Journal of Oceanography and Marine Research
Open Access
ISSN: 2572-3103
ISSN: 2572-3103
Research Article - (2013) Volume 1, Issue 3
The need for discovery of new antibiotics is imperative because previous evidence suggest that development of resistance to any new antimicrobial agents is inevitable. In addition, compounds not tested for antimicrobial activity also gives ample scope for rediscovering compounds with antimicrobial activity. This study screens extracts (Diethyl Ether and Butanol) of a marine red-Sea sponge Negombata magnifica for in vitro fungicidal activity against 10 plant and 3 fish pathogens. Fungicidal activity of the crude diethylether fraction of the sponge against test strains showed exceptionally high activity against three virulent fish pathogens Exophiala salmonis, Branchiomyces demigrans and Saprolegnia sp. Furthermore, Minimal Inhibitory Concentration (MIC) of the compound was evaluated with respect to Exophiala salmonis (1.5 μg/ml) Branchiomyces demigrans (0.75 μg/ml) and Saprolegnia sp. (3 μg/ml) respectively. Finally, bioassay guided isolation and purification led to the identification of the active compound as latrunculin B on the basis of spectroscopic data. This finding suggests that latrunculin B, could be considered as a lead molecule for the design of new antifungals for managing common fungal diseases in aquaculture. Toxicity testing of this compound on shrimps indicated that the compound was non toxic up to a concentration as high as 5 mg/ml suggesting its suitability for use in aquaculture.
<Keywords: Marine sponge; Antifungal activity; Aquaculture; Fish pathogens
Emerging diseases caused by increasing number of virulent fungal infection in natural populations cause threat to plants, animals and humans. Fungal invasive plant diseases cause preharvest loss to fruit and cash crops causing tremendous loss to the crop production and agricultural output in natural habitats. It is reported that more than 600 million people could be fed each year by halting the spread of fungal diseases in the world’s five most important crops [1].
Aquaculture has also been a growing activity for more than 2 decades worldwide and fungal infections (mycoses) are amongst the common diseases in hatchery and aquaculture systems leading to the demise of fish population resulting in great economic loss [1]. Fortunately, in most fungal infection, since only the external tissue of fish and eggs are involved a good antibiotic agent can solve most of the major fungal infections. Considering the importance of ecofriendly health management in agriculture/aquaculture and concern about the contamination, toxicological and environmental risks posed by synthetic fungicides has led to an increase in the popularity of developing natural products as a source of ecofriendly compounds possessing antifungal activity. In addition, the evolving resistance of microorganisms to existing antibiotics is an important issue, not only in human research but also in agriculture and aquaculture. Hence, replacement of existing antibiotics with more effective and safer ones has become an important topic to address.
Marine sponges have been widely studied and shown to be highly successful in the production of metabolites possessing various activities such as antibacterial, antifungal, antiviral, antitumor, cytotoxic, antiinflammatory and several others [2]. Recently, antibacterial activity of sponge extracts have been studied against virulent fish pathogens [3,4]. In the similar manner, fungi that are resistant to antibiotics must be tackled with more effective, newer and safer antifungal agents. The present study thus aims at finding antifungal compounds from a marine sponge Negombata magnifica (formerly Latrunculin magnifica) against plant and fish fungal pathogens. Several studies report on cytotoxic and antibiotic properties of Latrunculin B using clinical pathogens. Hence, we report here the isolation and identification of the compound responsible for antifungal activity against plant pathogen and against fish pathogens. The MIC of the active compound was determined on the fish fungal pathogens and further tested for its toxic effects on shrimps.
Extraction and fractionation of sponge sample
The sponge sample Negombata magnifica was collected by hand using SCUBA from a depth of 15m off Hurghada in the Red Sea (27°15’26"N 33°48’46"E). The collected sponge sample was frozen immediately after collection and kept frozen at -20°C until processed. The frozen sponge sample was thawed and extracted exhaustively with acetone, pooled together and concentrated on a rotary evaporator. The acetone free aqueous extract thus obtained was transferred into a separating flask and fractionated sequentially using Diethyl Ether (DE) followed by Butanol (Bu) to obtain the DE-fraction and the Bu fractions respectively. The crude extracts were screened for antifungal activity.
Fungal pathogens
The fish pathogens (Exophiala salmonis, Branchiomyces demigrans and Saprolegnia sp.) were isolated from Tilapia, Oreochromis mossambicus collected from the Vellar estuary (lat. 11°29’N and Long. 79°46’E) south east coast of India. Exophiala salmonis and Branchiomyces demigrans were collected from the infected gills of the fish while Saprolegnia sp. was isolated from the infected skin tissue of the fish. Isolation was carried out using standard techniques. Briefly, one gram wet weight of the fish sample from the infected region was rinsed thrice in sterile sea water and homogenized using a sterile mortar and pestle in 5ml sterile seawater. Serial dilutions (upto 4 dilutions) were made and spread plated on Sabourauds dextrose agar (SDA, Hi Media) containing 50 mgml-1 of antibiotic chloramphenicol to inhibit bacterial growth. Plates were incubated at 26°C for 2-3 days. The isolates were repeatedly subcultured until pure fungal isolates were obtained and stored on SDA slants until use.
The phytopathogens were generously provided by ICAR (Indian Council of Agriculture Research), Goa, India. They were isolated from diseased parts (leaves, shoot, twigs) of fruit trees showing symptoms of disease. Briefly, the diseased part of the plant was rinsed in sterile distilled water and cut aseptically into small portions. These aseptically cut portions were placed on Potato Dextrose Agar (PDA) plates which were then incubated at 27°C and examined for visual growth of fungal hyphae. The fungal isolates were repeatedly subcultured until pure colonies were obtained. All the fungal pathogens (fish and phyto) were preserved on PDA slants for enumeration and identification at Agarkar Research Institute, Pune, India.
Purification of the active compound
A slurry of the Diethyl Ether (DE) fraction in silica gel was prepared by dissolving the crude extract in minimum quantity of DE and dried under nitrogen. This dry slurry was loaded onto a silica gel glass column and initially eluted with hexane followed by increasing concentration of diethyl ether in hexane. Next elution was performed using chloroform followed by increasing concentration of methanol in chloroform and finally eluted with methanol. Like fractions were combined on the basis of TLC and the combined fractions were subjected to bioassay screening against pathogenic fungi. Bioassay guided fractionation and finally purification of the active component was affected using High Performance Liquid Chromatography (HPLC).
Experimental
The 1H and 13C NMR spectra were recorded in CDCl3 on a Bruker DRX NMR spectrometer operating at 400MHz for 1H and at 75 MHz for 13C. Chemical shift (δ) values are expressed in parts per million (ppm) and are referenced to the residual solvent signals of CDCl3 (Table 3). Its mass spectrum was recorded in a positive ionization mode, using a QTOF-XL MS/MS, Applied Biosystem instrument equipped with MDS Sciex Analyst Software (Canada). Analytical TLC was performed on Merck pre coated silica gel 60 F254. Spots were visualized by spraying with ceric sulphate, followed by heating at 110°C for 2 minutes. Column chromatography was carried out using silica gel of 60-120 mesh size.
Biological activity
Antifungal activities invitro for the crude fractions and the pure compound against several plant and fish pathogens are summarised in (Table 1). The agar disc diffusion assay as described earlier [5] was used for performing the antifungal activity. Briefly, paper discs of 6mm diameter were impregnated with 25 μg of the crude extract and 10 μg of the pure compound dissolved in diethyl ether. Discs were placed on Potato Dextrose Agar (PDA) plates possessing a lawn of the different strains to be tested. The cultures were incubated for 48 hours at 27°C to obtain maximum growth in the culture media so as to visualize the clear zone of growth inhibition around each disc. The zone was measure in millimetre and scored as (- no activity; + mild activity; ++ moderate activity; +++ Significant activity; and ++++ strong activity). Standard disc of ketoconazole (10ug/disc) was used as a positive control and solvent chloroform was used as negative control.
| Sr. No. | Fungal Pathogens | Source | Accession no. |
|---|---|---|---|
| Plant pathogens | |||
| 1. | Macrophomina sp | Psidium guajava | BCC-PP001 |
| 2. | Helminthosporiumsp | Zea mays | BCC-PP002 |
| 3. | Pyricularia oryzae | Oryza sativa | BCC-PP003 |
| 4. | Sclerotium rolfsii | Solanum tuberosum | BCC-PP004 |
| 5. | Aspergillus niger gr | Archus Sapodilla | BCC-PP005 |
| 6. | Penicillium piceum | Archus Sapodilla | BCC-PP007 |
| 7. | Trichoderma koningii | Mangifera indica | BCC-PP009 |
| 8. | T. longibrachiatum | Mangifera indica | BCC-PP010 |
| 9. | Trichoderma atroviride | Anacardium occidentale | BCC-PP011 |
| 10. | Aspergillus oryzae | Triticum aestivum | BCC-PP014 |
| Fish pathogens | |||
| 1. | Exophiala salmonis | O. mossambicus | BCC-FiP001 |
| 2. | Branchiomyces demigrans | O. mossambicus | BCC-FiP002 |
| 3. | Saprolegnia sp | O. mossambicus | BCC-FiP003 |
Table 1: In vitro Antifungal activities of crude fractions and pure compound against plant and fish pathogens.
Minimum Inhibitory Concentration (MIC) for antifungal activity
Minimum Inhibitory Concentration (MIC) values of latrunculin B against test fungi (Table 2) were determined according to the Kirby and Bauer disc diffusion method [6]. The discs loaded with the compound were prepared in the same way as described above. After inoculation of 250μl (108spores/ml) solution of fungi onto potato dextrose agar and spread over the surface, an array of the discs containing different concentrations (μg/ ml) 24, 12, 6, 3, 1.5, 0.75, 0.375, 0.1875 and 0.093 of the compound was placed on the plates to determine the MIC values of the compound. Pure solvent-treated and dried discs were used as negative control discs and standard ketoconazole was used as positive control. The fungal plates were incubated for 48-72 h at 28°C. Minimum Inhibitory Concentration (MIC) is defined as the lowest concentration of the pure compound that inhibits visible growth of the microorganism around the disc. MIC was determined in duplicate (Figure 1).
| Fish pathogens | MIC values (μg/ml) |
|---|---|
| Exophiala salmonis | 1.50±0.1 |
| Branchiomyces demigrans | 0.75±0.1 |
| Saprolegnia sp | 3.0±0.3 |
Table 2: The MIC values of the pure compound, latrunculin B against fish pathogens.
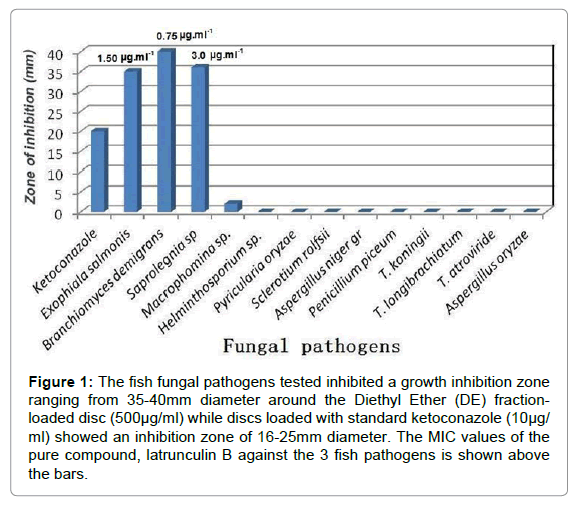
Figure 1: The fish fungal pathogens tested inhibited a growth inhibition zone ranging from 35-40mm diameter around the Diethyl Ether (DE) fraction-loaded disc (500μg/ml) while discs loaded with standard ketoconazole (10μg/ ml) showed an inhibition zone of 16-25mm diameter. The MIC values of the pure compound, latrunculin B against the 3 fish pathogens is shown above the bars.
Feeding-deterrence assay
The assay was performed as described by [7]. Briefly, the pure compound at a concentration of 5mg/ml was dissolved in acetone and added to a mixture of 50mg ground-lyophilized squid mantle, 30 mg alginate and 30 mg purified sea sand (granular size 0.1-0.3 mm). The mixture was homogenized using a mortar and pestle. The solvent was evaporated and permitted food red color dissolved in a few drops of distilled water was added to the mixture. The mixture was well mixed once again and exuded through a syringe into a 0.25mol/L CaCl2 solution. After 2-3 minutes, the hardened strand was briefly rinsed in seawater and cut into 10mm-long strips. Controls were prepared with acetone in the similar manner as described above. The shrimps collected and acclimatized in the laboratory were used for the assay using both compound and control in parallel. Experiments were performed using 10 shrimps in each 500ml plastic beakers containing 300ml of seawater in replicates of five. Only one colored food strip was given to each shrimp and the shrimps were not reused. The transparent exoskeleton of the shrimps enabled us to easily visualize the red food in the gut of the shrimps. The presence/absence of red spot in the stomach after 30 minutes indicated acceptance/rejection of the food while the dead/live shrimps after 24 hours indicated the toxic/nontoxic effect of the compound respectively.
Pathogenic fungi are having a pronounced effect on the global biota increasing the amount of fungal disease on them and this threat is ever increasing. Substantial data to support the idea that fungi pose a greater threat to plant and animal biodiversity relative to other taxonomic classes of pathogens is described and reviewed by [8]. Fungal diseases are second to bacterial infections that cause immense economic losses in aquaculture [9].
Crude Diethyl Ether (DE) and butanol (Bu) fractions of the marine sponge Negombata magnifica collected from the Red Sea was tested for its ability to inhibit the growth of fish and phyto-pathogens. In sharp contrast to the Bu fraction, which showed no activity, the DEfraction was strongly active to different extent as evident in Figure 1 Exceptionally significant activity was shown against all the three virulent fish pathogenic fungi; Exophiala salmonis, Branchiomyces demigrans and Saprolegnia sp. used in the study. The three cultures tested showed a growth inhibition zone ranging from 35-40 mm diameter around the DE fraction-loaded disc (500μg/ml) while discs loaded with standard ketoconazole (10μg.ml-1) showed an inhibition zone of 16-25mm diameter. Further bioassay guided isolation and purification of the Diethyl Ether (DE) fraction led to the identification of the active constituent Latrunculin B whose structure was elucidated using spectral data and confirmed by comparing its NMR data (Table 3, Figure 2) with literature values [10], thus confirming its structure. Its mass spectrum displayed a sodiated pseudomolecular ion (sodium adduct of molecular ion) at m/z 418.2755 [M+Na]+ indicating its molecular formula to be C20H29NO5SNa and its calculated mass is 418.1659. Latrunculin B was first isolated from the Red Sea sponge Negombata magnifica (formerly Latruncilin magnifica) by Kashmas’s group along with Lathrunculin A. This group is characterized by the presence of a macrolide1,3 fused to a tetrahydro-pyran containing a 2-thiazolidinone side chain. This compound has been widely studied for its anticancer and antitumor properties as well as against clinical pathogens and as antivirals [11]. As evident from Figure 1, the compound is practically inactive against phytopathogens; hence we highlight the use of this compound selectively against fish pathogens.

Figure 2: Structure of Latrunculin B and its 1H NMR spectrum.
| Position | 1H NMR (ppm) | 13C NMR (ppm) |
|---|---|---|
| 1 | - | 165.3 |
| 2 | 5.6, b,s | 117.7 |
| 3 | - | 154.6 |
| 4A | 2.66, m | 35.8 |
| 4B | 1.98, m | - |
| 5A | 2.34, m | 26.9 |
| 5B | 2.20, m | - |
| 6 | 5.25, m | 127.4 |
| 7 | 5.05, t | 135.8 |
| 8 | 2.68, m | 28.8 |
| 9A | 1.77, m | 31.2 |
| 9B | 1.12, m | - |
| 10 | 1.54, m, 2H | 31.3 |
| 11 | 4.25, t | 62.5 |
| 12A | 1.78, m | 35.3 |
| 12B | 1.52, m | - |
| 13 | 5.45, t | 68.7 |
| 14 | 2.14, d, 2H | 31.0 |
| 15 | - | 97.8 |
| 16 | 3.82, m | 61.2 |
| 17A | 3.46, m | 28.7 |
| 17B | 3.35, m | - |
| 18 | - | 174.8 |
| 19 | 1.91, s | 23.9 |
| 20 | 0.94, d | 22.2 |
| 15-OH | 5.3, s | - |
| NH | 7.01, s | - |
Table 3: NMR data of Latrunculin B in CDCl3a.
MIC of Latrunculin B for each fungus was as low as 1.50, 0.75 and 3.0 μg/ml for E. salmonis, B. demigrans and Saprolegnia sp. respectively (Table 2). Activity at such low concentration indicate its effectiveness as a lead molecule as an antifungal agent in aquaculture.
Fish infested with fungal pathogen is shown in Figure 3. E. salmonis known as black yeast infect fish and make them look darker and lethargic with erratic and abnormal swimming behaviour. Round yellow to white granulomas appear in the visceral organs like liver, kidney and spleen associated with prominent enlargement of the kidney [12]. Earlier reports on fish infected by E. salmonis are available [13-16].

Figure 3: Fish infected with fungal pathogen.
Fungal infection by Saprolegnia sp. causes saprolegniasis which is a most common presentation of water mould infection. They normally establish as small, focal infections, exhibiting white-to-brown tufts or cotton like growth on the skin, fins or gills causing superficial lesions. These especially affect fish which are debilitated (injured, diseased or stressed) and do not penetrate deep into muscles [17]. However, infection caused by Saprolegnia sp. can cause serious losses to spawns in hatcheries [18,19].
Branchiomyces demigrans causes branchiomycosis also called gill rot, which mainly affects the gill region of the fish leading to impaired oxygen exchange and osmoregulatory deficiency resulting in respiratory distress. The effect of parasitisation is reduction in gill raker count and gill surface area observed as a function of infestation. The maximum reduction in respiratory surface area was observed in the first gill arch and the minimum in the third gill arch. Infested fish had extremely pale gills, indicating severe anaemia and their gill rakers were seriously lost, apical edges damaged and out off gill lamellae heavily destroyed. Some secondary gill lamellae were fused or thickened. Between the gill lamellae, callus-like thickening was also observed. Although this is not a very common fungal infection, however, if it does occur, it can be deadly.
The study well explains the possible use of latrunculin B as a lead as antifungal agent in aquaculture further confirmed by its non toxicity to shrimps. Due to its various pharmaceutical applications and poor supply causing difficulty in obtaining steady supply of Latrunculin B, Hadas and group had attempted sponge sea ranching of the marine sponge Negombata magnifica for the mass production of latrunculin B [20].
In conclusion, we suggest that the secondary metabolite Latrunculin B produced by the marine sponge displayed promising potential as antifungal agent with specific reference to fish pathogens and hence could have application as lead compound in aquaculture and fish aquariums. In addition, since it is non-toxic to shrimps further shows that it would be safe in this application.
Authors are grateful to Dr S.W.A. Naqvi, Director NIO for constant encouragement. Sincere thanks to Dr P. Ramesh of ICAR, Goa for providing the fungal phytopathogens and Dr Singh of Agharkar Research Institute (ARI) for identifying the fungi. Gratitude and thanks to Dr Ernesto Mollo (ICN, Italy) for his valuable help in doing the feeding deterrent/shrimp toxicity assay. One of the authors (PD) sincerely acknowledges Dept of Biotechnology (DBT), Govt of India for the award of DBT-CREST post doc fellowship (2011). The authors are grateful to Dominique Melck for recording the NMR Spectra.