly at 36˚C (Figure 4b; Equation 12).
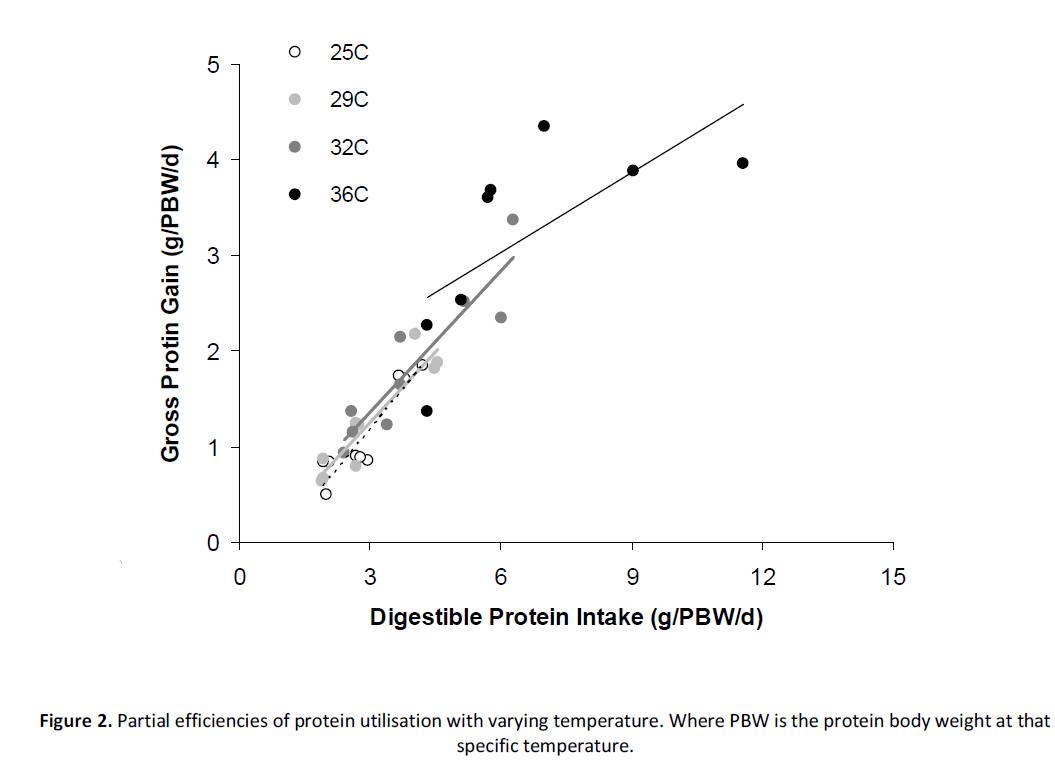
Figure 2: Partial efficiencies of protein utilisation with varying temperature. Where PBW is the protein body weight at that specific temperature.
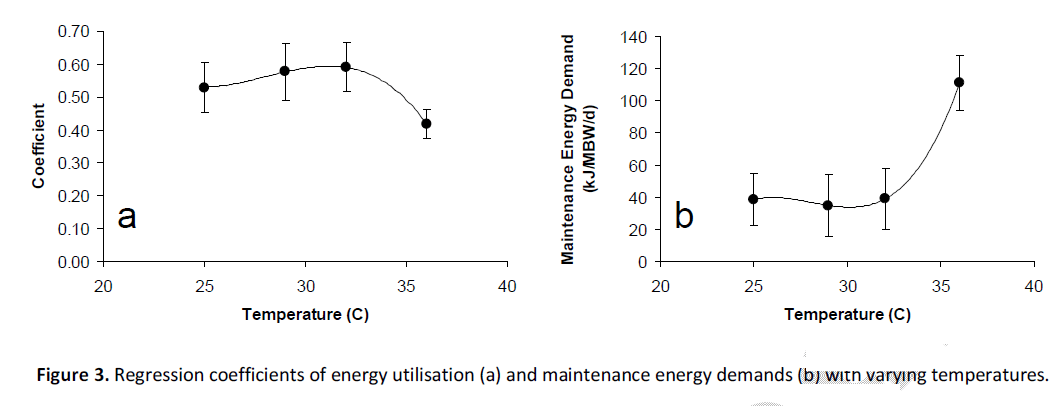
Figure 3: Regression coefficients of energy utilisation (a) and maintenance energy demands (b) with varying temperatures.
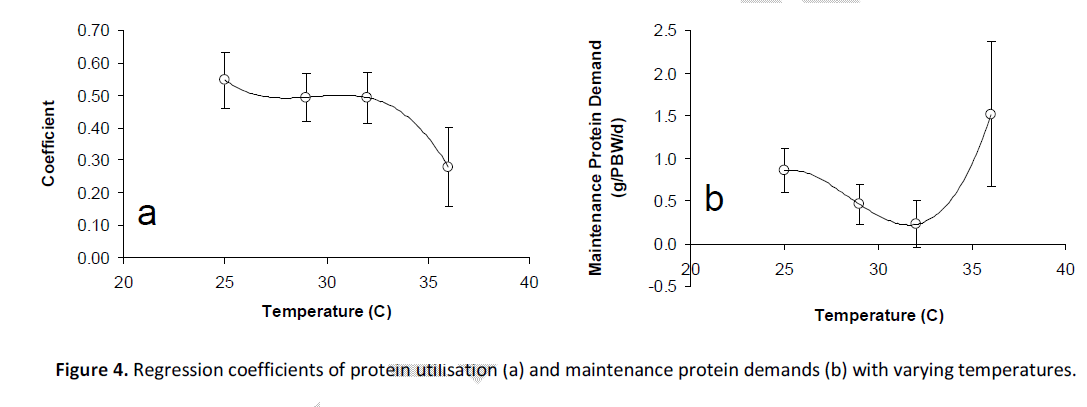
Figure 4: Regression coefficients of protein utilisation (a) and maintenance protein demands (b) with varying temperatures.
Eq 7. Protein gain @ 25°C (g/ kg0.69/d) = 0.546(±0.085) * DP intake – 0.470(±0.258), R2 = 0.85
Eq 8. Protein gain @ 29°C (g/ kg0.67/d) = 0.493(±0.074) * DP intake – 0.227(±0.235), R2 = 0.86
Eq 9. Protein gain @ 32°C (g/ kg0.72/d) = 0.493(±0.068) * DP intake – 0.115(±0.268), R2 = 0.85
Eq 10a. Protein gain @ 36°C (g/ kg0.89/d) = 0.279(±0.121) * DP intake + 1.361(±0.851), R2 = 0.47
Eq 10b. Protein gain @ 36°C (g/ kg0.89/d) = 2.267 * Ln (DP intake) – 0.946, R2 = 0.59
Eq 11. DP utilisation coefficient = 0.0009*temp3 = 0.0759*temp2 – 2.2245*temp = 22.19, R2 = 1.00
Eq 12. Maintenance DP demand (g/PBW/d) = 0.049*temp3 – 0.421*temp2 + 11.84*temp – 108.9, R2 = 1.00
This study aimed to establish the effect of different water temperatures on the partial efficiencies of energy and protein utilisation by juvenile Barramundi. These parameters, represented by the regression coefficients determined for energy or protein utilisation, respectively, are essential components of bioenergetic models predicting fish growth and feed utilisation [10].
Effect of temperature on energy utilisation
The growth achieved by the fish in this study was consistent with that expected of fish of this size at the water temperatures encountered during the growth study period [2,5]. Feed conversion across all temperatures was good, being less than 0.80: 1, but lowest at 29°C and highest at 36°C.
Poorer growth and feed utilisation in Barramundi reared at high temperatures (>35°C) has been reported by several authors [2,4,11]. However, there are few reports on the energetic implications of these changes with temperature stress [2]. At normal physiological temperatures (25°C – 32°C), the coefficient of partial efficiency of energy utilisation was observed to be 0.523 – 0.590, which is slightly lower (poorer) than that reported [5] for Barramundi fed the same diet (0.611 – 0.763). By comparison, the partial efficiency of energy utilisation by fish in the present study was significantly impaired at 36°C (i.e. 0.418 cf. mean25°C – 32°C of 0.564) relative to that recorded at the lower water temperatures. This response provides clear evidence of a reduction in the energetic efficiency of Barramundi at temperatures, which exceed their optimal physiological range. It has also been showed that feed efficiency ratio, productive protein value (PPV) and productive energy value (PEV) in small Barramundi (4-20g fish) was significantly poorer at 39°C compared to 27 – 36°C, consistent with what we report here in terms of a reduction in the partial efficiencies of utilisation of energy from 0.56 to 0.42 at 36°C [11]. However, the use of larger fish in the present study also demonstrates the potential to better define effects of heat stress than those effects seen on smaller fish, similar to that seen in other studies with larger fish [2].
Using linear regression to determine the partial efficiency of utilisation of energy (or protein) is a method that has been demonstrated to have substantial merit in terms of defining the energetics of growth and food utilisation in fish [12], but, there is continuing debate over whether the relationship between energy/protein intake and energy/protein deposition is linear or curvilinear [6-8]. Apart from fish reared at 36˚C, our data was consistent with a linear approach. However, unlike many studies our data was referred to predetermined metabolic weight exponents for each temperature regime prior to fitting individual regressions. One of the ironies of this approach is that when data is plotted on the same axes (Figures 1 & 2), Barramundi reared at 36˚C appear to exhibit higher DE intake and energy deposition rates than fish reared at more optimal temperatures even though their respective feed intake and growth rates were lower (Table 2). This anomaly can be attributed to the different metabolic body-weight exponents used on each temperature regression (~0.80 cf. 0.91; Table 2) and how this transforms the data. Therefore, while the allometric data shows higher activity, in reality it is comparatively lower on an untransformed basis. Another notable point was the loss of linearity in the 36°C data relative to the other temperatures. Use of a logarithmic curve allowed the estimation of the maintenance energy demand at this temperature, but its use also implies that the partial efficiency of energy utilisation is changing with energy intake level. This is consistent with reports in other studies that suggest that the relationship between energy intake and energy deposition is not always linear [5,7].
The maintenance energy requirements seen in the present study compare well with those documented by others working with this species [2,5]. At temperatures of 25°C – 32°C the maintenance energy demand was close to 40 kJ/ kg0.80/ d, not dissimilar to the 42.6 kJ/ kg0.80/d estimated in other studies [5]. However, at 36°C this dramatically increased to ~110 kJ/ kg0.91/ d. This means that more than double the amount of energy would be required to be consumed to maintain maintenance energy demands by this species at this temperature. So even with an increase in feed intake, the animal suffers an increasing proportion of its energy intake being diverted to maintenance requirements rather than growth. This explains the reduction in feed use efficiency seen at elevated temperatures [2,11].
The effects seen on energy utilisation can be largely explained by the changes seen in protein utilisation in this species at the higher temperatures. The partial efficiency of utilisation of protein at normal physiological temperatures (25°C – 32°C) was observed to be 0.493 – 0.546, which is similar to that reported for the same species (but of different sizes) fed the same diet (0.484 – 0.496) at 30°C [5]. However, at 36°C the partial efficiency of protein utilisation by the fish in the present study was demonstrated to be significantly impaired (0.279 cf. mean 25°C – 32°C of 0.510). This provides clear evidence of a reduction in the protein efficiency of this species during excessively high temperatures and this reduction in protein utilisation efficiency largely accounts for the changes seen in energy retention efficiency.
The dramatic increase in maintenance protein requirements seen in this study contrasts those seen at normal physiological temperatures (Figure 4). The maintenance protein requirements seen at normal physiological temperatures in the present study compare well with those documented by other studies on Barramundi [5]. At temperatures of 25°C – 32°C the maintenance protein demand ranged between 0.22 and 0.86 g/ kg0.70/ d (mean = 0.51 ± 0.18 g/ kg0.70/ d), not dissimilar to the 0.45 g/ kg0.70/ d estimated in earlier studies [5]. However, at 36°C the maintenance demand for protein increased to 1.52 g/ kg0.89/ d. This means that almost three times the amount of protein would be required to be consumed to maintain maintenance protein demands by Barramundi at this temperature.
Other workers have also identified similar escalations in relative protein demand in other species [3,11,13]. It was reported that significant effects were seen on protein degradation pathways in both Atlantic Salmon and Barramundi at high temperatures and that both the autophagic-lysosomal and ubiquitin-proteasome pathways were involved [3]. However, it was also showed no significant differences between 27°C and 33°C in terms of protein synthesis or degradation, but had they examined higher temperatures they might have observed significant reductions in synthesis and increases in degradation consistent with what is reported in terms of protein utilisation efficiency and maintenance protein demands at 36°C in the present study [14]. In earlier work it was shown that at 39°C the protein efficiency ratio was significantly reduced compared to 27 – 36°C [11]. Had larger fish been used in that study then effects at 36°C might too have been observed, as was seen with the present study. The use of very small fish in such thermodynamic studies also limits the potential of seeing thermal sensitivity effects with temperature variation as it is known that smaller fish are more tolerant of temperature variation than larger fish [2,15].
Several authors have examined the potential for dietary management of temperature related issues with Barramundi. The value of different dietary energy densities on the effect of temperature related energy conversion, albeit primarily intended to examine the potential for improving productivity at the lower end of the temperature spectrum (<20°C) was also evaluated [16]. These authors found that increasing energy density (while maintaining protein:energy ratio) resulted in improvements to both FCR and growth rate and that these effects were more pronounced at 20°C than 29°C.
Dietary management of the upper temperature ranges was explored in earlier studies and the use of diets with varying protein to energy ratios on the growth and protein synthesis of Atlantic Salmon was reported [17]. These authors found no benefit from increasing the protein to energy ratio for this species at 19°C. In contrast, the use of an increase in protein to energy ratio (27 vs 21 g/MJ) was found beneficial for reducing the impact of temperature stress on growth in Barramundi [4]. Although growth was still impacted at the higher temperature (36°C), it was not significantly reduced compared to the control temperature (30°C). By comparison, those fish fed a diet with a lower protein to energy ratio exhibited a significant decline in growth at 36°C.
The findings of this study demonstrate that with varying temperature that there can be significant changes in some of the key parameters of bioenergetic factorial models, long assumed to be constants. The implication of this is that these constants should be changed to a function that accommodates the effect of temperature on these parameters. Whilst within low to optimal temperature ranges there is little effect on these parameters, at high temperatures the significant change means that use of the constant values is not valid. The effects observed on these parameters at the higher temperatures also explain some of the features seen during heat stress with the higher protein turnover and reduced capability of the animal to utilise dietary protein for its own tissue synthesis [2,3]. Some further work on elements of this, examining what metabolic pathways are regulated, which specific proteins are catabolised during maintenance and what nutrient (amino acid) limitations there are to utilisation, would be useful to progress the development of diets to allow better management of high-temperature regimes.
Neither author has any competing interests with the publication of this work.
BG was involved in acquisition of funds to do this work, the conceptual design of the experiment, assisted the detailed design of the experiment, coordinated the analysis of the samples, did the calculations and analysis of the data and the writing of the manuscript. MB was involved in the detailed design of the experiment, the initiation and maintenance of animal trial, and contributed to the editing of the manuscript.
We would like to acknowledge the technical assistance of Kate Austen (nee Crass) and Trân Lê Câm Tú in the maintenance of this experiment. The analytical support of Ken Dods at the Chemistry Centre of WA and Kylee Swanson from the South Australian Research and Development Institute (SARDI) for performing some of the analytical work associated with this project is also acknowledged. We acknowledge the financial support of the Australian Centre for International Agricultural Research. This experiment was conducted with the approval of the Department of Fisheries animal ethics group. We also thank Dr Mark Booth for reviewing an early version of this work.