Journal of Hematology & Thromboembolic Diseases
Open Access
ISSN: 2329-8790
ISSN: 2329-8790
Research Article - (2016) Volume 4, Issue 2
Keywords: CDBPA; BL; Apoptosis; Tumor growth; Gene expression; Survival
Burkitt’s Lymphoma (BL) is a rare, aggressive and fast growing human tumor and was the first human tumor to be associated with an Epstein-Barr Virus (EBV) [1]. It is characterized by the translocation and deregulation of the c-myc oncogene on chromosome 8 [2]. BL is characterized by monomorphic medium-sized B cells with basophilic cytoplasm and numerous mitotic figures [3,4]. Three types are categorized: Endemic lymphoma refers to those cases occurring in African children (usually 4-7 years old, with a male: female ratio of 2:1) Sporadic BL occurs worldwide; it accounts for 1% to 2% lymphoma in adults and up to 40% of lymphoma in the U.S. and Western Europe [5]. Immunodeficiency-associated BL is usually associated with Human Immunodeficiency Virus Infection of the use of immunosuppressive drugs [6].
Patients often present with bulky disease and a high tumor burden due to a short doubling time of the tumor. Seventy per cent of patients present at advanced stages. BL is fatal if left untreated, but BL is curable. Concerning treatment of BL there are no authorized treatment guidelines for this condition available. There are individual clinical and research publications that support treatment regimens like CHOP, RCHOP R-CCODOX-M/R-IVAC, R-BFM and R HyperCVAD/HDAraC+ Methotraxate [7-9].
Cisplatin is currently recommended for treatment of second line patients [10,11]. Since BL is classified as a non-Hodgkin lymphoma (NHL) and therefore significant benefit need to be demonstrated in the context of the following products indicated for NHL: pixantrone demaleate, chlorambucil, cyclophosphamide, cytarabine, doxorunicin, bendamustine, lomustine, methotrexate, minoxantrone hydrochloride and vincristine sulfate.
Rituximab is only indicated for CD20 positive lymphomas. Meabco A/S Copenhagen, Denmark has recently developed a new anti-cancer complex Cis-diaminneplatinum (II) and an amphiphilic polymer containg a mix of benzene poly-carboxylic-acids (CDBPA).
The complex has an anti-cancer tumor growth efficacy and improves quality of life, decreased number of adverse effects and decrease tumor toxicity in patients with stage IV breast cancer patients [12-16]. The aims of the present study were to investigate, if the text article CDBPA has an inhibitory effect on Ramos cells in vitro and to describe the effects of CDBPA on cell cycle, apoptotic pathways and its gene expressions in vitro. Furthermore, we also tested whether CDBPA were able to suppress human lymphoma growth in a NOD-scid mouse xenograft transplantation model, significantly.
Chemical and biological reagents
Cell culture media and reagents were purchased from Biological Industries (Beit Haemek, Israel). BP-C1 was obtained free of charge from Meabco, Copenhagen, Denmark. BP-C1 is a benzene-polycarboxylic complex with cis-diammineplatinum [II] dichloride. This is a polymer complex of carbonic and oxicarbonic acids rich in carboxylgroups replacing the chlorine ions. One ml of BPC1 monoclonal antibody was purchased from Oncogene (Boston, MA, USA). Anti actin monoclonal antibodies was purchased from INC Biochemicals (Aurora, OH, USA). Secondary antibody peroxidase–conjugated goat anti–mouse IgG was purchased from Jackson Immune Research Laboratories (West Grove, PA, USA). All other chemicals were purchased from Sigma (Saint Louis, MO, USA) or other local sources.contains 0.5 mg of platinum-ammonium salts of benzenepolycarbonic acids, including 0.05 mg of platinum. Anti–caspase 3
Animals
Hundred twenty (120) female NOD-Scid mice (nomenclature: NOD/MrkBomTacPrdkcscid) 5 weeks of age were supplied by Taconic- Europe A/S. The cancer test model was established by xenotransplantation of human cancer cells to a s.c. site ar right flank of the mouce. In the present model 10×106 Ramos cells (ATTC number: CRL-1923) were combined with matrigel (1:1) and inoculated s.c. as 200 μL solution. Danish animal experiments license number: 2012-15-2934-00051 C1.
Cell culture
Hodgkin lymphoma cell line (RAMOS) was purchased from American Tissue Culture Collection. Cells were cultured at 37 °C in a humidified 5% CO2 atmosphere in DMEM medium supplemented with 100 U/ml penicillin, 100 mg/ml streptomycin, 2 mM of Lglutamine and 10% fetal calf serum (Biological Industries, Beit Haemek, Israel). Cells were cultured as described elsewhere [12].
Cell viability
Cells were seeded in 96-well plates (2×103 cells/well). The following day, cells were treated with BP-C1 in a spectrum of concentrations ranging from 10 to 150 μg/ml for 72 hours and cell viability was detected using Cell Proliferation Assay, XTT (Biological Industries, Beit Haemek, Israel).
This assay is based on the ability of metabolic active living cells to reduce the tetrazolium salt, XTT, to orange colored compounds of formazan. The absorbance of each sample was measured using an ELISA reader (Tecan, Spectra) at a wavelength of 450 nm with a reference absorbance of 620 nm. The results are presented as percentage of control and expressed as means ± standard deviation of three independent experiments in which each treatment was performed in triplicate.
Lactate dehydrogenase (LDH) release
Cellular damage, such as necrosis, causes an elevation of the LDH concentration in the medium. The integrity of the plasma membrane following treatment was determined by measuring LDH activity released into the culture medium as described before [17].
Cell cycle experiments
Cell cycle experiment was carried as described before [12]. Briefly, cells (2×106) were incubated for 48 h, with CDBPA - at concentrations of 10-150 μg/well. After treatment, cells were harvested by centrifugation, washed twice in cold phosphate buffer saline (PBS; Biological Industries, Israel) and centrifuged for 5 min at 2000 rpm and 4 °C. The cells pellet was fixed in cold ethanol (70%), (Frutarom, Israel) at -20 °C for overnight. Prior to analysis, ethanol was removed by centrifugation, and cells were treated with 0.1% NP-40 (Sigma, Saint Louis, MO, USA) for 5 min on ice. Cells were rinsed again and treated with RNase (Sigma) (100 μg/ml PBS) for 20 min at 4 °C and stained with propidium iodide (PI; Sigma), (50 μg/ml) for 20 min. Cell cycle analysis was performed using an automated flow cytometer (Becton Dickinson FACS Calibur, BD Biosciences), managed with CellQuest software. Sub-G1 appearance was used to quantify cells containing fragmented DNA.
Annexin V-FITC double-staining assay
Cells (1×106) were incubated for 24, 48, or 72 h with 100 μg/well of CDBPA. Cells were harvested by centrifugatuon, washed and suspended in cold PBS. Cell death was analyzed by double staining the cells with FITC-labeled Annexin V and Propedium Iodide (PI), using an Annexin V-FITC apoptosis detection kit, (BD Biosciences, San Jose, CA, USA), according to the manufacturer’s instructions as described elsewhere [12]. Cells were analyzed by flow cytometer (BD FACSCanto II), managed with FACSDiva software.
Western blot analysis
Proteins were extracted, using a RIPA buffer (1% NP-40, 0.5% sodium deoxycholate, 0.1% SDS, 1 mM EDTA) containing protease inhibitors (30 mg/ml aprotinin, 2.5 mg/ml leupeptin, 10 mg/ml pepstatin and 1 mM phenylmethyl fluoride) as previously described [12]. Western blot analysis was performed for the assessment of activation of Caspase-3, calibrated by actin. Cells (2×106) were incubated for 48 h with 100 μg/well of CDBPA. Following treatment, cells were harvested by centrifugation, rinsed by PBS, and lysed for 20 min on ice by glycerol lyses buffer [0.05 M 4-(2-hydroxyethyl)-1- piperazineethanesulfonic acid (hepes; sigma Saint Louis, MO, USA), 0.5% NP-40, 0.002 M Ethylendiaminetetra-acetic acid (EDTA), 0.25 M NaCl and 10% glycerol]); supplemented before use by 40 μl/ml protease inhibitor cocktail (Roche, Mannheim, Germany) and 1 μl/ml phenylmethylsulfonyl fluoride solution (PMSF; sigma, Saint Louis, MO, USA) (0.3 M).
Protein estimation of supernatants was carried out using the Bradford assay. Equal concentrations of protein samples (40 μg) were separated by SDS-PAGE, transferred to nitrocellulose membrane and blocked in 5% non-fat dry milk (Bio-Rad, hercules, CA, USA) in TBS/T buffer. Membranes were probed with rabbit polyclonal anticaspase- 3 antibody at 1:5000 (Abcam, Cambridge, UK).
The secondary antibody was peroxidase conjugated goat-anti-rabbit IgG at 1:10000 (Jackson Immune Research Laboratories). The membrane was incubated with EZ-ECL kit (Biological Industries, Israel) and the enhanced chemiluminescence was detected using gel imaging ChemiDoc™ XRS + System with Image Lab™ software (Bio- Rad). Membranes were re-probed with mouse monoclonal anti-actin antibody at 1:20000 (MP Biomedicals).
Gene expression analysis
Gene expression following treatment with 100 μg/well of CDBPA was detected using the Applied Biosystems® TaqMan® Array Plates (Applied Biosystems, Foster city, CA). A 96-well Plate, which contained 92 assays for apoptosis associated genes and 4 assays to candidate endogenous control genes, was used. The panel of assays in this plate targets genes from both the signaling pathways that initiate mammalian apoptosis, the death receptor regulated pathway, and the BCL-2 family pathway. After treatment, total RNA was isolated using a TRIzol reagent (Invitrogen, Carlsbad, CA). 1 μg of RNA was reverse transcribed using a reverse transcription system according to the manufacturer’s instruction. The housekeeping genes, HPRT1 and GAPDH, were used for internal control to correct the potential variation in RNA loading. All reactions were performed in a 25 μl volume containing the cDNA sample, TaqMan fast universal PCR mastermix, primers, and probes. Before the PCR cycles, samples were incubated for 2 min at 50 °C and for 10 min at 95 °C. Thermal cycles consisted of 45 cycles at 95 °C for 15 s and 65 °C for 1 min.
In vivo experiments
To study the efficacy of cancer treatment with test article CDBPA, 120 mice were inoculated with 10×106 Ramos cells 12 days before scheduled start of administration start of administration of the test article and reference animals. Female NOD-Scid mice (Taconic, Europe) were used tumor host animal. The Ramos cells were purchased from American Tissue Culture Collection. They were inoculated in a 200 μL solution containing 50% phosphate buffered saline (PBS) and 5+% matrigel solution. The treatment was conducted with a cohort of 75 animals with tumors with an average size of 319.7 mm3, which was the volume obtained 12 days after inoculation of the cells. The cohort of 75 animals were allocated to 5 groups with 15 animals in each group to have groups with approximately equal group mean values of tumor volume. The treatment with the test article and reference groups were started the same day as allocation to groups and then continued 3 times per week on Monday’s, Wednesday’s and Friday’s for 4 weeks or until humane end point were reached. Group 1 was administered with saline (s.c.) and respresenting the non-treated group, group number 2 was administered with CDBPA (31.5 mg/kg s.c.), group number 3 was administered with CDBPA (47.0 mg/kg), group number 4 was administred with CDBPA (62.5 mg/kg) and group number 5 was administered with the treatment regime R-CHOP (Rituximab 30 mg/kg, cyclophosphamide 40 mg/kg, doxorubicine 4 mg/kg, oncovin(vincristine) 0.5 mg/kg, prednisolone 0.2 mg/kg. Cyclophosphamide, doxorubicine, and oncovin were administered i.v. as a mixture once every 7 days and rituximab was administrered i.p. every 7 days where first day was the days of allocation into groups [14,15]. Prednisolone was administered p.o. once at the day of administration of the R-CHOP and then the following days which is a total of 5 days of administration per week. The body weights of the animals were recorded once a week and at increased frequency in case of appearance of clinical adverse symptoms. The tumor volume was estimated from recording of tumor length (L) and tumor width (W) by using the formula (L×W2) /2 and assuming that the tumor is a sphere.
Statistical analysis
Comparison of groups with regard to development in tumour size was carried out by using Analysis of Variance (ANOVA) with repeated measurements and Day 12 covariates. Kaplan Meier survival plot comparing each of the study groups 2 (CDBPA 32.5 mg/kg) and 3 CDBPA 47.0 mg/kg) with the vehicle group at 2 different endpoint settings. In panel a) and b) the endpoint (tumor size that defines death of cancer) is defined as 1000 mm3 and in panel c) and d) is the endpoint is defined as 1688 mm which also is the humane endpoint for tumor size in the present study. Statistical differences between the pairs of curves were analyzed by a Gehan-Breslow-Wilcoxon test and the P-values are indicated in each panel; p < 0.05 was considered as significant [18-20].
In vitro studies
Cell viability
The effect of CDBPA on viability of human Ramos cells was examined. The results indicated that treatment with CDBPA, 25 μg/ well for 72 h significantly (p < 0.001) reduced cell viability of Ramos cells. Cell viability was reduced by ~60% and ~85% following treatment with 25 or 50 μg/well, respectively (Figure 1). Treatment of the cells with 100 or 150 μg/well reduced cell viability by more than 95% (Figure 1).
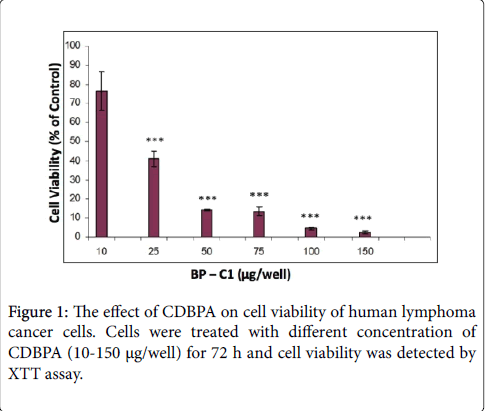
Figure 1: The effect of CDBPA on cell viability of human lymphoma cancer cells. Cells were treated with different concentration of CDBPA (10-150 μg/well) for 72 h and cell viability was detected by XTT assay.
No significant differences were observed in extracellular LDH levels at the concentrations of 50-150 μg/well compared to untreated cells (Data not shown). These results indicate that BP-C1 does not cause LDH leakage in RAMOS cancer cells .
Cell cycle analysis
Subsequently, in order to study the effect of BP-C1 on cell cycle progression, fluorescence-activated cell sorting (FACS) analysis was performed. Treatment of the cells with the active substance 100 μg/ml resulted in a reduction in G1, S and G2/M The distribution of the cells in G1 phase in control vs treated cells were 57% vs 12.3%, respectively. On the other hand distribution of the cells in sub-G1 phase was 7.6% in control and 64% in treated cells. These results may suggest an occurrence of cell deathphases of the cell cycle and an accumulation of the cells at sub-G1 phase (Figure 2).
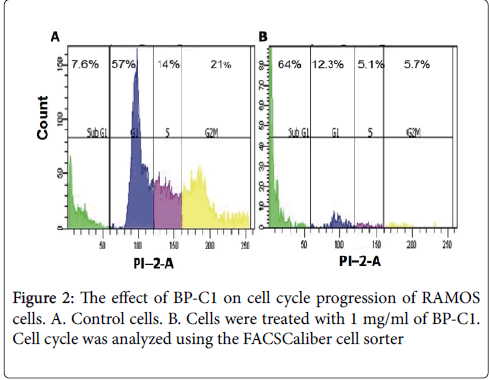
Figure 2: The effect of BP-C1 on cell cycle progression of RAMOS cells. A. Control cells. B. Cells were treated with 1 mg/ml of BP-C1. Cell cycle was analyzed using the FACSCaliber cell sorter
Detection of apoptotic cells by annexin V assay
Annexin V binds phosphatidylserine, which becomes externalized on the outer plasma membrane during early stages of apoptosis. On the other hand, PI binds to cellular DNA of cells during necrosis or later stages of apoptosis. RAMOS cells were incubated for 48 h with 100 μg/ml CDBPA. Cells were harvested and stained with annexin V and PI. The number of intact cells was significantly decreased in the treated cells (29.6%) with CDBPA compared to control cells (71.1%) as shown in Figure 3. Treatment with CDBPA significantly increased the number of Annexin V-FITC positive/PI-positive cells. Respective percentages of these cells (Q2+Q4) in control and treated cells for 48 h were ~28.6%, vs ~69.8% respectively.
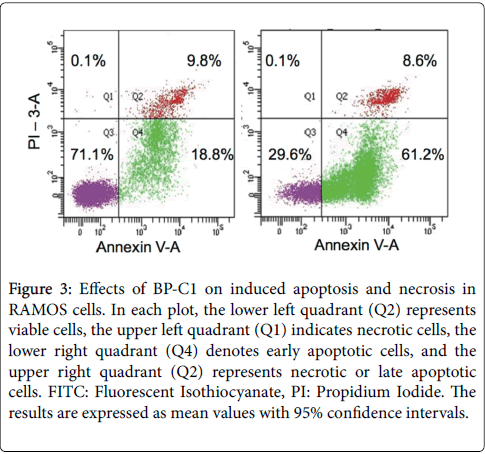
Figure 3: Effects of BP-C1 on induced apoptosis and necrosis in RAMOS cells. In each plot, the lower left quadrant (Q2) represents viable cells, the upper left quadrant (Q1) indicates necrotic cells, the lower right quadrant (Q4) denotes early apoptotic cells, and the upper right quadrant (Q2) represents necrotic or late apoptotic cells. FITC: Fluorescent Isothiocyanate, PI: Propidium Iodide. The results are expressed as mean values with 95% confidence intervals.
Effect of BP-C1 on gene expression
Following detection of apoptotic cells, the level of the effector caspace 3 was detected. The results indicated that BP-C1 has no effect on activation of caspase 3 (data not shown). Therefore, the effect of CDBPA on the expression of genes involved in apoptosis was detected using the Applid Biosystems® TaqMan® Array–KIT. Gene expression was quantified by qRT-PCR as described under “Materials and Methods” (Figure 4A).
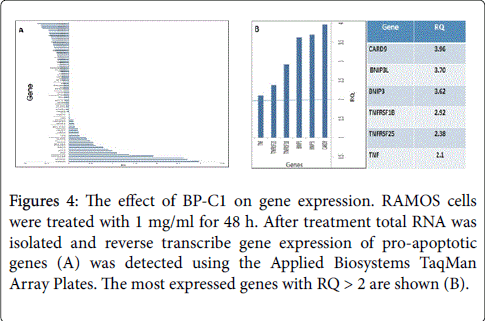
Figure 4: The effect of BP-C1 on gene expression. RAMOS cells were treated with 1 mg/ml for 48 h. After treatment total RNA was isolated and reverse transcribe gene expression of pro-apoptotic genes (A) was detected using the Applied Biosystems TaqMan Array Plates. The most expressed genes with RQ > 2 are shown (B).
The results revealed higher levels of mRNA transcripts of proapoptotic genes with RQ > 2; CARDS9 (RQ: 3,96), BNIP3L (RQ: 3.62), TNFRSF1B (RQ: 2.92), TNFRSF25 (RQ 2.38) and TNF (RQ: 2.1) (Figure 4B). On the other hand, gene expression quantified by qRTPCR revealed lower levels of mRNA transcripts of apoptotic-inhibitory genes; BCL2L13 (RQ: 0.69), BCL2L1 (RQ: 0.93) and XIAP (RQ: 0.83).
In vivo studies
CDBPA was administered in three doses (31.5, 47 and 62.5 mg/kg and the effects were compared to groups treated with either vehicle or reference treatment R-CHOP [14,15].
The outcome of the study confirmed that study group 3 with medium concentration of CDBPA (47 mg/kg) causes significant reduction in tumor growth over the 2-week treatment period (Figure 5). At study day 26 (2 weeks of treatment) the mean tumor volume (n=14) was 53% of the mean tumor volume of study group 1 (vehicle).
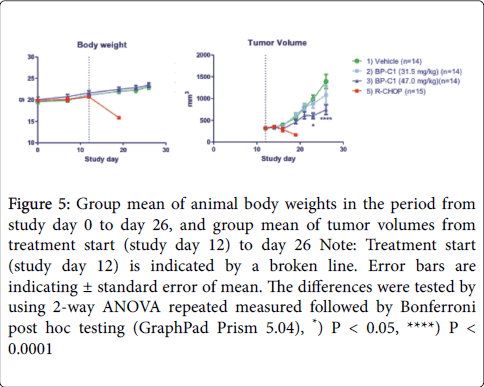
Figure 5: Group mean of animal body weights in the period from study day 0 to day 26, and group mean of tumor volumes from treatment start (study day 12) to day 26 Note: Treatment start (study day 12) is indicated by a broken line. Error bars are indicating ± standard error of mean. The differences were tested by using 2-way ANOVA repeated measured followed by Bonferroni post hoc testing (GraphPad Prism 5.04), *) P < 0.05, ****) P < 0.0001
The R-CHOP treatment caused an approximately 50% reduction of the mean tumor volume size during one week of treatment, however, the animals did loose body weight to a degree that required termination due to animal ethical endpoint (Figure 5 and 6). Study group 4 (CDBPA 62.5 mg/kg) was terminated due to extensive swelling at the place of S.C. injections. Besides the swelling at the injection site, study group 4 showed no clinical symptoms and the body weights matched the body weight development of the vehicle treated cohort (Figure 6).
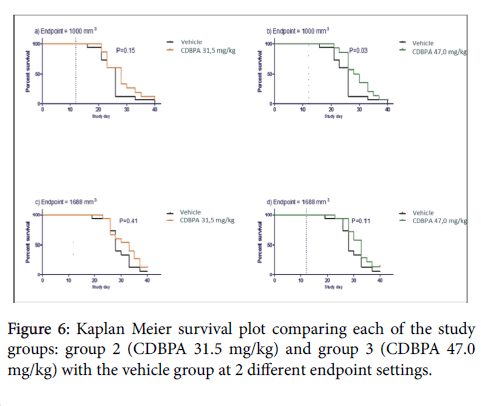
Figure 6: Kaplan Meier survival plot comparing each of the study groups: group 2 (CDBPA 31.5 mg/kg) and group 3 (CDBPA 47.0 mg/kg) with the vehicle group at 2 different endpoint settings.
The results of this study indicated that treatment of mice inoculated with Ramos cells with 47 mg/kg of the test article CBPDA caused a significant reduction of the tumor growth in a 2-week treatment period in NOD-scid mice with experimental Ramos cell B [21-30]. The mean tumor volume of the treated animals was 53% of the mean tumor volume of the vehicle group. The R-CHOP treatment caused a reduction to approximately half tumor size during one week of treatment, however the animals did lose body weight to a degree that required termination due to humane endpoint.
The CBPDA has been tested in clinical trials and has shown to exert anti-tumor activity in breast cancer patients stage IV [13-16]. To understand whether this anti-tumor activity is associated with a direct effect on tumor cells and/or indirectly through activation of the immune system, we have tested the effect of CBPDA on human peripheral mononuclear cells comprising lymphocytes and monocytes, as well as cytotoxic lymphocytes with specificity to tumor cells [15,16]. CBPDA demonstrated that it stimulated the production of cytokines such as TNF-alpha, IL-1 beta, GM-CSF, IL-6, Il-25 and INF-gamma [31]. All these cytokines increases the anti-tumor activity through multiple mechanisms. One mechanism may involve activation of dendritic cells, as all the above mentioned cytokines except IL-25 are able to induce differentiation and maturation of dendritic cells [32]. In addition IL-25 may exert direct cytotoxic/cytostatic activity of monocytes. Furthermore INF gamma production may also be able to activate direct anti-tumor activity of monocytes [33]. A gene expression profile analysis has been performed on Ramos indicated that CBPDA is significantly increased the expression of five genes that have an important role in cell apoptosis; CARD9, BNIP, TNFRSF-1B, TNFRSF25 and TNF-alpha. CARD9 positively regulates apoptosis and NF-κB activation thereby activating the pro-inflammatory cytokines (TNF, IL-25, IL-6, IL-2) and the anti-inflammatory cytokine (IL-10) subsequently resulting in an appropriate innate and adaptive immune response [34].
The BNIP have been associated with pro-apoptotic function and the dimeric mitochondrial protein encoded by this gene is known to induce apoptosis, even in the presence of BCL2 [35]. The TNFRSF-1B protein encoded by this gene is a member of the TNF-receptor superfamily. This protein and TNF-receptor 1 form a heterocomplex that mediates the recruitment of two anti-apoptotic proteins, c-IAP1 and c-IAP2, which possess E3 ubiquitin ligase activity. c-IAP1 is thought to potentiate TNF-induced apoptosis by the ubiquitination and degradation of TNF-receptor-associated factor 2, which mediates anti-apoptotic signals [36]. Knockout studies in mice also suggest a role of this protein in protecting neurons from apoptosis by stimulating anti-oxidative pathways. The TNFRSF25 receptor has been shown to stimulate NF-kappa B activity and regulate cell apoptosis [37]. The signal transduction of this receptor is mediated by various death domain containing adaptor proteins. Knockout studies in mice suggests a role of this gene in the removal of self-reactive T cells in the thymus and it is thought to be involved in controlling lymphocyte proliferation induced by T-cell activation.
The TNF-receptor super-family receptors have been shown to stimulate NF-kappa B activity and regulate cell apoptosis [38]. The signal transduction of this receptor is mediated by various death domain containing adaptor proteins.
It seems likely that the anti-tumor complex CDBPA affects the gene expression depending on the tumor cell type. Previous study in human breast cancer cells indicated that it is exposed to. BP-C1 activates genes that contain guanine-adenine (GA) repeats in their promotor region. Therefore, it was hypothesized that BP-C1 acts as GA-binding proteins (GABP) which are ets transcription factors that control gene expression in several important biological settings [12]. Thus, the intrinsic as well as extrinsic apoptotic pathways were stimulated by CDBPA both in breast cancer cells and as shown in this study in NHL patients. However, results of the present study indicated that different pathway may be involved in induction of apoptosis in human BL cells, various apoptotic genes were activated [12]. Platin has been used in several efficient 2nd line schemes such as RICE studied in pediatric patients [11] and in adults [39]. The new substance CDBPA has been developed for treatment of solid as well as hematological tumors due to its strong immune-modulatory effects besides its cytostatic effects which has been shown to reduce the occurrence of adverse events and decreasing the toxicity of the cancer disease itself, thereby improving the quality of life especially in stage IV breast cancer patients [13-16]. Thus the authors advocate that CDBPA due to its immune-modulatory effects should be used in 2nd line therapies instead the current used platin drugs to spare the patients for adverse effects, toxicity and to improve the quality of life for these patients. In our opinion CDBPA should be considered as a complete new anti-cancer substance and not as a cisplatin analog. The novelty of the CDPBA treatment modality lies not only in its antitumor efficacy with very low number of adverse events, but also in a cell target switch, now aiming in the present study at tumor lymphoblastic cells. The observed adverse effects are minor in patients treated with CDBP as compared to conventional chemotherapeutic drugs [40-42].
CDBPA acts on both; the extrinsic and the intrinsic apoptotic pathways of BL cells without causing cell cycle arrest but only reduction of the various phases. Moreover, the BL in vivo model has demonstrated good efficacy of CDBPA treatment in NOD-scid mice with a significantly improved therapeutic window as compared to the current standard of care R-CHOP.
The authors wish also to express their gratitude to Pipe Line Biotech A/S, Århus Denmark for the conduction of the present in vivo experiments and for an exemplary collaboration.