Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Research Article - (2015) Volume 3, Issue 3
Lipids formed in seeds of oilseed plant are stored in specific cytosolic structures called oil bodies, which are covered with oleosin proteins. Disruption of oleosin layer by heat treatment makes lipids susceptible to oxidation. By analyzing the amino acid sequence of Arabidopsis oleosin, unique domains were identified within the aminoterminus of the protein. Removal of these domains affected both the size of oil bodies and their susceptibility to lipid-specific stain, Nile Blue A. Amino-terminally truncated oleosin mutant, which was in fusion to green fluorescent protein (GFP), confined to rough endoplasmic reticulum (rER) suggesting that the amino-terminus of oleosin is required for the efficient lateral transportation of oleosin proteins from rER to oil bodies. The size of oil bodies covered by amino-terminal mutants was at least 5 times larger than native oil bodies. Based on the observations, a topological model was generated, which could explain how oleosin maintains and protects the integrity of oil bodies during seed storage.
Keywords: Oleosin; Green fluorescent protein; Oil body; Localisation
Accumulation and storage of lipids in seeds of oilseed plants is essential not only for survival, but also in pollen development and subsequent growth and maturation of plantlets; they also provide an important food source for animals and humans [1,2]. Lipids are stored in subcellular structures called oil bodies, which are also known as oleosomes or spherosomes [3,4]. It has been shown in vitro that a group of oil body-associated proteins, oleosins, define the oil body structure; oil bodies with a diameter smaller than 2 μm contain relatively more oleosin molecules than those with a diameter larger than 2 μm [5,6]. In developing oilseeds, oleosin is localized on the surface of the oil body, which is the main site of triacylglycerol storage [7]. There is evidence to suggest that oleosin is co-translationally integrated into rough endoplasmic reticulum (rER) membranes [8,9]. It is also known from biochemical experiments that biogenesis of oil bodies parallels the localisation of oleosins in rER membranes [10,11]. Even though signal sequence for targeting of oleosin to the sites where oil bodies are formed is not known [8,10,12,13] These data indicate that there is dynamic interaction between sites of oil body formation and oleosin synthesis [14,15]. Oleosins are unusual proteins because they possess three different functional domains: a 70–80-residue uninterrupted hydrophobic domain is flanked by relatively polar amino- and carboxy-terminal domains. Hydrophobic central domain is the key element for localisation of oleosin into oil bodies because lack of it totally abolishes localisation of oleosin into oil bodies [16]. Interestingly, hydrophobic central domain can replace the function of core protein of Hepatitis virus C in binding to lipid membranes indicating its vital but non-specific role in the events of membrane association [17]. Hydrophobic central domain, consisting 68-74 amino acids, is flanked by amino- and carboxyterminal domains, which are more variable in length with different plant species (50-70 and 55-98 amino acids, respectively). Using biochemical experiments, it has been demonstrated that short amino-terminal deletions (20 [N20] and 40 [N40] amino acids; respectively) do not disturb the transportation of oleosin into oil bodies, but longer amino-terminal deletions (N66 and N90) reduce the localisation or fully abolish it [11]. The carboxy-terminal part of oleosin is apparently not needed for localisation of oleosin into oil bodies [18]. However, its role in the topology of oleosin-oil body membrane association is not clear. The analysis of oleosin structure itself has been hampered by the membraneous nature of the protein, and structural analyses have mainly focused on the analysis of oleosin fragments produced and purified in bacteria using methods like CD-spectroscopy. Based on these analyses it was deduced that 80% of the β-sheet structure present in the central domain was arranged in parallel to the intermolecular β-sheet structure. The data suggested that interactions between adjacent oleosin proteins would give rise to a stable β-sheet structure that would extend around the surface of the seed oil bodies stabilizing them in emulsion systems [19]. We have demonstrated using GFP-fusions that rough endoplasmic reticulum (rER) is the site of oleosin biosynthesis and that oil bodies are laterally transported within the rER membranes [20]. Here it is presented that oleosins protect the oil bodies by preventing the oxidation and interaction of small molecules with oil bodies. Moreover, it is demonstrated that deletion of the N-terminal part of oleosin prevents efficient lateral transportation of oleosin from rER membranes to oil bodies and affects the size of oil bodies. This paper provides information about the role of oleosin in the maintenance of integrity of oil bodies, and indicates the vital role of oleosin in protection of lipids from external effects during seed storage. Molecular model on putative topology of oleosin at the lipid-water interface, i.e., oleosin-oil body complex is presented in support of our findings.
Plant material and biolistic gene transfer
Nicotiana benthamiana and Brassica rapa subsp. oleifera were grown in a soil in a greenhouse at 24°C and 20°C, respectively. Plants were illuminated using fluorescent lights (Lucalox®, type LU400/ HO/T/40) operated on a 16 hr day photoperiod. Mature, full-expanded, leaves were subjected to particle bombardment as described earlier [20].
Cloning and constructs
Arabidopsis thaliana oleosin gene carrying N-terminal deletion of 66 amino acids was created with PCR from full-length oleosin using primers:
OLE66-forw: 5’-GCACCATGGTTGGAACTGTCATA
OLE66-rev: 5’-GAACTCGAGAAGTAGTGTTGCTG.
PCR product was cloned in-frame with GFP. Full-length oleosin- GFP fusion has been described earlier [20].
Staining of oleosin-GFP mutants localized in oil bodies with neutral lipid stain, Nile Red
Epidermal cells of N. benthamiana expressing oleosin-66-GFP 24 hours after particle bombardment, were infiltrated with an aqueous solution (100 ng ml-1) of lipid-specific stain, Nile Red (Sigma-Aldrich, St. Louis, USA) [21]. Fluorescence from GFP and Nile Red were visualized using appropriate confocal microscope fluorescence filters as described earlier [20].
Analysis of conjugated dienes and volatile oxidation products
Oxidation stability tests were done with intact and heat deactivated oil bodies isolated from Brassica rapa subsp. oleifera seeds as described previously [22]. Heat-deactivation was done at 90°C for 5 minutes under nitrogen atmosphere. Stability test was done in closed test tubes at 37°C. Development of the oxidation was followed three times per week analysing conjugated dienes and volatile oxidation products, such as propanal, which are typical lipid oxidation products used as markers in food analysis [23]. Each measurement was repeated twice. Conjugated dienes were measured spectrofotometrically at 234 nm in methanol. Volatile compounds were measured with headspace gas chromatograph (Autosystem XL, Perkin Elmer, USA).
Computer analysis
The following proteomics softwares were used: GOR1, GOR3, GOR4, ScanProsite, PATTINPROT, Jpred, nnPredict, SOPM, SOPMA, HNN, SSP, NNSSP, and SSPAL. These tools are freely available at http://www.expasy.ch/tools/. The sequence alignment was performed using Multalin version 5.4.1. [24,25].
Heat-treatment makes lipid bodies susceptible to oxidation
To study the role of oleosin in protection of the oil bodies, we isolated oil bodies from Brassica rapa subsp. oleifera seeds, subjected the oil body fraction to oxidation using heat-treatment, and measured the formation of oxidation products by standard assays used in food analytics. The formation of conjugated dienes was observed both in intact and heat-treated lipid bodies during the assay. However, intact oil bodies were more resistant to oxygen than inactivated oil bodies (Figure 1A). Furthermore, the amount of volatile oxidation products also increased more rapidly in heat-treated than in intact lipid body fraction (Figure 1B) suggesting that heating makes oil bodies susceptible to denaturation by oxidation. These results based on biochemical assay give formal proof, although indirectly, that oleosin has a significant role in protection of lipids in oil bodies.
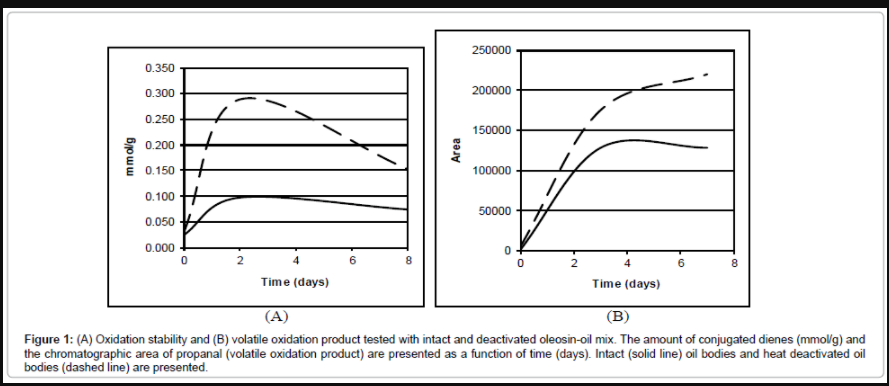
Figure 1: (A) Oxidation stability and (B) volatile oxidation product tested with intact and deactivated oleosin-oil mix. The amount of conjugated dienes (mmol/g) and the chromatographic area of propanal (volatile oxidation product) are presented as a function of time (days). Intact (solid line) oil bodies and heat deactivated oil bodies (dashed line) are presented.
Oleosin structure affects subcellular localization and size of lipid bodies
To further analyze the role of oleosin in topology of oil bodies, localization of native oleosin and its amino-terminally truncated version were analyzed as fusions to green fluorescent protein (GFP). Nicotiana benthamiana plants were subjected to particle bombardment to track transportation of native and amino-terminal oleosin mutant fused to GFP (designated as FL-oleosin-GFP and oleosin-66-GFP, respectively). Native oil bodies with the size less than 2 μm were released from rER and found sporadically distributed in the cytosol when cells were bombarded with FL-oleosin-GFP (Figure 2A). Instead, most oil bodies visualized using oleosin-66-GFP were retained in rER membranes (seen as solid green fluorescence around the region of oil body formation), and only some interacted with oil bodies (Figure 2B-2E). Interestingly, the size of such oil bodies was highly increased compared to native oil bodies (Figure 2B and 2D). Taken together, these observations suggest that the size of oil bodies and release of oleosin from rER in epidermal cells of N. benthamiana is strongly dependent on amino-terminal part of oleosin. To further confirm that oil body structure was affected by truncated oleosin mutants, cells expressing either FL-oleosin- GFP or oleosin-66-GFP transiently (at 24 hr p.i.) were stained with a neutral, lipid-specific stain, Nile Red, and simultaneously visualized by confocal microscopy. Nile Red penetrated (within 5 minutes) into oil bodies covered with truncated oleosin proteins (Figure 2C and 2E). In contrast, no staining of oil bodies covered with mature, full-length oleosin-GFP was found even after 4 hours incubation with the stain (Figure 2A). Thus, removal of the N-terminal part of oleosin makes the oil bodies accessible to hydrophobic molecules, and suggests that N-termini of oleosin proteins are essential for rigid oil body formation and also for the control of oil body size as a result of oleosin-oleosin interactions. These results support the hypothesis that oleosin prevents interaction of lipids with small molecules.
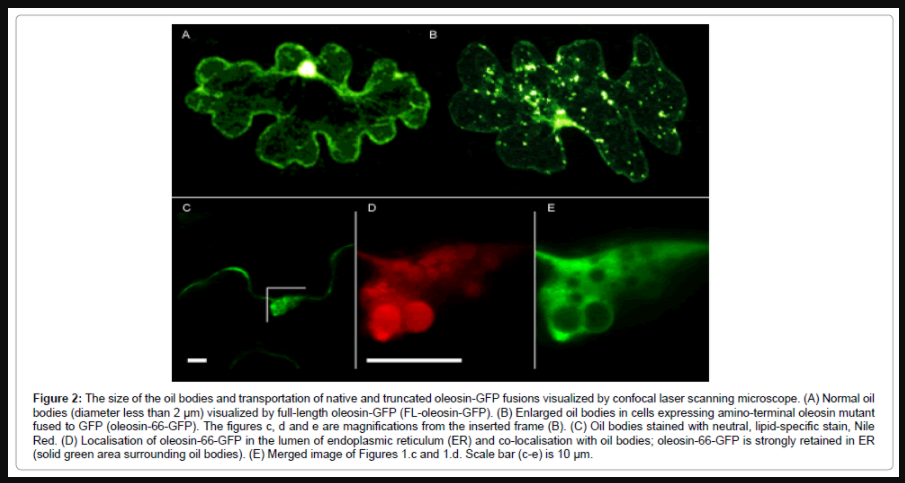
Figure 2: The size of the oil bodies and transportation of native and truncated oleosin-GFP fusions visualized by confocal laser scanning microscope. (A) Normal oil bodies (diameter less than 2 μm) visualized by full-length oleosin-GFP (FL-oleosin-GFP). (B) Enlarged oil bodies in cells expressing amino-terminal oleosin mutant fused to GFP (oleosin-66-GFP). The figures c, d and e are magnifications from the inserted frame (B). (C) Oil bodies stained with neutral, lipid-specific stain, Nile Red. (D) Localisation of oleosin-66-GFP in the lumen of endoplasmic reticulum (ER) and co-localisation with oil bodies; oleosin-66-GFP is strongly retained in ER (solid green area surrounding oil bodies). (E) Merged image of Figures 1.c and 1.d. Scale bar (c-e) is 10 μm.
Sequence analysis of oleosin protein predicts specific domains essential for lipid body structure
To obtain structural support to our biological observations, which suggest that oleosin has a major role in determining size and protection of oil bodies, the structure of oleosin proteins was further analyzed using various protein analysis programs. Previously, the oleosin structure has been analyzed using FT-IR, NMR or CD spectroscopy, and it has been suggested that the hydrophobic domain of oleosin is composed of 80% of the β-sheet structure, which is arranged in parallel to the intermolecular β-sheet structure [19]. However, problems in membrane protein secondary structure determination in aqueous medium or as dry film may reflect incorrect structural properties of proteins, because they are not in natural lipid environment. We analyzed oleosin protein sequences with models predicting protein behaviour in lipid-water emulsions [26]. According to the consensus results from the analysis of amino-terminal portion (residues 1-40) of Arabidopsis oleosin using proteomics tools (http://www.expasy.ch/tools/), the segment composed of residues 1-40 has mainly a random coil structure with smaller stretches of α-helical and β-strand structures. For example, prediction made using GOR1, GOR3, GOR4, Jpred, nnPredict and SOPM methods suggested that the arginine residues of segment 23-28 interact with the phospholipids in the core of the oil body, while the aspartic residues of the same segment participate in the formation of the negatively charged surface of the organelle. The whole segment of Arg-Asp-Arg-Asp-Gln-Tyr-Gln-Met (amino acids 23-30) is β-strandstructured at the lipid:water interface, and therefore essential for coverage of the surface of oil bodies. The results with ScanProsite and PATTINPROT software surprisingly indicated that the pattern Arg- Asp-Arg-Asp-Gln-Tyr (residues 23-28) found in Arabidopsis oleosin appears to be present only in very few protein sequences in SWISSPROT databank: three of these proteins are oleosins (major oleosin nap-II fragment [P29111], oleosin bn-III [P29110], and oleosin bn-V, fragment [P29109]) from Brassica napus, and one is encoded by a plant pathogen, Ralstonia solanacearum [27]. While oleosin bn-V (OLE5) and Arabidopsis oleosin (OLE1), have only one such segment, the oleosin nap-II (OLE2) has two repetitive segments: residues 7-12 and 27-32. Moreover, the oleosin bn-III (OLE3) sequence includes three repetitive segments across equal amino acid interval: residues 19-24, 29-34 and 39-44 (Figure 3). All of these putatively β-strand-structured six residues long segments are located in the amino-terminal domain of the corresponding oleosins suggestive of the highly unique nature and importance of the amino-terminus of oleosins in the maintenance of the oil body structure.

Figure 3: Multiple sequence alignment of oleosins with hierarchical clustering.
It has been suggested that the carboxy-terminal domain of all oleosins contains a potential amphipathic α-helical region of about 15- 20 residues [28]. According to the consensus prediction of the secondary structure (methods GOR1, GOR3, GOR4, nnPredict, SOPM, SOPMA, HNN, SSP, NNSSP, and SSPAL) of the carboxy-terminal domain of Arabidopsis oleosin, two amphipathic α-helical structures can be identified in the segment 120-173 of the primary sequence: 130-Lys- Leu-Asp-Ser-Ala-Arg-Met-Lys-Leu-138 and 141-Lys-Ala-Gln-Asp- Leu-Lys-Asp-Arg-Ala-149. These two α-helices are constructed by the same pattern, which can be written as Lys-h-o-o-h-[Arg/Lys]-x-[Arg/ Lys]-h, where h stands for a hydrophobic residue (Ala, Cys, Phe, Ile, Leu, Met, Val, Trp, Tyr), o for a polar or negatively charged residue (Asp, Glu, Gly, His, Asn, Gln, Ser, and Thr), and x for any residue. Again, the results with ScanProsite and PATTINPROT software indicated that a double pattern, which corresponds to two such α-helices separated by 1-10 residues: Lys-h-o-o-h-[Arg/Lys]-x-[Arg/Lys]-h-x(1,10)-Lysh- o-o-h-[Arg/Lys]-x-[Arg/Lys]-h, exists only in Arabidopsis oleosin (residues 130-149), major oleosin nap-II (residues 132-151), oleosin bn- III (residues 144-163) and oleosin bn-V (residues 132-151). If a helical wheel is drawn using the sequences that putatively form α-helix (amino acids 130-138 and 141-149), the first and eighth positively charged residues of these sequences will be on one side of the wheel. Another side of the wheel (third, fourth and seventh positions of the sequences) is mainly hydrophilic with one or two negatively charged amino acids. Thus, the positively charged residues of two -helices interact with phospholipids on the surface of oil bodies, and the negatively charged residues are oriented towards the exterior of the helix. The secondary structure predictions do not show any content of -helical or β-strand structures from residues 162-167 of oleosin. However, because of the alternation of positively and negatively charged residues, we deduced that this sequence can produce β-stranded conformation on the lipid phase, and its spatial structure may be determined by interactions between oleosins and the core of the oil bodies. Recently, it has been shown that such intermolecular formation of β-sheet is possible in lipid-water emulsions [26], which is in line with our observations. Thus, we propose that amino- and carboxy-termini of oleosin bind intermolecularly to each other forming an antiparallel β-sheet structure (Figure 4A), which is anchored the hydrophobic central domain onto the surface of oil bodies (Figure 4B). By analogy with the experiments obtained with the outer surface protein A [26], conformation of such shell will be highly stable. In conclusion, the simplest structure to explain interactions obtained by computer analyses of amino- and carboxy-termini is a monolayer of antiparallel β-sheets that are formed by the putative β-stranded segments of amino- and carboxy-terminal domains, and a monolayer of parallel α-helical sheets, which are built by the amphipathic α-helices of carboxy-terminal domains (Figure 4). These results and those of expression of plant oleosins in yeast and animal cells [17,29] and the rare existence of oleosin domains strongly support the view that oleosin-like protein structures are unique in their ability to function at water-lipid interface to cover oil bodies.
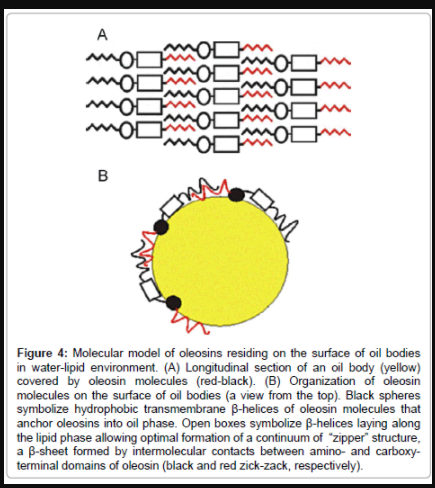
Figure 4: Molecular model of oleosins residing on the surface of oil bodies in water-lipid environment. (A) Longitudinal section of an oil body (yellow) covered by oleosin molecules (red-black). (B) Organization of oleosin molecules on the surface of oil bodies (a view from the top). Black spheres symbolize hydrophobic transmembrane β-helices of oleosin molecules that anchor oleosins into oil phase. Open boxes symbolize β-helices laying along the lipid phase allowing optimal formation of a continuum of “zipper” structure, a β-sheet formed by intermolecular contacts between amino- and carboxyterminal domains of oleosin (black and red zick-zack, respectively).
In this study we imaged the effects of native and truncated oleosins on subcellular localization of lipid bodies and performed extensive motif/domain analyses of oleosin protein. The results allowed us to propose a topological model that could explain how oleosin protects oil bodies e.g., during seed storage. In prior studies we have analyzed the accumulation of oleosin in rER using green fluorescent protein (GFP) fusions and shown the first real-time visualization evidence that oleosin moves laterally within ER membranes and co-localizes with oil bodies [20]. Oleosins have been shown to bind to lipid bodies via its central domain but no specific function has been assigned to amino- or carboxy-termini in oleosin-oil body topology. In addition, the formal proof of the role of oleosin in protection of lipid molecules in lipid bodies has been missing. To study more closely the role of oleosin in protection of the oil bodies, we isolated oil bodies from Brassica rapa subsp. oleifera seeds as described previously [11], and subjected the oil body fraction to oxidation using heat-treatment (Figure 1). The results showed that heat-treatment affects oil body structure because seed oil is readily oxidized (Figure 1A and 1B). As purified oil body fraction is mainly composed of triacylglycerols and protein(s) of which lipid-binding oleosin forms major part [11], it is concluded that heattreatment of oil bodies resulted in, at least partial, denaturation of oleosin layer covering the oil bodies and/or oil body structure resulting in susceptibility of lipids to oxidation.
While we failed to produce and purify intact oleosin in different protein expression systems and thus cannot provide proof using in vitro reconstitution experiments that oleosin is denatured by heat-treatment used in the oxidation experiments, which would explain the oxidation of oil bodies, we were prompted to further elucidate the oxidationprotective properties of oleosin by microscopical techniques and bioinformatics. Localization of native oleosin and its amino-terminally truncated version were analyzed as fusions to green fluorescent protein (GFP). Amino-terminal part of oleosin has previously been shown to be crucial for the localisation of oleosin in the oil bodies using biochemical assays. We were the first to use real-time imaging of oleosin accumulation onto rER membranes, and showed that the wildtype oleosin-GFP fusion was correctly transported and guided from the site of synthesis into the sites of oil body formation in rER membranes [20]. Earlier it was also shown that a truncated oleosin mutant with the deletion of first 66 amino acids (N66) has reduced expression in various plant tissues [11]. However, these results are based on protein analysis and not localisation studies. Therefore, the techniques used in previous studies do not provide solid evidence on oleosin localisation, transport and structure of oil bodies. In contrast to previous studies, we observed that the deleted N-terminal part of oleosin prevents the efficient lateral transportation of the oleosin from rER to oil bodies (Figure 2). Yet, the size of oil bodies was enlarged in cases compared to oil bodies covered with full-length oleosin proteins. Oil bodies covered with truncated oleosin molecules were also stained with Nile Red, whereas oil bodies covered with intact oleosins were not. As Nile Red is a small molecule with molecular size of 730 Da [21], the results imply that surface of oil bodies is covered by a tight net of oleosin molecules, which will not allow small molecules to pass or diffuse through. Thus, intact oleosin structure is required for protection of the lipid phase and preservation of structural integrity of oil bodies. Similarly, it has been shown in stability studies performed with central domain of oleosin and oil bodies, where oil bodies connected to central domain were unstable compared to full-length oleosin covered oil bodies [19]. Thus, amino- and carboxy-terminal domains are needed for full stability of oil bodies.
To obtain structural support to our biological observations, which suggest that oleosin has a major role in determining size and protection of oil bodies, the structure of oleosin proteins was further analyzed using various protein analysis programs (Figure 3). Previously, the oleosin structure has been analyzed using FT-IR, NMR or CD spectroscopy, and it has been suggested that the hydrophobic domain of oleosin is composed of 80% of the β-sheet structure, which is arranged in parallel to the intermolecular β-sheet structure [19]. It has also been suggested that the amino-terminal domain of the oleosin isoform from sunflower reconstituted into liposomes possessed increased β-helical and β-strand structuring compared to the purified and renatured aminoterminal domain [19,22,30]. However, it is evident that spectroscopical methods are severely restricted for the study of membrane protein in the reconstitution conditions, because of non-isotropic motion of membrane proteins in lipid phase [30]. Thus, the present models of oleosin localisation, including all three (amino-, central, and carboxyterminal) domains, are based on careful pH studies, in which charges of oil body and oleosin are calculated together resulting deduced conformational structure of oleosin [28]. Problems in membrane protein secondary structure determination in aqueous medium or as dry film may reflect incorrect structural properties of proteins, because they are not in natural lipid environment. Therefore, we noticed that generation of an alternative model including folding aspects of lipid phase results not only in more plausible explanation for oleosin-lipid interaction, but also in new insight about adjacent oleosin molecules. Similar models have not been created previously, due to lack of programs that could predict protein structure at lipid:water interphase. We used the observations of the behaviour of protein structures in lipid-water emulsions (Lefevre and Subirade 2003) to develop a model of oleosin on oil body surface (Figure 4).
According to the analysis of the amino acid sequence of Arabidopsis oleosin (SWISS-PROT acc. no. P29525) the carboxy-terminal domain of Arabidopsis oleosin contains a mixed-charge hexapeptide Asp-Arg- Asp-Arg-Thr-Arg (DRDRTR; residues 162-167) characterized by the presence of a repetitive alternation of Arg and Asp. This sequence of residues 162-167, which is located after two amphipathic β-helical structures, is very similar to sequence of residues 23-28 of the aminoterminal domain. Except for the major oleosin nap-II (residues 164- 169), oleosin bn-III (residues 184-189) and oleosin bn-V (residues 172- 177), at least two examples of this hexapeptide exists in known proteins. The first one is a protein called synaptotagmin C (P24507), which is a synaptic vesicle protein and a putative trigger of exocytosis in animal systems [31]. The second one is an Arabidopsis homolog of U1 snRNP 70K gene [32], which is involved in alternative splicing of pre-mRNAs in animals. Interestingly, the plant homolog of this protein contains repetitive sequence of Asp-Arg-Asp-Arg-Thr-Arg domain, which is a potential nuclear localization signal. Thus, these data not only imply that domains found in oleosin protein are important for its function particularly in water-lipid environment but they also suggest that they may serve as localization signals. The relevance of this to oleosin function remains to be shown, but these data suggest the presence of putative localization signal in the protein.
To summarize, we have demonstrated that seed oil is oxidized by heat treatment indicative of the protective role of oleosin. The structural features defining the integrity of oil bodies was analyzed creating amino-terminal deletions of oleosin, and it was shown that they define the size of oil bodies. The oleosin sequences were further analyzed to develop a model to explain the phenomenon. We suggest that oleosins form tight shell around the oil body, in which the hydrophobic central domain of oleosins, modelled as an α-helical hairpin [33], anchors the oleosins tightly into oil bodies. By analogy with the experiments obtained with the outer surface protein A [34], conformation of such shell will be highly stable. These results and those of expression of plant oleosins in yeast and animal cells [17,29], and the rare existence of oleosin domains (Figure 3) strongly support the view that oleosin-like protein structures are unique in their ability to function at water-lipid interface to cover oil bodies and probably to facilitate packing of lipids into oil bodies. Giving that oleosins form extremely tight net to cover lipids in oil bodies suggests that their structure may have key importance in preventing chemical alterations of lipids during their storage in seeds [35].
This project was funded by grants from TEKES, Foundation of Finnish Food and Drink Industry, Academy of Finland (no. 48719, 53864 and 54799) and European Union (AIROPico, FP7-PEOPLE-2013-IAPP Grant no. 612308).