Journal of Proteomics & Bioinformatics
Open Access
ISSN: 0974-276X
ISSN: 0974-276X
Research Article - (2017) Volume 10, Issue 2
The identification of up- and downregulated as well as absent proteins in the central nervous system is necessary to understand the interplay of migration, differentiation and integration of neuronal progenitor cells at different stages of development. In a first step, differentially expressed proteins of the cerebral cortex of the laboratory rat at three significant stages of development were identified. The cerebral cortex needs differential abundances of proteins during ontogenesis and uses its high plasticity postnatally to adapt to many types of intrinsic and extrinsic changes. This study focuses on the identification of specific proteins which are differentially expressed during postnatal development. Cerebral cortices of P7, P90 and P637 old wistar rats were dissected and analyzed by twodimensional polyacrylamide gel electrophoresis (2DE) and matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF-MS) analysis. The identified and differentially expressed proteins are subdivided into 13 different classes. Proteins of the functional classes of the carbohydrate metabolism, structural and regulatory proteins as well as proteins involved in the energy metabolism show the highest differential abundance within the analyzed stages of development. Cytoskeleton proteins like neurofilaments and β-actin are downregulated in early development. In contrast, some proteins which are necessary for migration and motility are upregulated in P7 versus P90 animals. Furthermore, proteins for vesicular trafficking like drebrin and Gdi2 are upregulated in P7. In aged animals oxidative stress sensors, proteins necessary for autophagy of dysfunctional mitochondria, growth control and hypoxia tolerance (Ppp1ca, Eno1) turned out to be upregulated. Overall, energy consumption and differentiation processes as well as specific regulatory mechanisms can be observed at least indirectly by differential abundances of proteins during the investigated stages of ageing.
Keywords: Brain; Development; Cerebral Cortex; Proteomics; Rat
A: Axon; CBB: Coomassie Blue; CpG: Cytoplasmic Granule; CNS: Central Nervous System; Cts: Centrosome; Cr: Chromosome; CSK: Cytoskeleton; CP: Chaperones; CPM: Cytoplasm; CPV: Cytoplasmic Vesicle; CTS: Cytosol; Cx: Cerebral Cortex; ECM: Extracellular Matrix; ER: Endoplasmic Reticulum; Eds: Endosome; ExR: Extracellular Region; GC: Growth Cone; GJ: Gap Junction; Golgi: Golgi Apparatus; HGC: Heterotrimeric G-protein Complex; LA: Lipid Anchor; Lyso: Lysosome; M: Membrane; Micro: Microsome; MMT: Mitochondrial Matrix; MM: Mitochondrial Membrane; MIM: Mitochondrion Inner Membrane; MIMS: Mitochondrion Intermembrane Space; MOM: Mitochondrion Outer Membrane; MEL: Melanosome; Mito: Mitochondrion; MT: Microtubule; NC: Nucleus; NF: Neurofilament; NM: Nucleus matrix; NP: Nucleoplasm; P: Proteasome; PA: Proteins Antioxidants; PAAM: Proteins Amino Acid Metabolism; PB: Proteins Biosynthesis; PCM: Proteins Carbohydrate Metabolism; PD: Proteins Degradation; Per: Peroxisome; PEM: Proteins Energy Metabolism; PEMA: Proteinaceous Extracellular Matrix; PFM: Proteins Fat Metabolism; PMP: Peripheral Membrane Protein; PR: Proteins Regulation; PST: Proteins Signal Transduction; PTM: Proteins Transmitter Metabolism; RS: Ribosome; S: Synapse; Sc: Secreted; SCc: Spliceosomal Complex; SER: Smooth Endoplasmic Reticulum; SP: Structural Proteins; SV: Synaptic Vesicle; Ss: Synaptosome; SR: Sarcoplasmic Reticulum; TP: Transport Proteins; ULC: Ubiquitin Ligase Complex
Within the intrauterine development from embryonal days (E1- E22) the neurulation occurs during E7 [1,2]. The CNS develops mainly between E14 and E22 [3]. Neuroplasticity is a fundamental process for brain development and neuro-ontogenesis [4]. It describes the property of synapses, neurons and whole brain regions to adapt their properties depending on their biological task. Different types of neuroplasticity are: evolutionary, reactive, adaptive and reparative plasticity [5]. Synaptic plasticity is the most common form of neuroplasticity during aging and it describes the activity-dependent change of the synaptic transmission strength [6]. In addition, cortical plasticity follows the activitydependent change of the brain size, the connectivity or the activation patterns of cortical networks [7]. Strength and length of stimuli lead to specific interactions of parts of the nervous system which may change the structure of neuronal tissue at the ultrastructural and microscopic level. In terms of differences in the weight of rat brains as well as body mass, changes of the abundance pattern of proteins can be determined. Especially in the first two postnatal months a strong increase in the rats’ brain mass occurs. Between postnatal day P2 and P25, the biggest mass increase can be determined, whereas a further increase happens around P60 [8]. Also until P275 an additional mass increase occurs, but on the whole in a decreased and slower pattern [9].
In the first postnatal weeks a considerable number of migratory processes (e.g. of the glial progenitor cells) are necessary for the development of the CNS. This neuronal migration stops at the time of birth; however, exceptions for different types of cells occur. For example, the migration of neuroblasts in the rostal migratory stream (Figure 1) from the subventricular zone to the olfactory bulb is a physiological process in the adult rat. In addition, neuronal progenitors at the border of the granular cell layer and the hilus show neurogenesis and migrate into the granular layer [10]. The synaptogenesis proceeds in two different phases: early stage (P1-P5) and later stage (P15-P20) [11]. As regards the number of neural and non-neural cells, differences during the development have been observed. While the number of non-neural cells is limited to 4 million cells (ca. 6% of the total cell number) after birth, the number increases up to 140 million cells in the adult rat brain (ca. 50% of the total cell number). A large portion of these non-neural cells originate from the cerebellum (90%). On the whole, the growth of the non-neural cells (except for the cerebellum) is finished by the end of the third postnatal week [8].

Figure 1: Overview of the rat brain at the level of the Cx. In (a-c) three-dimensional reconstructions of the rat brain from dorsal (a), ventral (b) and lateral (c) are shown. Reconstruction has been build using the neuroVIISAS framework (http://neuroviisas.med.uni-rostock.de/neuroviisas.shtml). (d) A sagittal section with circumscribed cerebral cortex. (e) Horizontal section of the rat cerebral cortex of the left and right hemisphere. The black arrow indicates the source at the rostrolateral origin of the rostral migratory stream.
The Cx develops from the prosencephalon (Figure 1) [12]. The development of the Cx is subdivided into different phases. Originating from a three-layer structure, the ventricular, the intermediary and the early marginal zone (preplate) build a new layer (cortical layer), located between preplate and intermediary zone. From here the migration of postmitotic neurons (cortical plate) begins where densely packed and radiating arranged cell extensions are visible. The ventricular zone can be divided into an inner and an outer zone. The inner zone shows a higher cell density than the outer zone [13-16]. An immigration of neurons occurs at the base of the cortical plate [17-18]. Differentiation and synaptogenesis are associative, partly overlapping processes [19]. Within the cortical plate, neuronal cell layering can be divided into the upper marginal zone and the underlying subplate [20]. The cells of the subplate show the characteristics of differentiated neurons and, therefore, can process synaptic information from afferent nerve fibers [21]. Additionally, a projection of the neurons themselves into the cortical plate, the thalamus and the colliculus superior is observed [22].
The Cx shows the largest increase of mass during the first postnatal week. In the following weeks of the first postnatal month, the brain grows progressively, although the increase of its mass decelerates. Between P25 and P60, the mass of the brain decreases again, indicating adolescence [8,23]. The remaining increase of neurons in the Cx is relatively small. The largest increase of neurons in the neocortex and the hippocampus takes place within the first days after birth. This initial cortical process of growth leads to the generation of 1-5 million neurons during the first four days after birth. All in all, an increase of about 24 million neurons in the neocortex and about 4 million neurons in the hippocampus is known. After this surplus of neurons in the neocortex and hippocampus, a substantial decrease of the total cell number (60- 70% of the neurons within the second postnatal week) occurs.
The aim of this study was to determine the differences in protein abundance during the development and aging of the Cx at ages of the brain P7, P90 and aged individuals at P637. Several studies [24-26] focus on the transplantation of immortalized progenitor cells in 6-OHDA Hemiparkinson animal models [24,25] and the differences in the abundance of this area of the brain at different points of development [26]. Hovakimyan et al. [24] and Haas et al. [25] as well as Schwarz and Freed [27] showed that the micro-environment of neonatal brains seems to be important for the differentiation of transplanted neuronal progenitors. Therefore, a further characterization of the abundance patterns of proteins during development in the Cx of the rat brain might provide a better understanding of neuronal development. The determination of candidate proteins which may support the survival and differentiation of transplanted progenitor cells in neonatal and adult caudate-putamen could serve as targets to optimize transplantation. In the past, different studies were performed which concentrated on the properties of the analyzed cells, but these are not associated with the changes in the CNS. For example, factors like the cell-cell interaction, processes in the signal transduction as well as regulatory processes play an important role. Furthermore, such factors and processes seem to be relevant in influencing the differentiation of neuronal stem cells. Here, we searched for differentially expressed proteins which could be of importance for the neuronal differentiation and for the integration of neuronal progenitors in an adult neuronal environment.
Treatment
Male Wistar rats (Rattus norvegicus, Charles River, Sulzfeld, Germany) of different ages (7, 90, 637 postnatal days) with n=6 animals in each age group (2 animals per cage) were used for this study [28,29]. The animals were housed at 22 ± 2° C under an artificial day and night rhythm with a 12 h light-dark cycle with free access to water and standard nutrition. The animal treatment and experimental procedures were conducted in compliance with the regulations and licensing of the local authorities (Landesamt für Landwirtschaft, Lebensmittelsicherheit and Fischerei Mecklenburg Vorpommern, Germany) and the Animal Care and Use Committee of the University of Rostock. Pursuant to the European Communities Council Directive of 24 November 1986 (86/609/EEC) and in accordance with the abovementioned local authorities adequate measures were taken to minimize pain or discomfort.
Perfusion and dissection
At defined dates (7, 90, 637 days postnatal), perfusion was performed. The animals were anesthesized with ether and killed by intraperitoneal Pentobarbital-Na+-injection (60 mg/kg BW). Transcardial perfusion was performed with 100-400 mL (bodyweight depending) cooled (4° C) 0.9% w/v NaCl-solution. After decapitation and brain dissection, the dissected brain regions were weighed and stored at -80° C until homogenization. From pentobarbital injection to -80° C storage it took less than 5 min.
Homogenization
The extraction of proteins was performed according to published standardized protocols [26,30,31]. In the following the steps are described shortly. Proteins of each developmental stage (P7, P90, P637) were separated in six 2D-gels. For P7, P90 and P637, each gel image presents a single dissected brain region. The brain sections were incubated with (9 × probe mass (mg)) μL lysis buffer consisting of 7 M urea (Sigma, Steinheim, Germany), 2 M thiourea (Sigma), 4% w/v CHAPS (Sigma), 70 mM DTT (Sigma), 0.5% v/v Bio-Lyte Ampholytes pH 3-10 (Fluka, Buchs, Switzerland) and a mixture of protease inhibitors (Roche, Basel, Switzerland) additionally enriched with (0.1 × probe mass (mg)) μL Pepstatin A and PMSF (Fluka) and snap-frozen at -150° C. The samples were quickly thawed and transferred into a 2 mL Wheaton potter (neo-lab, Heidelberg, Germany) for homogenization. In the next step, glass beads (0.034 × Vtotal μl) (Roth, Karlsruhe, Germany) were added to the suspension, following a 15 s sonication, 15 s vortexing (repeated six times) and finished by shock freezing the suspension at -150° C. After fast thawing the samples, they were put in a beaker on a magnetic stirrer that was filled with ice water for 15 min. Finally, the samples were centrifuged at 17,860× g for 20 min at 4° C. The supernatant was very carefully removed using a 2 mL syringe (Becton Dickinson, Heidelberg, Germany) with a 0.5 × 25 mm needle (Becton Dickinson), because of a thick lipid coverage derived from myelinated nerve fibers. The protein concentration of the supernatant was determined by the Bradford assay.
Two dimensional polyacrylamide gel electrophoresis (2DE) rehydration
The first dimension was performed in a PROTEAN IEF cell system (Bio-Rad, Berkeley, CA, USA). Protein extracts of 1 mg protein were loaded on immobilized pH 3–10 nonlinear gradient strips with a length of 17 cm (GE-Healthcare, Buckinghamshire, UK; Art.: 17-1235-01) and actively rehydrated with 300 μL rehydration buffer consisting of 6 M urea (Sigma), 2 M thiourea (Sigma), 2% w/v CHAPS (Sigma), 16 mM DTT (Sigma), 0.5% v/v Bio-Lyte Ampholytes pH 3-10 (Fluka) at 50 V for 12 h at 20° C.
First dimension: isoelectric focusing
After rehydration, to reduce artifacts, electrode wicks (Bio-Rad) were added. Focusing started with the “conditioning step” (2 h) which subdivides into two sub-steps: (a) linear voltage rise to 500 V, step-hold 30 min; (b) linear voltage rise to 2500 V, step-hold 1 h. After that, the “slow voltage ramping” (2.5 h): quadratic voltage rise to 8000 V and the “final focusing”: actual process of focusing (duration: 50.000 Vhrs) was performed. During the whole IEF the temperature was constantly kept at 20° C. After focusing the strips were stored at -80° C.
Second dimension: polyacrylamide gel electrophoresis
Focused IPG-strips were equilibrated in two steps of 30 min each in 5 mL of freshly prepared SDS equilibration solution consisting of 1.5 M Tris-HCl pH 8.8 (Roth), 6 M urea (Sigma), 30% v/v glycerol (Sigma), 2% w/v SDS (Sigma), trace of bromophenol blue (Roth) supplemented with 10 mg/mL DTT and 40 mg/mL iodoacetamide. The strips were transferred on 12% v/v homogeneous self-cast sodium dodecyl sulfate polyacrylamide gels (200 × 250 × 1.5 mm). At 125 V per gel (Power Pac 1000, Bio-Rad), they were run in the PROTEAN Plus Dodeca Cell (Bio-Rad). To ensure a constant buffer-temperature of 10° C, a cooling device (Julabo F10, Julabo Labortechnik, Seelbach, Germany) was used.
Fixation and staining
Fixation was performed with acetic acid-methanol solution (45% v/v methanol, 1% v/v acetic acid) overnight. Staining of the gels was performed in a colloidal CBB G250 solution (1 g/1000 mL) (Roth) as described previously [32,33]. After 24 h, the gels were destained with ultrapure water and were held in cold storage (4° C) with ultrapure water until digitization.
Gel analysis
Digitization: The stained gels (n=6) were scanned in transparence mode as 12 bit gray scale tif-images with a F4100 scanner (Heidelberg, Heidelberg, Germany) at 300 dpi resolution. Gels were rinsed in 0.02% w/v sodium azide (Aldrich Chemie, Steinheim, Germany), shrinkwrapped in plastic and stored at 4° C until picking for MALDI-TOFMS (Figure 2).

Figure 2: Overview of the reference gel images from the Cx. The coomassie blue stainings of the gels have similar intensities. (a) The 2DE-gel image of a P7 animal. (b) The 2DE-gel image of a P90 animal is presented. (c) Image of a 2DE-gel of an aged P637 rat.
Digital gel processing: For 2DE-gel image analysis, the software package Progenesis PG200 Version 2006 (Nonlinear Dynamics Ltd., Newcastle upon Tyne, UK) was used. The gels were registered to a reference gel (the gel which contained most spots with optimal separation and staining quality and least artifacts) and manually edited spots (Figure S1) were matched to allow comparability of all gels (Figures 2-3).

Figure 3: Differential abundance of proteins of P7 in comparison to P90. Search of proteins was performed against the Swiss Prot database. Annotation of accession numbers are from Swiss Prot in the reference gel (P90, Co reference gel: 61064). Protein mixes and multiple protein presentations in different spots can be derived from branching lines. Black: upregulated proteins. See also supplemental figures S2-S4 with differential abundance of P90 in comparison to P7, P637 in comparison to P90 and P90 in comparison to P637.
Determination of differentially abundant protein spots
Protein spots in 2DE were quantified by normalizing spot volumes using Progenesis PG200 (Nonlinear Dynamics Ltd., Newcastle upon Tyne, UK) and spot volume differences were calculated. After the comparison of the normalized gray value-spot volumes of all generated spot pairs with Access and Excel (Windows, Microsoft Corporation, Redmond, WA, USA), the comparison of the spot volumes was determined by calculation of the spot volume quotient (SVQ, P90/P7 or P90/P637).
Only those spots were considered to be significantly up- or downregulated that showed a SVQ (Spot Volume Quotient) of 0.6 or less and 1.67 or greater [26,31,34]. The differences were evaluated significantly if differentially expressed spots were detected in at least four gel images (correlated spots) belonging to one test group. If multiple spots of one protein were detected by mass spectrometric analysis, the differential abundance was determined by the mean of their individual abundance levels. Additionally, each protein existing in a mixed spot with at least another protein in one developmental stage was marked individually as well as if one protein was present in multiple spots per stage (Table S1). The classification of the differentially expressed proteins in their respective functional protein groups itself was generated by a comparison of the proteins’ properties; in addition, the basic function of each protein is briefly described.
Bioinformatics analysis
Histological mapping and 3D reconstructions of the rat cerebral cortex were realized in neuroVIISAS (http://neuroviisas.med.unirostock.de). Diagrams were generated with Excel 2013 (Microsoft). 2D-gel image and spot analysis was performed with Progenesis PG200 (shown above). Differentially proteins were subjected to bioinformatics analysis for protein identification (Entrez Gene: https://www.ncbi.nlm.nih.gov/gene/) and UniProtKB/Swiss-Prot (http://www.uniprot.org) as well as UniProtKB/TrEMBL). Classification of differentially proteins was done by using GeneCards (http://www.genecards.org/) and PANTHER (http://www.pantherdb.org). The visualization of intergroup relations was done with a Venn diagram (http://bioinformatics.psb.ugent.be/cgi-bin/liste/Venn/calculate_venn.htpl). The pathway analysis was performed with consensus PathDB (http://cpdb.molgen.mpg.de/), STRING (http://string-db.org/) and Reactome (http://www.reactome.org/).
Mass spectrometric analysis of protein spots
Following published standardized protocols [35], protein spots were excised from the gels with a spot picker (Flexys Proteomics picker, Genomic Solutions, Ann Arbor, MI, USA), transferred into 96-well plates, and subjected to in-gel digestion with trypsin. The gel plugs were washed twice with 30% acetonitrile (ACN) in 25 mM ammonium bicarbonate and 50% ACN in 10 mM ammonium bicarbonate, respectively, shrunk with ACN, and dried at 37° C. The dried gel plugs were re-swollen with 5 μL protease solution (sequencing grade trypsin, 10 ng/μL in 3 mM Tris–HCl, pH 8.5, Promega, Madison, WI, USA) and incubated for 8 h at 37° C. Thereafter, 5 μL of extraction solution (0.3% trifluoroacetic acid, 50% ACN) were added and the samples were agitated at room temperature for 30-60 min before the peptide extracts were transferred into the 96-well collection plates. The resulting peptidecontaining solution was prepared for MALDI analysis by spotting 0.6 μL of the tryptic digest and 0.45 μL of matrix solution consisting of 9 mg/mL α-cyano-4-hydroxy-cinnamic acid (CHCA) in 50% ACN, 0.1% trifluoroacetic acid on standard stainless steel MALDI plates. MALDIMS analysis was performed on a 4700 Proteomics Analyzer MALDITOF/ TOF mass spectrometer (Applied Biosystems, Foster City, CA, USA). All acquired spectra were processed using 4700 Explore™ software (Applied Biosystems, Warrington, Cheshire, UK). For protein identification, spectra were submitted to MASCOT (version 2.4.0, Matrix Science, London, UK) via the MASCOT Deamon. Searches were performed against the subset of rat proteins of the UniProtKB protein sequence database (2012_01; 42755 sequences from Rattus). A mass tolerance of 60 ppm and one missing cleavage site were set, oxidation of methionine residues was considered as variable modification, and carbamidomethylation of cysteines as fixed modification. Peptide masses of trypsin autoproteolysis products and matrix-derived peaks were excluded. Identifications with Mascot scores greater than 59 were considered significant (p<0.05). All results were examined carefully for reliability and occurrence of multiple proteins in the same sample. The search was first restricted to proteins from Rattus and thereafter repeated for rodent proteins to include proteins still missing in the sequence data of Rattus. Entries from TrEMBL were used only, if no homologous protein was contained in Swiss-Prot or if a higher number of peptides matched to a sequence from the TrEMBL database.
Immunoblot analysis
The homogenated protein samples of the Cx were dissolved in sample buffer (1/1 dilution with 2x SB). For immunoblotting, total cellular extracts (30 μg per lane) were separated by SDS-Page using 4-20% polyacryamide gels (Bio-Rad, Munich, Germany) and transferred to PVDF membranes (0.2 mm, Bio-Rad, Munich, Germany). The blots were blocked with 5% non-fat dry milk powder in TBS for 30-60 min and incubated with the individual primary antibodies. The following antibodies were used (dilutions are given in the brackets): rabbit polyclonal anti-PARK 7/DJ-1 (1:800, Abcam, Cambridge, UK), rabbit polyclonal anti Drebrin (1:1000, Abcam, Cambridge, UK), mouse monoclonal anti neurofilament 68 (1:400, Sigma-Aldrich, St. Louis, Missouri, USA), mouse monoclonal anti neurofilament 160/200 (1:1000, Sigma-Aldrich, St. Louis, Missouri, USA), rabbit polyclonal anti α-Internexin (1:1000, Merck Millipore, Darmstadt, Germany). After washing, membranes were incubated with secondary HRP-conjugated anti-mouse (1:2000) and anti-rabbit (1:2000) IgG, and visualized by the Enhanced Chemiluminescence (ECL) (Lumixx plus 250) procedure as described by the manufacturer (Biostep GmbH, Burkhardtsdorf, Germany). For the molecular marker, the Precision plus Protein All Blue Standard (Bio-Rad Laboratories, Inc., Hercules, USA) was used. The analysis of the Western Blot protein signals was performed and recorded by using the Proxima 2850, CL and UV fluorescence/chemiluminescence system (Biostep GmbH, Burkhardtsdorf, Germany) including its analysis software (ProXima AQ-4, Ref. 1.28/CLIQS, version 1.1).
Identification of proteins
As described before, the 2D-PAGE method was performed for the protein separation of the Cx and its different ages (P7, P90, P637) followed by digital gel analysis (“gel-matching”, “spot-warping”) and a differential spot analysis combined with a protein identification (MALDI-TOF) to determine differences in the protein abundance during development and aging.
In Figure 2, the images of the reference gels of the different developmental stages are presented ((a) P7, (b) P90 and (c) P637). The spot compositions proved to be comparable. On average, 740 (±57) spots were detected in the six gels of P7, 729 (±50) spots in the gels of P90 and 488 (±43) spots in the gels of P637. An example of the manual spot editing (segmentation, delineation) by using a gel image of the Cx in Progenesis PG200 is documented in supplemental Figure S1.
In addition, the differential expressed proteins were classified into 13 functional protein groups.
In the next section, an overview of the up- and downregulation of the different protein groups within each developmental stage (P7, P637) is listed and described in relation to the comparison group (P90) for the brain region of the Cx. The specific differential expressed proteins are described in detail in Table 1, in the following text they are only described in a quantitative manner.
| Accession | Entry name | Gene name | Funct. Group/Protein Name | Regulation [P7] | Regulation [P637] | Cell. Localisation |
|---|---|---|---|---|---|---|
| Proteins Carbohydrate Metabolism | (SVQ ≤0.6) | (SVQ ≥1.67) | ||||
| P51635 | AK1A1_RAT | Akr1a1 | Alcohol dehydrogenase [NADP(+)] | up | - | Cts, M |
| P07943 | ALDR_RAT | Akr1b1 | Aldose reductase | down | - | Cp |
| P09117 | ALDOC_RAT | Aldoc | Fructose-bisphosphatealdolase C | - | up | Cp, Mito, A |
| P08461 | ODP2_RAT | Dlat | Dihydrolipoyllysine-residue acetyltransferase component of pyruvate dehydrogenase complex, mitochondrial | down | - | Mmt |
| Q01205 | ODO2_RAT | Dlst | Dihydrolipoyllysine-residue succinyltransferase component of 2- oxoglutarate dehydrogenase complex, mitochondrial | down | down | Mito |
| P04764 | ENOA_RAT | Eno1 | Alpha-enolase | down | up | Cp, M |
| P07323 | ENOG_RAT | Eno2 | Gamma-enolase | down | - | Cp, M |
| P04797 | G3P_RAT | Gapdh | Glyceraldehyde-3-phosphate dehydrogenase | down | - | Cp, Nc |
| P42123 | LDHB_RAT | Ldhb | L-lactate dehydrogenase B chain | down | - | Cp |
| O88989 | MDHC_RAT | Mdh1 | Malate dehydrogenase, cytoplasmic | down | - | Cp |
| B1WC26 | B1WC26_RAT | Nans | N-acetylneuraminic acid synthase | down | down | Cp, Cts |
| P26284 | ODPA_RAT | Pdha1 | Pyruvate dehydrogenase E1 component subunit alpha, somatic form, mitochondrial | down | - | Mmt |
| P49432 | ODPB_RAT | Pdhb | Pyruvate dehydrogenase E1 component subunit beta, mitochondrial | down | - | Mmt |
| P25113 | PGAM1_RAT | Pgam1 | Phosphoglycerate mutase 1 | down | - | Nc, Cts |
| P16617 | PGK1_RAT | Pgk1 | Phosphoglycerate kinase 1 | down | down | Cp |
| P11980 | KPYM_RAT | Pkm2 | Pyruvate kinase isozymes M1/M2 | down | - | Cp, Nc |
| B2RZ24 | B2RZ24_RAT | Pkm3 | Succinyl-CoA synthetase beta-A chain | down | - | Mito, Mmt |
| P48500 | TPIS_RAT | Tpi1 | Triosephosphateisomerase | down | - | Nc, Cts |
| Q68FY0 | QCR1_RAT | Uqcrc1 | Cytochrome b-c1 complex subunit 1, mitochondrial | down | - | Mim |
| Proteins Amino Acid Metabolism | ||||||
| P10860 | DHE3_RAT | Glud1 | Glutamate dehydrogenase 1, mitochondrial | down | up | Mmt |
| P09606 | GLNA_RAT | Glul | Glutamine synthetase | down | up | Cp, Mito |
| F1M9V7 | F1M9V7_RAT | Npepps | Aminopeptidasepuromycin sensitive | - | up | Nc, Cp |
| O09175 | AMPB_RAT | Rnpep | Aminopeptidase B | - | down | Sc, Cp |
| Proteins Fat Metabolism | ||||||
| Q5XI22 | THIC_RAT | Acat2 | Acetyl-CoA acetyltransferase, cytosolic | down | down | Cp |
| Q6P6R2 | DLDH_RAT | Dld | Dihydrolipoyl dehydrogenase, mitochondrial | - | up | Mmt |
| P17425 | HMCS1_RAT | Hmgcs1 | Hydroxymethylglutaryl-CoA synthase, cytoplasmic | up | - | Cp |
| O35760 | IDI1_RAT | Idi1 | Isopentenyl-diphosphate Delta- isomerase 1 | p90: abs | - | Per |
| P30349 | LKHA4_RAT | Lta4h | Leukotriene A-4 hydrolase | down | - | Cp |
| O35264 | PA1B2_RAT | Pafah1b2 | Platelet-activating factor acetylhydrolase IB subunit beta | - | up | Cp |
| Proteins Energy Metabolism | ||||||
| P15999 | ATPA_RAT | Atp5a1 | ATP synthase subunit alpha, mitochondria | down | down | Mim, M |
| P31399 | ATP5H_RAT | Atp5h | ATP synthase subunit d, mitochondrial | down | down | Mito, Mim |
| P25809 | KCRU_RAT | Ckmt1 | Creatine kinase U-type, mitochondrial | down | down | Mito, Mim |
| Q66HF1 | NDUS1_RAT | Ndufs1 | NADH-ubiquinone oxidoreductase 75 kDa subunit, mitochondrial | down | up | Mim |
| Q641Y2 | NDUS2_RAT | Ndufs2 | NADH dehydrogenase [ubiquinone] iron-sulfur protein 2, mitochondrial |
down | - | Mim |
| D3ZG43 | D3ZG43_RAT | Ndufs3 | NADH dehydrogenase (Ubiquinone) Fe-S protein 3 (Predicted), isoform CRA_c | down | - | Mito, Mm |
| P19234 | NDUV2_RAT | Ndufv2 | NADH dehydrogenase [ubiquinone] flavoprotein 2, mitochondrial | - | p637: abs | Mim |
| P20788 | UCRI_RAT | Uqcrfs1 | Cytochrome b-c1 complex subunit Rieske, mitochondrial | down | - | Mim |
| Degratory Proteins | ||||||
| P60901 | PSA6_RAT | Psma6 | Proteasome subunit alpha type-6 | up | up | Cp, Nc |
| Q63347 | PRS7_RAT | Psmc2 | 26S protease regulatory subunit 7 | up | - | Cp, Nc |
| Q63570 | PRS6B_RAT | Psmc4 | 26S protease regulatory subunit 6B | - | p637: abs | Cp, Nc |
| Proteins Antioxidants | ||||||
| Q9Z339 | GSTO1_RAT | Gsto1 | Glutathione S-transferase omega-1 | down | - | Cp, Cts |
| P08009 | GSTM4_RAT | Gstm3 | Glutathione S-transferase Yb-3 | p7: abs | - | Cp |
| P35704 | PRDX2_RAT | Prdx2 | Peroxiredoxin-2 | down | - | Cp |
| O35244 | PRDX6_RAT | Prdx6 | Peroxiredoxin-6 | down | - | Cp, Lyso |
| Q9Z0V6 | PRDX3_RAT | Prdx3 | Thioredoxin-dependent peroxide reductase, mitochondrial | down | up | Mito |
| Proteins Biosynthesis | ||||||
| O35567 | PUR9_RAT | Atic | Bifunctional purine biosynthesis protein PURH | - | up | Mito, Cts |
| Q68FR6 | EF1G_RAT | Eef1g | Elongation factor 1-gamma | up | - | Rs, Cts |
| Q5I0G4 | SYG_RAT | Gars | Glycine--tRNA ligase | up | - | Cp, Mito |
| Q9WTT6 | GUAD_RAT | Gda | Guanine deaminase | up | - | Cts |
| Q6P799 | SYSC_RAT | Sars | Serine--tRNA ligase, cytoplasmic | up | - | Cp |
| P85834 | EFTU_RAT | Tufm | Elongation factor Tu, mitochondrial | down | - | Mito |
| Proteins Signal Transduction | ||||||
| P59215 | GNAO_RAT | Gnao1 | Guanine nucleotide-binding protein G(o) subunit alpha | down | - | M, hGc |
| P54313 | GBB2_RAT | Gnb2 | Guanine nucleotide-binding protein G(I)/G(S)/G(T) subunit beta-2 | down | - | M, Cp |
| P62994 | GRB2_RAT | Grb2 | Growth factor receptor-bound protein 2 | - | up | Nc, Cp |
| P63086 | MK01_RAT | Mapk1 | Mitogen-activated protein kinase 1 | down | - | Nc, Cp |
| F1M7Y3 | F1M7Y3_RAT | Rap1gds1 | RAP1, GTP-GDP dissociation stimulator 1 | up | - | Cp |
| Proteins Regulation | ||||||
| P48037 | ANXA6_RAT | Anxa6 | Annexin A6 | down | - | Cp, Mel |
| P27139 | CAH2_RAT | Ca2 | Carbonic anhydrase 2 | down | - | Cp |
| P45592 | COF1_RAT | Cfl1 | Cofilin-1 | down | - | Nm, Cp |
| Q07266 | DREB_RAT | Dbn1 | Drebrin | up | - | Cp |
| P50399 | GDIB_RAT | Gdi2 | Rab GDP dissociation inhibitor beta | up | - | Cp, M |
| Q68FS4 | AMPL_RAT | Lap3 | Cytosol aminopeptidase | - | up | Cp |
| Q99MZ8 | LASP1_RAT | Lasp1 | LIM and SH3 domain protein 1 | down | down | Cp, Csk |
| Q64361 | LXN_RAT | Lxn | Latexin | down | - | Cp |
| O88767 | PARK7_RAT | Park7 | Protein DJ-1 | down | - | Cp, Nc, Mito |
| P62138 | PP1A_RAT | Ppp1ca | Serine/threonine-protein phosphatase PP1-alpha catalytic subunit | - | up | Cp, Nc |
| Q6AXV4 | SAM50_RAT | Samm50 | Sorting and assembly machinery component 50 homolog | down | down | Mom, Cp |
| P70566 | TMOD2_RAT | Tmod2 | Tropomodulin-2 | down | down | Cp, Csk |
| Q5RKI0 | WDR1_RAT | Wdr1 | WD repeat-containing protein 1 | down | - | Cp, Csk |
| Chaperones | ||||||
| P18418 | CALR_RAT | Calr | Calreticulin | down | down | ER |
| P63036 | DNJA1_RAT | Dnaja1 | DnaJ homolog subfamily A member 1 | p7: abs | - | M, La |
| P63018 | HSP7C_RAT | Hspa8 | Heat shock cognate 71 kDa protein | up | up | Cp, Mel |
| P63039 | CH60_RAT | Hspd1 | 60 kDa heat shock protein, mitochondrial | down | - | Mmt |
| Q5XHZ0 | TRAP1_RAT | Trap1 | Heat shock protein 75 kDa, mitochondrial | down | down | Mito |
| Structural Proteins | ||||||
| P60711 | ACTB_RAT | Actb | Actin, cytoplasmic 1 | down | - | Cp, Csk |
| Q62950 | DPYL1_RAT | Crmp1 | Dihydropyrimidinase-related protein 1 | up | - | Cp, Csk |
| P85845 | FSCN1_RAT | Fscn1 | Fascin | up | up | Cp, Csk |
| P47819 | GFAP_RAT | Gfap | Glial fibrillary acidic protein | down | up | Cp |
| D4A6B2 | D4A6B2_RAT | Immt | Mitochondrial inner membrane protein | down | - | Mito, Mim |
| P23565 | AINX_RAT | Ina | Alpha-internexin | down | - | Nf |
| P19527 | NFL_RAT | Nefl | Neurofilament light polypeptide | down | - | Nf, A |
| P12839 | NFM_RAT | Nefm | Neurofilament medium polypeptide | down | - | Nf, Csk, A |
| B5DFG5 | B5DFG5_RAT | Sep-06 | Septin-6 | down | - | Cr, Cp |
| B0BNF1 | SEPT8_RAT | Sep-08 | Septin-8 | down | down | Cp, Csk |
| Q6AYZ1 | TBA1C_RAT | Tuba1c | Tubulin alpha-1C chain | down | - | Cp, Csk |
| Transport Proteins | ||||||
| P85515 | ACTZ_RAT | Actr1a | Alpha-centractin | up | - | Cp, Cts |
| P02770 | ALBU_RAT | Alb | Serum albumin | up | - | Sc |
| Q5M7T6 | Q5M7T6_RAT | Atp6v0d1 | ATPase, H+ transporting, lysosomal 38kDa, V0 subunit d1 | down | down | M |
| P62815 | VATB2_RAT | Atp6v1b2 | V-type proton ATPase subunit B, brain isoform | down | - | M, Mel |
| P85969 | SNAB_RAT | Napb | Beta-soluble NSF attachment protein | down | down | M |
| Q9QUL6 | NSF_RAT | Nsf | Vesicle-fusing ATPase | down | - | Cp |
| Q9Z2L0 | VDAC1_RAT | Vdac1 | Voltage-dependent anion-selective channel protein 1 | down | down | Mom, M |
| P81155 | VDAC2_RAT | Vdac2 | Voltage-dependent anion-selective channel protein 2 | down | - | Mom |
Table 1: Proteins identified by peptide mass fingerprinting in spots from Coomassie-stained gels arranged by their function. Accession: Accession number from UniProtKB/ Swiss-Prot; Entry name: Entry name from UniProtKB/Swiss-Prot; Gene name: from UniProtKB/Swiss-Prot; Regulation (P7)/Regulation (P637): Differential abundance in comparison to P90.The full description for the cellular location is given in the list of abbreviations. SVQ: Spot volume quotient. Further parameters are shown in Table S1.
Mass spectrometric results of cortical protein identification
At P7, 82 proteins were found which showed a difference in their abundance in comparison to P90. This includes 63 proteins which were downregulated compared to P90, whereas 16 proteins showed an upregulation. In addition, two proteins were analyzed only in P90 and, therefore, were absent at P7. One protein was absent at P90 (Table 2).
| Category | Amount [P7] (Total value) | Amount [P637] (Total value) |
|---|---|---|
| Total amount | 82 | 36 |
| Upregulation | 16 | 17 |
| Downregulation | 63 | 17 |
| Absent | 2 | 2 |
| Absent P90 | 1 | 0 |
Table 2: Overview of the differential abundance of the proteins of the Cx in P7 and P637 in comparison to P90.
Differences in the abundance of proteins of P7 in comparison to P90
The differential abundance of upregulated proteins in P7 (P7>P90) is shown in Figure 3, whereas, upregulated proteins in P90 (P7< P90) rats are documented in supplemental Figure S2.
The largest number of differentially expressed proteins belongs to the carbohydrate metabolism (total amount: 22%) (Figure 4). 17 downregulated (P7< P90) proteins and one upregulated (P7>P90) protein of P7 compared to P90 (Figure 5) were found in this category. For example, the protein alcohol dehydrogenase (Akr1a1) which catalyzes the reduction of a number of aldehydes, including the aldehyde form of glucose, and is thereby implicated in the development of diabetic complications by catalyzing the reduction of glucose to sorbitol which was upregulated at the developmental stage of P7. The protein glyceraldehyde-3-phosphate dehydrogenase (Gapdh) displays a protein which showed a downregulation in comparison to P90. This protein has both glyceraldehyde-3-phosphate dehydrogenase and nitrosylase activities and thereby plays a role in glycolysis and nuclear functions. Therefore, it is a key enzyme in glycolysis that catalyzes the first step of the pathway by converting D-glyceraldehyde 3-phosphate (G3P) into 3-phospho-D-glyceroyl phosphate. Also the protein pyruvate dehydrogenase E1 component subunit beta, mitochondrial (Pdhb) showed a downregulation at this developmental stage. It represents a member of a multienzyme complex that catalyzes the overall conversion of pyruvate to acetyl-CoA and carbon dioxide and provides the primary link between glycolysis and the tricarboxylic acid cycle.

Figure 4: Relative frequencies of differentially expressed cortical proteins of P7 and P90 rats. The differentially expressed proteins were subdivided into 13 groups. Most differential proteins were involved in carbohydrate metabolism. Diagram was generated by using Excel 2013 spreadsheet software (Microsoft).

Figure 5: Number of differential expressed proteins of 13 protein categories of the Cx [P7 vs. P90] (20 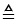 100%). Upregulated and downregulated proteins beside absent proteins are visible. Absent proteins occur in the P7 animals in the groups of chaperones and antioxidant proteins. pcm: carbohydrate metabolism, paam: amino acid metabolism, pfm: fat metabolism, pem: energy metabolism, pd: degradatory proteins, pa: antioxidants, ptm: transmitter metabolism, pb: biosynthesis, pst: signal transduction, pr: regulative proteins, cp: chaperones, sp: structural proteins, tp: transport proteins. Diagram was generated by using Excel 2013 spreadsheet software (Microsoft).
100%). Upregulated and downregulated proteins beside absent proteins are visible. Absent proteins occur in the P7 animals in the groups of chaperones and antioxidant proteins. pcm: carbohydrate metabolism, paam: amino acid metabolism, pfm: fat metabolism, pem: energy metabolism, pd: degradatory proteins, pa: antioxidants, ptm: transmitter metabolism, pb: biosynthesis, pst: signal transduction, pr: regulative proteins, cp: chaperones, sp: structural proteins, tp: transport proteins. Diagram was generated by using Excel 2013 spreadsheet software (Microsoft).
The structural proteins also showed a high amount of differentially expressed proteins. 13% of differentially regulated proteins belong to this class where two upregulated (P7>P90) and nine downregulated (P7< P90) proteins could be identified. For the upregulated proteins at this developmental stage, for example, the dihydropyrimidinaserelated protein 1 (Crmp1) showed a higher abundance at the early developmental stage of P7 compared to P90. The protein fascin (Fscn1) was also upregulated at this stage, which organizes filamentous actin into bundles and the formation of microspikes, membrane ruffles, and stress fibers. Therefore, it is important for the formation of a diverse set of cell protrusions, such as filopodia, and for cell motility and migration. Different members of the neurofilaments were downregulated in P7, for example, the protein alpha-internexin (Ina) as well as the neurofilament light polypeptide (Nefl) and the neurofilament medium polypeptide (Nefm) which showed a higher abundance at P90 compared to P7. These different members of the neurofilaments belong to the type IV intermediate filament heteropolymers which are composed of light, medium, and heavy chains. In general, neurofilaments comprise the axoskeleton and they functionally maintain the neuronal caliber. They may also play a role in intracellular transport to axons and dendrites and their morphogenesis. Additionally, for the neurofilaments (Nefl, Nefm) this trend of abundance could be verified by Western Blot analysis (Figure 9). Also the protein actin, cytoplasmic 1 (Actb) showed a lower abundance at P7 in comparison to P90. This protein represents a member of one of the six different actin proteins. Actin members are highly conserved proteins that are involved in cell motility, structure, and integrity.
Another large differential abundance was found for the regulatory proteins (total amount: 13%). They showed a stronger tendency to downregulation (P7< P90) (nine downregulated proteins, two upregulated proteins). In this category, for example the downregulated protein annexin A6 (Anxa6) represents a member belonging to a family of calcium-dependent membrane and phospholipid-binding proteins which have been implicated in membrane-related events along exocytotic and endocytotic pathways.
Another downregulated protein in comparison to P90, Protein DJ-1 (Park7), functions as a redox-sensitive chaperone and as a sensor for oxidative stress. It can protect neurons against oxidative stress and cell death. In P7 the Rab GDP dissociation inhibitor beta (Gdi2) as well as the protein Drebrin (Dbn1) showed a higher abundance (P7>P90). While Gdi2 presents a member of GDP dissociation inhibitors which regulates the GDP-GTP exchange reaction of members of the rab family and is therefore involved in vesicular trafficking of molecules between cellular organelles, Dbn1 functions as a cytoplasmic actinbinding protein that plays a role in the process of neuronal growth. Also by Western Blot analysis the upregulation of Dbn1 at P7 in comparison to P90 could be confirmed (Figure 9).
In the remaining functional protein classes, i.e. the transport proteins (10% total amount, two upregulated (P7>P90) and six downregulated (P7< P90) proteins), the proteins of the energy metabolism (Total amount: 9%, seven downregulated (P7< P90) proteins), the chaperones (Total amount: 6%, three downregulated (P7< P90) proteins and one upregulated (P7 >P90) protein, one protein was absent at P7), proteins of the biosynthesis (Total amount: 6%, four upregulated (P7>P90) and one downregulated (P7< P90) protein), antioxidant proteins (total amount: 6%, four downregulated (P7< P90) proteins, one protein absent at P7), proteins of the fat metabolism (total amount 5%, two downregulated (P7< P90) proteins, one upregulated (P7 >P90) protein, one protein is absent at P90), proteins involved in the signal transduction (total amount: 5%, one upregulated (P7>P90) and three downregulated (P7< P90) proteins), proteins of the amino acid metabolism (total amount: 2%, two downregulated (P7< P90) proteins a well as the degratory proteins (total amount: 2%, two upregulated (P7>P90) proteins), differences in the abundance of the protein categories could be determined as well. Only proteins involved in the metabolism of neurotransmitters showed no detectable differences in these ages of the brain during the analysis.
Differences in the abundance of proteins of P637 in comparison to P90
Gels documenting the differential up- and downregulation of proteins with annotated accession numbers of the time points P90 and P637 are shown in supplemental Figures S3 and S4.
Several differentially expressed proteins could be found at P637 and P90 of development. In principle, a smaller number of differentially expressed proteins were detectable as compared to P7. Taken together, a change in their abundance was observable for 36 proteins between both developmental stages. 17 proteins showed an upregulation (P637>P90), 17 proteins were downregulated (P637< P90) and two proteins were only detectable at P90, respectively they were absent at P637 (Table 2). The proteins of the energy metabolism show the most differentially expressed proteins (total amount: 14%) (Figure 6). Three proteins showed a downregulation (P637< P90), whereas one protein was upregulated (P637>P90) and one protein was absent at P637 (Figure 7). As for the two subunits of the mitochondrial ATP synthase (subunit alpha (Atp5a1), subunit d (Atp5h)) showed a lower abundance (P637< P90). Both subunits participate in the catalysis of ATP synthesis by using an electrochemical gradient of protons across the inner membrane during oxidative phosphorylation. The NADH-ubiquinone oxidoreductase 75 kDa subunit (Ndufs1) was upregulated which has NADH-dehydrogenase activity and oxidoreductase activity and transfers electrons from NADH to the respiratory chain. NADH dehydrogenase flavoprotein 2 (Ndufv2) was analyzed as absent at P637 in comparison to P90 which also functions as a subunit of the mitochondrial membrane respiratory chain NADH dehydrogenase (Complex I) that is believed to belong to the minimal assembly required for catalysis. Other functional protein classes possessing similar amounts of differentially expressed proteins (14%) were determined. Three regulatory proteins were downregulated (P637< P90) and two proteins were upregulated (P637>P90). The downregulated tropomodulin-2 (Tmod2) presents a neuronal-specific member of the tropomodulin family of actin-regulatory proteins. The protein caps the pointed end of actin filaments preventing both elongation and depolymerization, whereas the capping activity of this protein is dependent on its association with tropomyosin. The serine/ threonine-protein phosphatase PP1-alpha catalytic subunit (Ppp1ca) was upregulated (P637>P90) and is needed for cell protection against oxidative stress and cell death acting as an oxidative stress sensor and redox-sensitive chaperone. This protease is also required for the correct mitochondrial morphology and its function as well as for the autophagy of dysfunctional mitochondria.

Figure 6: Relative frequencies of differentially expressed cortical proteins of P637 and P90 rats. The differentially expressed proteins were subdivided into the same 13 groups as done above. Most differential proteins were involved in carbohydrate and energy metabolism. Structural and transport proteins decreased, however, proteins of the amino acid metabolism increased by comparing with P7 and P90. Diagram was generated by using Excel 2013 spreadsheet software (Microsoft).

Figure 7: Number of differential expressed cortical proteins of the 13 protein categories of P637 and P90 rats (20 100%). Absent proteins occur in the energy ight="23"> 100%). Absent proteins occur in the energy metabolism and degradatory groups of proteins. Absent proteins occur in the P7 animals in the groups of chaperones and antioxidant proteins. Diagram was generated by using Excel 2013 spreadsheet software (Microsoft).
The proteins of the carbohydrate metabolism (three downregulated proteins and two upregulated proteins) showed the same amount (14%) of differentially expressed proteins at this developmental stage. Phosphoglycerate kinase 1 (Pgk1) showed a downregulation (P637< P90). It catalyzes the conversion of 1.3-diphosphoglycerate to 3-phosphoglycerate within the glucose metabolism. Fructose bisphosphate aldolase C (Aldoc) was upregulated (P637>P90). It shows a relatively strong abundance in the hippocampus and in cerebellar Purkinje cells. It represents a glycolytic enzyme that catalyzes the reversible aldol cleavage of fructose-1.6-biphosphate and fructose 1-phosphate to dihydroxyacetone phosphate and either glyceraldehyde- 3-phosphate or glyceraldehyde. Alpha-enolase (Eno1) showed a stronger abundance signal (P637>P90). It is a multifunctional enzyme that takes part in glycolysis, growth control, hypoxia tolerance and allergic responses.
Within the amino acid metabolism (total amount 11%), three upregulated (P637>P90) and one downregulated (P637< P90) proteins were found. Furthermore, the proteins of the fat metabolism (total amount: 8%) showed two upregulated (P637>P90) and one downregulated (P637< P90) protein. The same amount of differentially expressed proteins could also be determined within the group of transport proteins (three downregulated (P637< P90) proteins), the structural proteins (one upregulated (P637>P90) and two downregulated (P637< P90) proteins) as well for the chaperones (one upregulated (P637>P90) and two downregulated (P637< P90) proteins). The proteins which are involved in degratory processes (total amount: 6%, one upregulated (P637>P90) protein, one protein was absent in P637), proteins of the signal transduction (total amount: 3%, one upregulated (P637>P90) protein) as well as the proteins with antioxidant properties (Total amount: 3%, one upregulated (P637>P90) protein) and the proteins of the biosynthesis (total amount: 3%, one upregulated (P637>P90) protein) also showed a differential abundance, but to a lesser extent.
In the set or Venn diagram (Figure 8) differentially expressed proteins were identified that share different experimental groups. The three upregulated proteins Psma6, Hspa8 and Fscn1 are upregulated in the P637 and P7 group in relation to P90. In both groups six differentially expressed proteins that are downregulated in P7, however, upregulated in P637 with regard to P90 could be identified: Ndufs1, Prdx3, Glud1, Eno1, Gfap, Glul. 16 differentially expressed proteins that are downregulated in the P7 as well as in the P637 group with regard to the P90 rats were found: Acta2, Atp6v0d1, Samm50, Lasp1, Nans, Atp5h, Calr, Sep08, Atp5a1, Napb, Dlst, Pgk1, Ckmt1, Trap1, Tmod2, Vdac1.

Figure 8: This Venn diagram shows the logical relations between the differentially proteins of all experimental groups. The number proteins that show differences in two groups are indicated by overlapping regions. up7: number of upregulated proteins in the P7 group in relation to the P90 group. down7: number of downregulated proteins in the P7 group in relation to the P90 group. up637: number of upregulated proteins in the P637 group in relation to the P90 group. down637: number of downregulated proteins in the P637 group in relation to the P90 group.
Western blot validation
Five exemplary proteins with differential abundances in ages of the brain P7, P90 and P637 were validated using Western Blot analysis, since this method is one possibility to verify the proteome.
The structural proteins α-Internexin (Ina), the neurofilaments (neurofilament medium polypeptide (NF-M, Nefm), neurofilament light polypeptide (NF-L, Nefl)) and the regulatory protein DJ-1 (Park7) were downregulated (P637< P90) in the Western Blots (Figure 9). The precise values for the abundance changes (band volumes) can be found in Supplemental Table S2.

Figure 9: Validation of single differentially expressed proteins of the Cx with Western Blot analysis (rabbit polyclonal anti Drebrin, rabbit polyclonal anti-PARK 7/DJ-1, rabbit polyclonal anti α-Internexin, mouse monoclonal anti neurofilament 68, mouse monoclonal anti neurofilament 160/200, between the developmental stages (P7, P90 and P637). Differential abundances of proteome analysis are shown in brackets.
Drebrin (Dbn1) was upregulated at P7 (P7>P90) which was confirmed by the Western Blot. On the basis of Figure 9, especially the band between 75 kDa and 150 kDa (red circle, Figure 9) showed a stronger signal compared to the lanes of P90 and P637, whereas the Dbn1 antibody used, typically detects a signal with a molecular weight of 65 kDa for this protein. A possible explanation for the additionally detected bands could be a result due to the large number of negatively charged residues in this protein.
The postnatal development and aging of the cerebral cortex (Cx) of the laboratory rat (Figure 1) can be quantified on the level of mRNA, immunohistochemical detection of proteins, histochemical and chemoarchitectonis as well as macroscopic small animal MRI studies. Here, we investigated the development and aging of the cortical proteome by gel electrophoresis and analytical mass spectrometry to reveal differentially expressed proteins during postnatal P7 (juvenile), P90 (adult) and P637 (aged) laboratory rats. An insight into the development of the postnatal development of the cortical connectome may offer new targets for genetherapy and transplantation of progenitor cells for neurodegenerative diseases.
The proteome of the cerebral cortex (Cx) of the laboratory rat (Figure 1) was analyzed by mass spectrometry at different dates of development (postnatal P7 (juvenile), P90 (adult), P637 (aged)) to determine those proteins that are differentially expressed during significant stages of postnatal development and aging. Proteins which are involved in processes of proliferation, migration and differentiation were of particular interest. The analysis should contribute to a better understanding and characterization of the proteome development of whole tissue preparation including the microenvironment. The knowledge of differentially expressed proteins could support the successful transplantation of progenitor cells in models of neurodegenerative diseases.
The Cx does not consist of a homogeneous cell population (neurons, glia and other cell population, extracellular matrix, neuropil). Protein abundance of cells and ECM of different cortical regions overlap in the gels. Therefore, a proteomic analysis of this large part of the brain also displays protein changes of non-neural cell populations and neuropil besides the differentially expressed proteins of the neural cells. In addition, using the method applied in this study also involves certain limitations, for example, a protein separation within the range of approximately 10-100 kDa and between a pH-range of 3-10. Hence, it is possibe to analyze a high percentage of the proteome of the Cx, but not its full composition. As shown in Figure 2, it is observable that especially the lateral parts of the gels show a lower resolution than the central parts which can lead to a possible misinterpretation of the regulation analysis of some protein spots.
Therefore, it is possible to analyze a major part, but not the entire proteome of the Cx [36]. Additionally, it has to be mentioned that no analysis of enzyme activity was performed in this study. The enzymatic activity of the specific proteins which can be influenced by factors like the substrate availability or the surrounding pH-value, for example, are beyond the scope of this investigation [26].
Furthermore, it is known that some proteins may be compromised in multiple spots within different locations in gels which can be a reason for different posttranslational modifications and isoforms of the same protein. As a result, a particular spot of a single protein may only show an average change of the spot volume but this does not concurrently apply for a specific protein of such a mixture of proteins within the same spot. However, with additional mass spectrometric detection methods like the stable isotope ratio mass spectrometry (SIRMS) it would be possible to identify both the protein and its variants more precisely and to overcome the sample-to-sample recovery variabilities associated with non-SIRMS MS-proteomic methods. Also an improved analysis of the lower expressed and abundant proteins would be allowed by using affinity-enrichment-MS methods and targeted biomarker discovery applications (e.g. IDBEST™, iTRAQ™) [37]. The UniProtKB protein sequence database in the version of 2012_01 with 42755 sequences from Rattus has been used. An update of the search could reveal further aspects of identified proteins. However, the search was first restricted to proteins from Rattus and thereafter repeated for rodent proteins to include proteins still missing in the sequence data of Rattus. Hence, entries from TrEMBL were used only, if no homologous protein was contained in Swiss-Prot or if a higher number of peptides matched to a sequence from the TrEMBL database.
Therefore, two databases should provide a reliable identification of the proteins that were found in this study. The results of this study which deals with the proteomic analysis of the Cx of the rat can be seen as a first step to obtain an overview of the differentially expressed proteins in this multiarea part of the laboratory rat prosencephalon.
Proteins exhibit changes of abundance in independent samples at significant developmental stages of the Cx. The different abundance levels of the proteins were determined and the changes were compared with findings in the literature. In general, specific changes in the abundance of cytoplasmatic proteins in the postnatal development of the Cx were identified.
Abundance changes of proteins of the carbohydrate metabolism of P7 and P90 are relatively strong. At P637, mainly proteins of the energy metabolism, regulatory proteins and proteins participating in carbohydratic processes demonstrated the highest abudance of differentially expressed proteins. These findings suggest that the above mentioned categories of proteins are important for the development of the rat brain, for example, for the maintenance of the cytoskeleton and the energy metabolism of the rat Cx.
Most of the proteins of the carbohydrate metabolism (total amount of differentially expressed proteins: P7 vs. P90: 22%; P637 vs. P90: 14%) at the developmental stage P7 are involved in the tricarboxylic acid cycle and glycolysis. Almost all of these differentially expressed proteins show a downregulation at the early stage of P7 compared to P90. For example, the proteins gamma- and alpha enolase (Eno1, Eno2) are downregulated towards P90. While Eno1 presents a multifunctional enzyme that, besides having its functional role in glycolysis, plays a part in various processes such as growth control, for example, Eno2 catalyzes the conversion of 2-phosphoglycerate to phosphorenolpyruvate and which also has neuroprotective properties [38]. Already Wilbur and Patel [39] described the activities of several proteins involved in the tricarboxylic acid cycle and pyruvate metabolism showing low abundance levels in newborn rats, whereas they are increased markedly to adult levels during 10-30 postnatal days but do not decrease in adulthood. Also other members of this group like phosphoglycerate kinase 1 (Pgk1) and its isoenzymes (pyruvate kinase isozymes M1/M2 (Pkm2)) participate in glycolysis and show a downregulation (P7< P90). The sole upregulated protein in this category (alcohol dehydrogenase (AKR1A1)) is a member of proteins which catalyze the reduction of mevaldate to mevalonic acid and of glyceraldehyde to glycerol and are also involved in a variety of aromatic and aliphatic aldehydes to their corresponding alcohols. Further, differentially regulated proteins which are directly involved in various steps of the carbohydrate metabolism (e.g. aldose reductase (Akr1b1), pyruvate dehydrogenase E1 component subunit alpha (Pdha1), and pyruvate dehydrogenase E1 component subunit beta (Pdhb)) could be identified. Summarizing, carbohydrate proteins show a tendency to a lower abundance at P7 in comparison to P90. This juvenile (P7) carbohydrate downregulation could be the outcome of a postnatal recreation. The placenta supplements of the rat’s mother seem to deliver sufficient supplies for the carbohydrate metabolism of the juvenile rat brain. After birth, maternal feeding also provides enough glucose, lactate and ketone bodies for the juvenile metabolism. Within P5 the rat exhibits the highest ketone body concentration which subsequently decreases up to P30 [40]. By the change of the energy substrate in the adult animal, a higher abundance of proteins which are important for the metabolism of glucose is known [41]. Under normoxic conditions, the scope of glucose uptake accounts for one fifth of the amount in the adult animal [42]. Also Wilbur and Patel [39] were able to demonstrate that different proteins involved in the tricarboxylic acid cycle and the pyruvat metabolism show a lower abundance level in newborn rats. Whereas these abundance levels increase between postnatal days P10-P30, a reduction of the abundance is not detectable. Partly, for some of the proteins of the glucose metabolism an upregulation was also detectable between P90 and P637. An explanation for this could be the reduction of the metabolisation of glucose at the higher age which can lead to an impairment of performing cognitive tasks. An upregulation of these proteins can present an indication for the reaction of the animal to maintain physiologic basic states and, therefore, to produce enough energy for the neural integrity of the animal.
Major differences in the differential abundance of structural proteins between P7 and P90 were found (total amount of differentially expressed proteins: P7 vs. P90: 13%; P637 vs. P90: 8%). Most structural proteins are cytoskeletal components which are indispensable for the highly complex morphologies of dendritic and axonal arbors, which are developing in this period. This concerns the dynamic organization of the cytoskeleton [43], the axonal and the dentritic expansion as well as the establishment of synaptic contacts [44-46]. The gene of beta-Actin (Actb) (P7< P90) encodes one of six different actin proteins. Actins are highly conserved proteins that are involved in cell motility, structure, and integrity. The determined differential abundance could be explained by the associated dynamic intracellular mechanism of this protein. The protein can be found in two different forms in the cell: G-actin describes the globular monometric form, whereas F-actin forms helical polymers. Both forms present flexible structures which have important roles inside dynamic, regulated actin-networks [47]. The protein has the capability of four main functions in this network: the composition of microfilaments by forming polar rails for kinesin motorproteins (for the vesicle and organelle transport), as a component for the cytoskeleton, influencing the cell motility by (de-) polymerisation of fibrils, as well as the connection of myosin for the generation of the actomyosin complex.
Members of the septin protein family show a differential regulation within the determined developmental stages. These proteins present GTP-binding macromolecules which are involved in the control and regulation of cellular processes to synthesize filaments. As described in other sources, because of their filamentous organization as well as their association with cell membranes, actin filaments and microtubles these proteins are referred to as cytoskeletal components. In general, septin filaments show no polar formation, but their composition is comparable to the formation of intermediary filaments [48].
The protein septin-6 (Sept6) shows a downregulation at the developmental stage P7 compared to P90. Septin-8 (Sept8) is downregulated at P7 and also exhibits a downregulation at P637 compared to P90. The findings suggest a high occurrence of the expression of these proteins at the developmental stage P90. As stated before, because of the differential regulation of the different members of this protein family combined with their dynamic properties, e.g., the impact of dendritogenesis and axogenesis [49], this may give an indication for their differential abundance.
Furthermore, different members of the neurofilament family with a differential abundance between the investigated developmental stages could be found. The proteins neurofilament light polypeptide (Nefl) as well as neurofilament medium polypeptide (Nefm) show a downregulation (P7< P90). This observation was confirmed by Western Blot analysis (Western Blot validation (Figure 9)). The neurofilaments belong to the type IV intermediate filaments which are classified in light, medium and heavy chain proteins. Functionally, these proteins play a major role in axonal growth, neuronal polarity and axonal signal transduction [50]. As described by Riederer et al. [51], differences in the regulation can occur during the ontogenesis, whereas a distinction has to be made between different cell types and their abundance. In the cerebellar cortex of cats afferent mossy fibres and climbing fibres express Nefl and Nefm at the time of birth. However, in other cell types a different abundance of these proteins was observed. For example, the axons of basket cells express these proteins one month postnatal. Relatively late, Nefl and Nefm are expressed in parallel fibres, detectable at the beginning of the third month after birth. Therefore, this study can provide an idea why the proteins show a higher differential abundance after P7, whereas no differentiation is found between the different compartments.
The protein dihydropyrimidinase-related protein 1 (Crmp1) as well as the protein fascin (Fscn1) are upregulated (P7>P90). While Fscn1 belongs to the familiy of actin-binding proteins and organizes F-actin into parallel bundles, it also plays a critical role in cell migration, motility, adhesion and cellular interactions, dihydropyrimidinase-related protein 1 (Crmp1) belongs to the dihydropyrimidinase-likeproteins family which plays an important role in the development and maturation of neurons as well as the axonal growth [52]. The determined upregulation of this protein at the early stage of P7 can be explained by an indication of an increased formation of neuronal components which are required for the synaptic signal transduction for example.
Proteins of the energy metabolism also show a major percentage of differentially regulated proteins within the Cx (total amount of differentially expressed proteins: P7 vs. P90: 9%; P637 vs. P90: 14%). An upregulation of cortical proteins of the energy metabolism was found [53]. With development, the oxidative energy metabolism of the cortex decreases and the adaption to oxidative stress conditions is reduced [54]. Land et al. [55] and Hamada-Kanazawa [56] show that mitochondria increase in the brain by the factor of 6 between P7 and P25. In mitochondria of older animals, a smaller number of respiratory subunits can be found leading to a decrease of energy production [57].
The mitochondria are indirectly involved in the development of neuronal networks and neuronal pasticity (neuronal differentiation, neurite outgrowth, neurotransmitter release as well as the dentritic alteration) by purposing energy components (ATP and NAD+) as well as the regulation of subcellular Ca2+ ions and stabilizing the redox-homeostasis [58]. A certain number of proteins of the energy metabolism contribute to these components, subunits of the respiratory chain and the ATP-synthesis. They will be outlined in the following.
Mitochondrial ATP synthase subunit alpha (Atp5a1) is downregulated at developmental stages P7 and P637 compared to P90, whereas the mitochondrial ATP synthase subunit d (Atp5h) shows the same abundance (downregulated at P7 and P637 compared to P90). The protein ATP synthase is a key-protein required for the production of energy. The differential regulation of some of its subunits within the brain region examined indicates an increased abundance of these proteins after P7 (shown in previous paragraph). The differential regulation of these proteins in the aged rats could be caused by a reaction of the animal to counteract the reduction of the energy production. In addition, several members of the respiratory chain (NADH dehydrogenase [ubiquinone] iron-sulfur protein 2, mitochondrial (Ndufs2), NADH-ubiquinone oxidoreductase 75 kDa subunit, mitochondrial (Ndufs1), NADH dehydrogenase (Ubiquinone) Fe-S protein 3, isoform CRA_c (Ndufs3), NADH dehydrogenase [ubiquinone] flavoprotein 2, mitochondrial (Ndufv2)) are differentially expressed during different stages of development. While Ndufs1, Ndufs2 and Ndufs3 are downregulated at P7 and exhibit no abundance changes around P637 (besides the protein Ndufs1 which is upregulated at P637), Ndufv2 has the identical abundance level at P7 and P90, whereas this protein is found to be absent at P637. In general, the protein NADH dehydrogenase plays an important role in mitochondria. It forms the complex 1 of the respiration chain and links the catalyzation of the oxidation of NADH with the reduction of coenzyme Q. The reduced abundance of NADH dehydrogenase (P7< P90) is probably associated with the ongoing neurogenesis [59], the synaptogenesis and maturation of the neurons coupled with an increased energy consumption [60] after P7 during development. The protein cytochrome b-c1 complex subunit Rieske (Uqcrfs1) shows a downregulation at developmental stage P7 (P7< P90). This protein presents a component of the ubiquinol cytochrome c reductase complex which itself is known to be involved in the production of an electrochemical gradient as well as the resulting synthesis of ATP within the mitochondria. The downregulated subunit of the cytochrome c reductase complex exposes a comparable regulation with the components of the NADH dehyrogenase as was discussed before. The location of these subunits in the mitochondria coincides with similar differential abundances. The differential abundance of this protein may indicate a possible role in the maturation of the Cx especially during neurogenesis.
Regulatory proteins show an obvious differential abundance (total amount of differentially expressed proteins: P7 vs. P90: 13%; P637 vs. P90: 14%). The chaperone like protein DJ-1 (Park7) is downregulated at P7 (P7< P90) and can protect neurons against oxidative stress and cell death. In Western Blot analysis, the downregulation of this protein has been confirmed (Figure 9). As demonstrated by Mitsumoto and Nakagawa [61], DJ-1 may act as an endogenous indicator of oxidative stress and may exhibit an anti-apoptotic effect. As stated by Im et al. [62], Mo et al. [63] and Klawitter et al. [64], this protein also has the ability to inhibit oxidative stress inducing the following apoptosis. Moreover, it has been reported that this protein can prevent apoptotic cell death by recruiting and activating an additional protein (anti-apoptotic protein kinase, Akt) which functions as a serine/threonine-protein kinase and regulates many processes including metabolism, proliferation, cell survival, growth and angiogenesis. A higher abundance of this protein at the adult stage of P90 compared to juvenile rats (P7) could indicate a reaction of the animal to increasing oxidative stress during aging, thus acting as one of the early signs to prevent this factor.
Tropomodulin-2 (Tmod2), the neuronal form of the protein tropomodulin is a regulatory protein downregulated at the stage of P7 (P7< P90). The protein stabilizes the actin filaments and regulates the length of the filaments [65,66]. The regulatory protein cofilin-1 (Cfl1) is also involved in the organization of actin and influences the morphology of cells. The protein is downregulated at P7 in the Cx. In previous studies, an upregulation of this protein could be determined at P7 in other regions of the brain (olfactory bulb, cerebellum) [31,67]. Also Gurniak et al. [68] show that a high abundance of this protein is essential at the early levels of development. For example, the closure of the neural crest combined with the migration of cell populations is strongly reduced in the absence of this protein. The protein drebrin (Dbn1) shows an upregulation at P7 (P7>P90). This protein is neuronspecific and located in synapses to support neuronal growth. The upregulation of Dbn1 was confirmed in Western Blot analysis at P7 (Figure 9). It was shown that a higher abundance of this protein is present especially between E19 and the first postnatal week. A decreased abundance of Dbn1 seems to be associated with loss of memory in Alzheimer's disease [69].
The protein family of the annexins can bind lipids (mostly acid phospholipids) of the cell membrane with one or several negative charges to oligomerise to trimers or hexamers. If a hexameric formation is established, the requirement for building of transport proteins for ion channels is given. The functional group of the annexins is besides the group of calmodulines necessary for binding calcium [70]. Annexin A6 (Anxa6) shows a downregulation (P7< P90). It is known that the development of the brain and maturation of the rat itself is excessive around P10 and the synaptogenesis sets in during the third and fourth postnatal week [71-73]. Furthermore, Giambanco et al. [74] show that Anxa5 and the accumulation of this protein during the first postnatal week indicates a coincidence of differential regulation and brain development.
The interactions of differential proteins were determined by an induced network module analysis. The consensusPathDB (http:// cpdb.molgen.mpg.de/), PANTHER, Reactome and STRING were used to assemble networks of up- and downregulated proteins. The consensusPathDB turned out to generate interactions of differential proteins in one connected component networks. Most of the structural proteins which are differential are downregulated in P7 animals (P7< P90). However, fascin (Fsnc1), drebrin (Dbn1) and Rab GDP dissociation inhibitor beta (Gdi2) are upregulated. Fascin is necessary to form actin bundles of filamentous actin. This is mandatory for mobility and migration. Especially in the early phase of postnatal development these processes seems to be correlated with fascin upregulation found here. Like fascin upregulation is an important factor for neuronal growth. Therefore, drebrin upregulation appears to be associated with the fascin upregulation since both proteins concordantly support processes of neuronal development. In addition, Gdi2 is important for vesicular trafficking that is located within the plasma membrane where neuronal motility as well as neuronal growth are linked. These three upregulated and functionally concurrent proteins have multiple interaction partners in the pathway centric protein analysis found with the consensusPathDB. They can interact on each other upon one intermediate protein, only (Figure 10). It was found that all upregulated P7 proteins can build one connected network component. Hence, these consensual upregulated proteins may interact directly or indirectly to develop structural components of cortical cell populations.

Figure 10: Interactions of upregulated and downregulated proteins of the P7 group. P7 proteins were compared with P90 proteins. Black names: differentially expressed proteins, magenta names: intermediate nodes, orange lines: protein interactions, blue lines: gene regulatory interaction, green lines: biochemical interaction, blue rectangle: gene label, gray rectangle: family unknown.
In the P637 group a downregulation of three different ATPases was found: Atp5a1, Atp5h, and Atp6v0d1. After generating the network of protein interactions a massively biochemically interlinked (green dashed lines) subnetwork that is connected with the three differentially expresses ATPases was detected (Figure 11). Downregulation of these 3 ATPases may directly affect such a ATPase-subnetwork in aged rats because most of the other ATPases (magenta names in Figure 11) are direct interaction partners of the downregulated Atp5a1, Atp5h and Atp6v0d1.

Figure 11: Interactions of upregulated and downregulated proteins of the P637 group. P637 proteins were compared with P90 proteins. The massive biochemical interactions of different ATPase’s are obvious. 3 of them (in black) were downregulated in P637 rats. Black names: differentially expressed proteins, magenta names:intermediate nodes, orange lines: protein interactions, blue lines: gene regulatory interaction, green lines: biochemical interaction, blue rectangle: gene label, orange rectangle: RNA label.
In summary, most cytoskeleton proteins (Neurofilaments, β-actin) are downregulated in P7 (P7< P90). Some proteins of migration and motility as well as proteins for maturation and development are upregulated in P7 animals (P7>P90). Additionally, Drebrin and Gdi2 which are important for vesicular trafficking and neuronal growth are upregulated in P7 (P7>P90). In aged animals oxidative stress sensors, proteins necessary for autophagy of dysfunctional mitochondria, growth control and hypoxia tolerance (Ppp1ca, Eno1) turned out to be upregulated (P637>P90). Overall, energy consumption and differentiation processes as well as specific regulatory mechanisms can be observed at least indirectly by differential abundances of proteins during ageing.
Michael Wille carried out the spot segmentation and drafted the manuscript. Antje Schümann carried out the two-dimensional polyacrylamide gel electrophoresis. Grit Mutzbauer optimized two-dimensional polyacrylamide gel electrophoresis for brain tissue. Michael Kreutzer participated in the database management of the spot lists and performed the picking of spots. Michael O. Glocker, Andreas Wree and Oliver Schmitt participated in the design of the study. Oliver Schmitt performed the dissection of the material and the sample preparation. Oliver Schmitt conceived of the study, and participated in its design and coordination and helped to draft the manuscript. All authors read and approved the final manuscript.
We appreciate the outstanding work of Stefan Mikkat by identifying the proteins with the technique of MALDI-TOF-MS.