Forest Research: Open Access
Open Access
ISSN: 2168-9776
ISSN: 2168-9776
Research Article - (2017) Volume 6, Issue 3
When performing grafts, it is thought that by using plants of the same species the engraftment rate will be high; when the above is not fulfilled, the grafter, type of graft or the origin of the scions is blamed, but the anatomy of the parts forming the graft is not considered. The aim of this study was to evaluate the anatomy of four pine species used as rootstock and scion to define which species to use as rootstock based on cambium shape and bark thickness. Before grafting, 2-cm stem fractions were cut from plants that were used as rootstocks and scions. The evaluated species were Pinus patula, P. greggii, P. leiophylla and P. teocote. Cuts of 10 μm in thickness were made to determine bark and xylem thickness. Photographs were also taken with a photomicroscope to observe the "shape" of the vascular cambium. The experimental design was completely randomized with a 4x2 factorial arrangement, using 12 measurements as the sample size with four replicates. P. patula had greater bark thickness, while P. leiophylla obtained the greatest xylem thickness. In the images of the anatomical cuts, it was determined that the vascular cambium is continuous for all the species both in rootstock and in scions, but it is only circular in the P. teocote and P. leiophylla rootstocks, and the P. leiophylla scions did not present this feature. Based on the anatomy of the cambium, P. greggii, P. patula and P. teocote scions have greater possibilities of engraftment if they are grafted onto P. teocote andP. leiophylla because the cambium is circular. Tissue anatomy is one of the areas that must be known before making grafts in order to make appropriate decisions regarding the materials to be used.
<Keywords: Vascular cambium; Grafts; Pinus greggii; P. leiophylla; Pinus patula; Pinus teocote; Xylem
Grafting is the vegetative propagation method with the widest application in fruit trees, and it has also gained ground in the field of horticultural crops [1-3]. Sanou et al. [4] demonstrated that this technique can be used in wild species with more than 80 % engraftment.
This propagation technique has been used for different purposes in several species, for example in Passifora edulis Sims to eliminate the mosaic virus that causes fruit hardening [5], in Anona muricata L. to determine compatibility with the rootstocks used [6] and in Citrus sinensis L. to find the best interaction in three CTV-tolerant rootstocks and optimize tree development [7]. It is also used in tree species such as Pinus patula Schiede ex Schltdl. etCham. to produce seeds from superior trees [8].
In fruit and vegetables, grafting has been used to improve fruit quality, increase or reduce scion vigor, improve resistance to diseases and soil pathogens, and enhance tolerance to salinity, flooded soils, or high temperatures [9, 10, 11]. In forest species, mainly conifers, this technique has been basically used in breeding programs by establishing seed orchards from selected trees to produce seed that will generate trees with higher productivity [12,13].For edible seed production,Pinus pinea L. grafting has been used for more than 20 years [14].
A decisive factor in the failure of grafts is incompatibility, which refers to the lack of union between stock and scion, which affects the growth and productivity of the plants, and may even lead to their death [15-17].
Pereira et al.[16] mention that the incompatibility process involves anatomical, biochemical, physiological and molecular interactions between the rootstock and scion; this process can occur in a short time or years after the grafting [18].
In spite of the benefits of the grafting technique, pine propagation in Mexico is carried out by means of seeds, for reforestation and timber and non-timber plantations [19]. Recently asexual seed orchards have been established by grafting selected trees in the field. For example, Aparicio et al. [20] report 29 % engraftment in 3,000 Pinus patula grafts.
On the other hand, grafts have been made between different species in order to benefit from the characteristics of certain stocksthat are able to vegetate in certain areas where the crop has no opportunity to subsist on its own [12]. Grafting Pinus patula ontoP. radiata D. Don has caused the former to grow smaller in size without a significant reduction in the production of strobili, which could represent an advantage in cone harvesting[21].Climent et al.[22] showed that Pinus nigrasubsp. salzmannii(Dunal) Francoscions grafted ontoP. brutiaTen. had more male and female strobili per branch than whenP. nigraArnold stocks were used.
Hartmann et al.[23] mention that the success of grafts depends on several factors highlighted by:the ability of the grafter, the vitality of the scions andstocks and the environmental conditions. However, the need to study the anatomy of the rootstock and scion that are used is neglected, so the failure is attributed to the aforementioned factors.Darikova et al.[24] observed anatomical changes in the growth rings of 45- to 47-year-old grafts of P. sibiricaDu Tourand P. sylvestrisL., scion and stock respectively. Their interaction influences the diameter of the tracheids, the area of the lumen, the thickness and area of the cell wall and the production of cells in the growth rings, which modifies the distribution of growth regulators through the graft union.The species studied wereP. patula var. patula, P. greggii var. australis Donahue etLópez, P. leiophylla Schiede ex Schltdl. etCham. and P. teocoteSchiede ex Schltdl.etCham. The first two species belong to the serotine group [25], and are widely used in Mexican and international reforestation [26]. The last three species grow in sites that are more limiting than those of the first one, so they could confer resistance to water deficit or tolerance to soils less fertile than those of P. patula. P. teocote and P. leiophyllahave the ability to resprout when cut, and perhaps this ability serves to increase the grafting percentage of other species.
Therefore, the objective of this study was to evaluate and generate information regarding the anatomy of rootstocks and scions in four pine species and to define which species to use as rootstock according to these characteristics. For three of these species, there areno previous studies when used in grafts.
Plant material
Four pine species were used as rootstocks and scions:P. patula, P. greggii var. australis, P. leiophyllaandP. teocote. The scions were collected from trees with desirable genetic characteristics in sexual seed orchards located in Acaxochitlán, Hidalgo for P. patula, in Zacatlán, Puebla for P. greggii andin Texcoco, state of Mexico for P. leiophylla. The P. teocotescions are from superior trees in a natural stand located in Chignahuapan, Puebla. The donor trees were identified with the numbers 1, 2, 3 and 4 for P. greggii; 20, 39, 32 and 52 for P. patula; 10, 11, 162 and 90 for P. leiophylla and 1, 2, 3 and 4 for P. teocote. The approximately 1.5-year-old rootstocks were obtained from a forest nursery inIxtacamaxtitlán, Puebla, products of a mass seed lot.
Preparation of anatomical cuts
Stem samples from the scions and rootstocks of the four-pine species were collected and placed in 70 % alcohol. They were then fixed in FAA (10 % formaldehyde, 5 % acetic acid, 52 % 96° ethyl alcohol, and 33 % water) for 8 h; each sample was washed three times for 10 minutes with tap water, and then dehydrated with changes every 8 h in the following solutions: 96° ethyl alcohol at 30, 40, 50, 70, 85 and 100 % absolute alcohol 1: 100 % absolute alcohol 2 (1,2 represent two different absolute alcohol jars); 96° ethyl alcohol at 50%, 50 % xylene, 100 % xylene, 100 % xylene and 100 % xylene. They were then placed in paraffin 1 and 2 for 8 h each, at a temperature of 60-65 °C (1,2 represent two different paraffin jars); blocks were formed and accommodated. The blocks placed in paraffin were mounted on a wood block. Afterwards, 10-μm-thick cuts were obtained with an 820-rotary microtome (American Optical USA®).
The cuts were adhered to slides with chromium adhesive (to prepare 500 mL of chromium adhesive, 1.25 g of gelatin, 0.25 g of phenol, and 0.25 g of chrome alum were used) and placed on a plate at 60 °C for 12 h to ensure adhesion to the slides. The cuts were dewaxed in a xylene series (100 %) xylene 1, xylene 2 and xylene 3, absolute alcohol 1 and 2, alcohol 85, 70 and 50 %; they were then placed in safranin in a saline solution for 1 h minimum (0.05 safranin, 2 % granulated salt, dissolved in water), alcohol 30, 50, 70, 85 and 100 %. Afterwards, the slides containing the samples were removed one by one and a green dye was applied with a dropper for 10 to 20 seconds (the excess was removed using a wash bottle and absolute alcohol). It was again subjected to 100 % alcohol, xylene 1, xylene 2 and xylene 3. Synthetic resin was applied and the slide coverslips were put in place; they were left on the plate for 24 h at 60°C.
Measurement of anatomical cuts
The anatomical cuts were observed and photographed with a Carl Zeiss photomicroscope III (2x) with a digital camera for PAXcam 3 microscopy. Afterwards, bark and xylem thickness measurements were performed using ImageJ software (version 1.48).
Experimental design
The experimental design was completely randomized with a factorial arrangement with eight treatments (4 × 2), resulting from four pine species and two conditions (rootstock and scion). The sample size used was 12 anatomical cuts with four replicates. Tukey’s range test (p ≤ 0.05) was performed with the SAS 9.0 statistical package.
Bark thickness
The average bark thickness of the four species was 622 μm, and ranged from 511.48 in P. greggii to 823.26 μm in P. patula, whereas in the rootstocks the mean was 376 μm, 65 % lower than the scions, and ranged from 312.30 in P. greggii up to 527.48 μm in P. teocote. By observing the test of means in the bark, the P. patula scion statistically outperformed all rootstocks and scions evaluated. P. patula had the greatest thickness (823.26 μm), while the lowest value was for P. greggii, with a 163 % difference (Table 1). The rootstocks have lower bark thickness than the scions (Figure 1). In P. teocote the rootstock bark is similar to that of the scion, with only a 9 % difference between them; this characteristic can mean an advantage at the moment of grafting, since at the moment of joining both parts they present a higher matching possibility, whereas in P. patula the largest difference between the two parts used for grafting, 155 %, occurred.
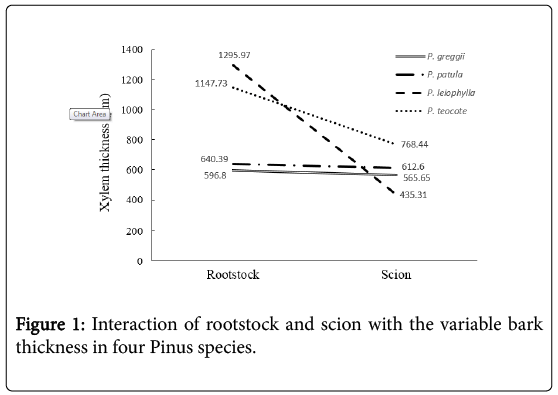
Figure 1: Interaction of rootstock and scion with the variable bark thickness in four Pinus species.
| Species | Condition | Thickness (µm) | |||
|---|---|---|---|---|---|
| Bark | Xylem | ||||
| P. greggii | Rootstock | 312.30 ± 10.48 | e | 596.79 ± 8.06 | de |
| Scion | 511.48 ± 10.33 | c | 565.65 ± 9.56 | e | |
| P. patula | Rootstock | 322.52 ± 4.82 | de | 640.39 ± 10.01 | d |
| Scion | 823.26 ± 13.89 | a | 612.60 ± 8.55 | de | |
| P. leiophylla | Rootstock | 341.34 ± 3.93 | d | 1295.97 ± 16.30 | a |
| Scion | 578.36 ±12.51 | b | 435.31 ± 14.21 | f | |
| P. teocote | Rootstock | 527.48 ± 8.94 | c | 1147.73 ± 35.07 | b |
| Scion | 573.98 ± 11.31 | b | 768.44 ± 16.31 | c | |
| CV | 14 | 15.48 | |||
Table 1: Bark and xylem thickness in four pine species used as rootstocks and scions.
Xylem thickness
The average xylem thickness of the scions in the four-pine species was 595.50 μm, ranging from 435.31 to 768.44 μm for P. leiophylla and P. teocote, respectively. In the rootstocks the mean was 920.22 and the variation from 596.79 to 1295.97 μm, for P. greggii and P. leiophylla, respectively, being opposite to the behavior observed in bark. The test of means showed that the xylem thickness in P. leiophylla (rootstock) was statistically superior to that in the other scions and rootstocks, and the same species had the lowest thickness with a 198 % difference in relation to the former (Table 1). In the xylem thickness × species interaction, the behavior is opposite to that of bark thickness, where the stocks used have greater xylem thickness than the scions (Figure 2). In P. teocote the bark thickness is practically the same in the rootstock and in the scion (Figure 1). Whereas P. patula is the one with the greatest contrast, while P. greggii and P. leiophylla show similar values.
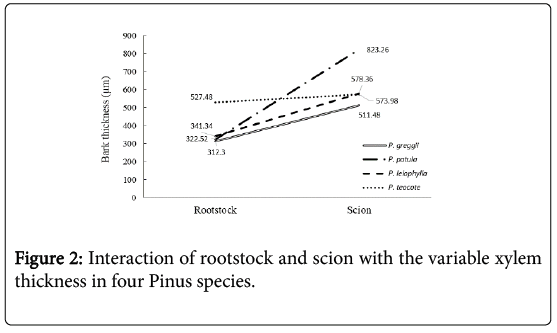
Figure 2: Interaction of rootstock and scion with the variable xylem thickness in four Pinus species.
The rootstock-scion correlation for xylem thickness shows that P. patula and P. greggii have similar behavior and there are no differences in xylem thickness between scion and rootstock. On the other hand, P. leiophylla shows the biggest difference, as the thickness of the rootstock xylem exceeds that of the scion by 198 %; in P. teocote this difference is 49 % (Figure 2). Means with the same letter in each column are statistically equal (Tukey p ≤ 0.05); CV= coefficient of variation.
Shape of Vascular Cambium
The conditions of the rootstock and scion in the studied species show different anatomical characteristics; the vascular cambium, although continuous in all cases, is not circular in all species.
The anatomical cuts of the scions with the exception ofP. leiophylla (Figure 3-6) show the vascular cambium continuous and with some circularity. On the other hand, therootstocks showing circular cambium are P. leiophylla (Figure 5B) and P. teocote (Figure 6B), the latter having the least undulations.
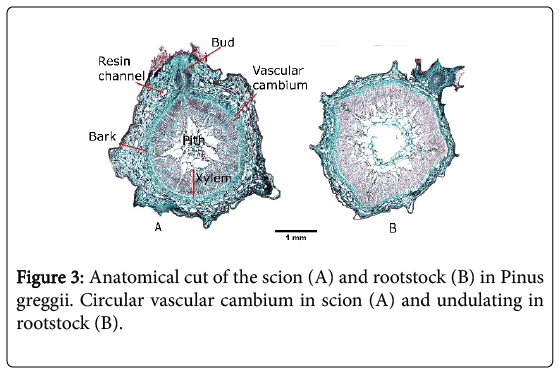
Figure 3: Anatomical cut of the scion (A) and rootstock (B) in Pinus greggii. Circular vascular cambium in scion (A) and undulating in rootstock (B).
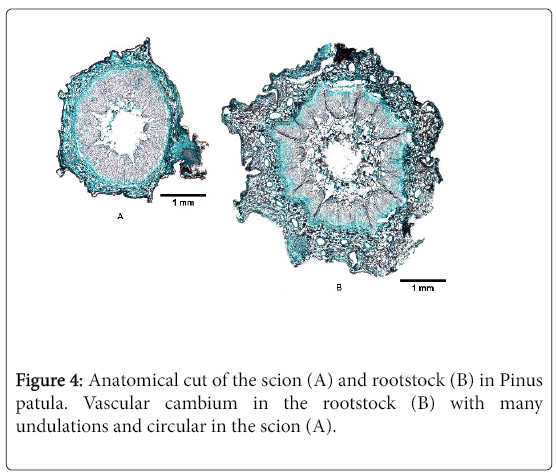
Figure 4: Anatomical cut of the scion (A) and rootstock (B) in Pinus patula. Vascular cambium in the rootstock (B) with many undulations and circular in the scion (A).

Figure 5: Anatomical cut of the scion (A) and rootstock (B) in Pinus leiophylla . Vascular cambium with undulations and a greater proportion of bark in scion (A).
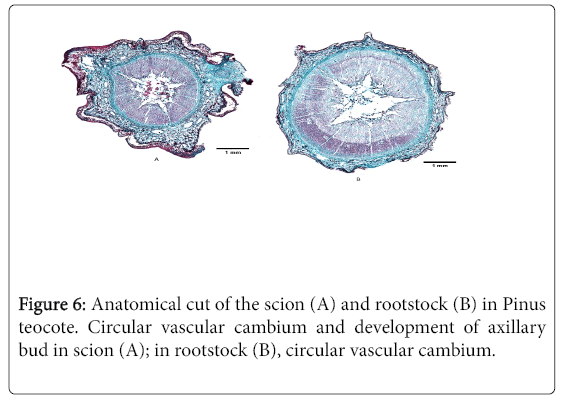
Figure 6: Anatomical cut of the scion (A) and rootstock (B) in Pinus teocote. Circular vascular cambium and development of axillary bud in scion (A); in rootstock (B), circular vascular cambium.
By comparing the anatomy of thecambium in Pinus greggii (Figure 3A and 3B), it can be seen that the cambium of the scion is circular except for two points where two buds develop. It is important to note that in pine, because the internodes are short, the formation of budscan modify the shape of the cambium. The rootstock anatomical cut (Figure 3B) shows that the cambium is irregular.
The cambium in the P. patula scion is continuous and oval due to the shape of the graft, with a projection at the top due to the development of a bud, whereas the cambium of the rootstock has undulations towards the interior of the xylem and protuberances that may be related to the development of axillary buds (Figure 4A and 4B).
(Figure 5A) shows the cut of the Pinus leiophylla scion, which is the most irregular of all the pine species, with small undulations. On the other hand, the rootstock cambium is circular and the bark thin and uniform, which could be an advantage at the time of grafting, and have a major probability that the vascular tissues coincide because they are circular and continuous (Figure 5B).
The cambium of the P. teocote scion like P. greggii is circular with a small protuberance due to the development of an axillary bud; these two species have the most circular cambium of the four-species studied. The cambium of the P. teocote rootstock is circular and together with that of P. leiophylla are the ones that show better anatomical characteristics in relation to the shape of the cambium.
According to Hartmann et al. [23], three characteristics must be fulfilled in order to be able to graft a plant: a circular vascular cambium, a prompt response to the cut and the ability to form callus in the graft union; therefore, to know the first characteristic it is necessary to study the anatomy of the species used. However, the cambium is not always circular, as seen in the images of the present study. Salcedo et al.[27] showed that the cambium in Pouteria sapota(Jacq.) Moore & Stearn has morphological irregularities and discontinuity in the ring of cells in the area near the apical bud, resulting in dead tissue in the graft union.
When studying the anatomy in grafts, the number of vessels or tracheids per square millimeter, and radial and tangential vessel diameter in the xylem are mainly studied; in the same way, the study of anatomy can provide an explanation as to how it influences the growth of shoots in grafted trees [28, 29].
Determining the anatomy of scions and rootstocks prior to grafting is useful because it allows knowing the basic structures that will be in contact, so that there is a connection between the two parts, thereby avoiding anatomical incompatibility. This incompatibility occurs because of a poor vascular connection, vascular discontinuity and degeneration of the phloem in the graft union area [30,31], causing an imbalance in the transport of water and nutrients that affects plant growth [2,32].Kawaguchi et al. [33] mention that solanaceous plants, the vascular bundles discontinuity in the graft union area affected the translocation of water and nutrients between rootstocks and scions.
In the case of pines, little is known about the anatomy of rootstocks and scions of the species used, so the low engraftment percentageis attributed to the ability of the grafter, the type of graft used, or errors in post-graft management. Copes [34] mentions that in Pseudotsuga menziesii (Mirb.) Franco the development of vascular cambium in 80% of the grafts happens from day 17 to 23 after being grafted; however, when the rootstock and scion have different diameters,its formation can be delayed by up to three months.
Soto et al. [35] obtained 95% engraftment in grafts of Jatropha cinerea (Ortega) Muell. Arg. as rootstock and Jatropha curcas L. as scion, observing that the vascular cambium of both species is continuous and circular. In addition,J. cinerea has wider cambium than J. curcas, which favored the formation of callus for cell regeneration in the union area.
In anatomical cuts in Litchi chinensis Sonn. cv "Jingganghongnuo" as scion and 13 cultivars as rootstocks, the compatibility of some of these combinations is shown by the perfect insertion of the scion and rootstock, in addition to the development of new vascular cambium [36]; this may be due to the rootstock having circular and continuous cambium, as observed in P. teocote (Figure 6B).
The anatomy of the four-pine species studied indicates that this is a factor that must be taken into account before grafting, to avoid anatomical incompatibility. On the other hand, it shows that P. teocote and P. leiophyllahave greater possibilities of being used as rootstocks because they present the circular characteristic in the cambium, if they are grafted withP. greggii, P. patulaandP. teocotescions.
The above must be corroborated with later anatomical studies showing the anatomical development of the union of these combinations, complemented with nutritional and physiological studies.
The anatomy in pine grafts is a poorly explored area of study in which more detailed studies are still needed to understand the success or failure of the grafts.
The shape of the vascular cambium in pines can be circular or undulated.
Pinus teocote and P. leiophylla can be used as rootstocks due to the circular shape of the vascular cambium.
The vascular cambium in scion of P. greggii, P. patula y P. teocote is circular.
The rootstocks used generally have lower bark thickness than the scions.