Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Research Article - (2016) Volume 5, Issue 5
Panax ginseng is traditionally used for enhancement of physical capacities, especially in the condition of severe fatigue. To evaluate the ability of ginseng in enhancing performance, we focused on the gene expression of AMPK, PGC-1α and SIRT1 genes, involved in energy balance. Gene expression was evaluated in C2C12 myotubes, using P. ginseng C.A. Meyer standardized extract G115, at various concentrations (10-50-100-200 mg/L) for 24, 48, 72 h of incubation. G115 increased significantly the expression of AMPK and PGC-1α in the dose and time-dependent manner: at the concentration of 10 mg/L there is an increase after 72 h of incubation while at 50 mg/L after 48 h and maintained for 72 h. G115 increased expression of SIRT1 at 10 mg/L after 24 h, while at 50 mg/L after 24 and 48 h. The treatment enhanced cellular ATP levels and O2 consumption only at 50 mg/L. We investigated AMPK, PGC-1α and SIRT1 expression in gastrocnemius muscle during acute and prolonged exercise in mice treated with G115. G115 showed no effect on acute exercise compared to untreated mice, but after prolonged exercise, there were significant increases of AMPK and PGC-1α. Expression of SIRT1 was also significant in treated animals’ undergone prolonged exercise. Our data indicate that treatment with G115 increases AMPK and SIRT1 expression that may be related to the induction of PGC-1α expression which improves the energy balance, as demonstrated by increased production of ATP and oxygen consumption. In addition, G115 is able to counteract the inhibition of gene expression of AMPK and PGC-1α due to the prolonged exercise.
<Keywords: Panax ginseng; Energy; Gene expression; G115
Panax ginseng (P. ginseng C.A. Meyer, Araliaceae) has been used in oriental medicine for over 2000 years having a long history as a general tonic promoting health and it is believed to be a panacea and to promote longevity [1]. The bioactive compounds of P. ginseng consist mainly of ginsenosides, polysaccharides, phenolics and polyacetylenes [2]. It is generally believed that ginsenosides and their metabolites are the major active principles of P. ginseng. These are saponins derived from the triterpenedammarane structure and are of two different types of derivatives of two major aglycones: protopanaxatriol and protopanaxadiol.
All ginsenosides have been reported to exhibit various biological activities such as anti-cancer, anti-diabetic, anti-obesity, neuroprotective, radioprotective, antiamnestic and antiageing effects [2]. Extensive scientific research has documented and reviewed the useful effects of P. ginseng C.A. Meyer. However, as the ginsenosides content of ginseng extracts can vary depending on a number of factors including the species, the age and part of the plant used, and the time of year of harvest, there is a high variability in composition of products in the market; this variability has affected the results of the studies, often confounding a clear interpretation of data generated with different extracts. In addition, the use of non-standardized products leads to a poor reproducibility of results, and lack of batch-to-batch uniformity. To address these issues, the compound used in our study was the G115 extract (Ginsana Products, Lugano, Switzerland), standardized to an invariable 4% ginsenosides content. This extract is produced in Switzerland under the control of Swissmedic, the Swiss agency for the authorisation and market supervision of all therapeutic products, and constantly controlled for appropriate standardization and microbiological quality, as well as for the possible presence of impurities. After the optimal standardization of G115 was achieved, in 1984 Dr. Soldati and Prof. Tanaka documented a relationship between the age of the plant and the content of ginsenosides [3,4]. Since then, only roots harvested after 5 years of growth are used for the extract. In the last 30 years, extensive analytical, preclinical and clinical research has been carried out to study the quality, efficacy and safety of G115, which has consequently attracted widespread research interest, and made it possible to generate reproducible results. Several clinical studies have been conducted with this extract, particularly in the area of performance enhancement [5-9]. In our previous study, we developed a murine model of stressful exercise in which mice were subjected to chronic swimming exercise, in order to examine the effects of ginseng extract on immune function, focusing in particular on Toll like receptor gene expression after G115 extract administration [10]. It was therefore deemed relevant to further investigate G115 activity and mechanism of action in respect of energy production. Ginseng is considered as an adaptogen, which is a substance assisting normalization of body system functions altered by stress. Studies with animals show that ginseng, or its active components, may prolong survival to physical or chemical stress [4]. Then, since exercise is considered a form of stress, people subject to exercise frequently make use of adaptogens. The ability of ginseng to improve an individual's strength, endurance and speed of recovery has been investigated in many studies [11-13], however, with some criticisms [14]. Various methodological problems such as inadequate sample size and lack of double-blind control and placebo paradigms may have contributed to conflicting results. Enhancement of physical work capacity was selectively observed in studies with larger subjects’ number, where ginseng was taken with sufficient dosage (200–400 g/day of standardized P. ginseng root extracts containing 4% ginsenosides) and for not less than 8 weeks; age and physical condition of the subjects seems also relevant for the identification of ergogenic effects [15]. Ginseng has been also traditionally used to enhance wellbeing, energy and recovery of physical strength and the potential for recovery is corroborated by studies with animal models of severe fatigue [16,17]. Anti-mental fatigue effects of ginseng extract G115 have been reported in clinical trials on healthy volunteers [18]. Although to date, there are not many information about the anti-fatigue effects of ginseng polysaccharide, they seem to play a relevant role for these therapeutic properties possibly related to prevention of oxidative stress [19]. In animal studies the G115 extract exerted a dose dependent inhibition of the formation of free radicals in rats subjected to physical stress exercise on a treadmill [20]. A clinical study [21] suggests that the beneficial effect of 4 weeks ginseng administration observed in patients with idiopathic chronic fatigue (ICF) may be partly attributed to its antioxidant properties, and oxidative stress is considered as a main contributor to the pathology of chronic fatigue [22]. Moreover, animal studies with the extract G115 show enhancement of glucose transportation in the cells [23], and its pre-treatment increased the production of the Toll like receptor 4 in mice during physical stress [10].
While each individual study mentioned here cannot on its own be taken as a proof of efficacy, these investigations, taken together provide, albeit weak, a support for a beneficial effect of ginseng in these indications.
To evaluate the possible mechanism of action of ginseng in enhancing performance, we focused our attention on the expression of AMPK, PGC-1α and SIRT1 genes, which are involved in the energy balance, acting as a coordinated system that controls energy intake.
AMPK activation plays a relevant role in the transcriptional adaptation to physiological condition of energy expenditure and in the maintenance of intracellular energetic equilibrium under ATP-depleting energetic stresses like exercise, [24]. AMPK regulates metabolic enzymes, increasing the expression of those genes related to mitochondrial respiration, glucose transport and glycolysis [25,26].
Furthermore, AMPK is a modulator of the transcription factor coactivator peroxisome proliferator-activated receptor γ coactivator- 1α (PGC-1α) [27]. PGC-1α is a critical regulator of mitochondrial biogenesis, cellular respiration and energy substrate utilization in skeletal muscle, under acute contraction [28,29] or exercise training [30], by activation of transcription factors modulating a transcriptional pathway that regulates energy homeostasis [31].
PGC-1α activity is also regulated by Silent Information Regulator T1 (SIRT1) that directly interacts and deacetylates PGC-1α. SIRT1-mediated regulation of PGC-1α activity has a major role in the metabolic adaptations to energy metabolism in different tissues [32]. A number of articles were published to demonstrate that the pharmacological or physiological activities of ginsenosides are associated with AMPK [33-39].
In this study, we tested the hypothesis that P. ginseng extract G115 influences the production of energy through the activation of AMPK. Since SIRT1 can also directly interact with PGC-1α [40,41], additionally we evaluated the effect of P. ginseng on the expression of SIRT1.
P. ginseng C.A. Meyer (G115) powder was provided by Ginsana SA. (Bioggio-Switzerland). This root extract has been standardized at 4% of ginsenosides and characterized by HPLC (Figure 1).
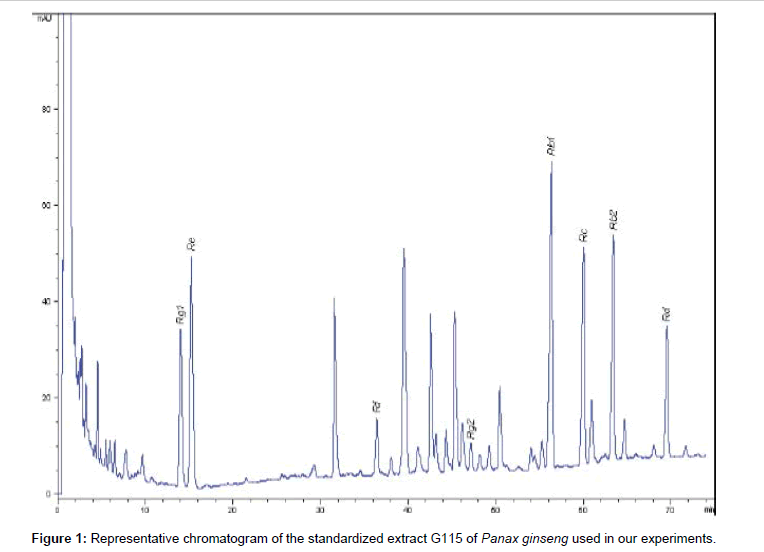
Figure 1: Representative chromatogram of the standardized extract G115 of Panax ginseng used in our experiments.
Ginseng preparation
Since ginsenosides are activated by deglycosylation by colonic bacteria in large intestine, before the transit to the circulation [42], the in vitro experiments were performed after degradation and bioconversion of G115 at acidic (gastric) conditions and in the presence of intestinal microbiota. 100 mg of G115 was incubated with 10 mL of an artificial gastric fluid (37 mM NaCl, 0.03 M HCl, 3.2 mg/mL pepsin, pH 1.6) at 37°C for 2 h. After incubation, the acidity was neutralized by adding 0.5 mL of 2.2 M NaOH. Then, 10 mL of an artificial intestinal fluid (30 mM K2HPO4, 160 mM NaH2PO4, 20 mg/mL pancreatin) were added, and the pH was adjusted to 7.4 using 0.6 M NaOH and 0.2 M HCl. The mixture was shaken in a 37°C incubator for an additional 2 h, centrifuged at 3000 × g for 30 min at 4°C and filtered. Five millilitres of the filtrate were applied to a Sep-Pak® Vac C18 column (Waters) to remove the polar compounds, such as proteins and ions. The loading sample was washed with 30 mL of water and then eluted with 15 mL of methanol. The methanol was removed, and the residue was dissolved in water and then freeze-dried.
Cell culture
C2C12 mouse skeletal muscle cell line was maintained in DMEM (Euroclone, Pero, Italy) with 10% calf serum. Differentiation was induced at 80-90% confluence by switching the cell media with DMEM with 2% horse serum (Euroclone, Pero, Italy). Experimental treatments were started after 96 h by which time the myoblasts were differentiated to form myotubes.
Digested G115 was added to the medium at concentration of 10- 50-100-200 mg/L for 24, 48 and 72 h. After this time, to minimize the possible effects of nutrient deprivation on the cells, they will be washed once in drug-free, fresh culture medium and equilibrated in it for an additional 3 h at 37°C. Cells were then collected and washed twice in PBS.
For the O2-consumption, the cells were suspended (107 cells per mL) in a buffer containing 118 mM NaCl, 4.8 mM KCl, 1.2 mM KH2PO4, 1.2 mM MgSO4, 1 mM CaCl2, 10 mM glucose, and 25 mM Hepes (pH 7.2). For the remaining experimental procedures, the cells were sedimented by centrifugation and kept at 80°C until use.
Animals
Male 6-weeks-old BALB/c pathogen-free mice (Charles River Laboratories, Calco, Italy) were housed in the Pathogen-Free Facility. Animals were treated in accordance with European Community Guidelines and the protocol was approved by the Institutional Animal Care and Use Committee of the Ministry of Healthy- Italy; mice were sacrificed under anaesthesia with urethane (1.6 g/kg).
Mice were subjected to exercise, using a swimming model in which were kept swimming daily for 60 min in a plastic tank filled with water at a temperature of 25( ± 2)°C.
Animals (10 animals per group) were divided in: control group, control ginseng group, swimming group and swimming ginseng group. Ginseng groups were treated daily with 25 mg kg–1 day–1 of extract G115 by oral gavage. Swimming exercise was performed every day for two weeks. After the first session (acute) or after last session of prolonged exercise, animals were anesthetized and sacrificed; gastrocnemius muscles were isolated and snap frozen in liquid nitrogen for Real time PCR analysis.
Gene expression analysis
RNA was extracted from cells using the Trizol reagent (Invitrogen- Life Technologies, Carlsbad, CA) and treated with Dnase (DnafreeTM, Ambion Inc.- Life Technologies, Carlsbad, CA) to avoid falsepositive results due to amplification of contaminating genomic DNA. Concentrations of RNA were determined by the absorbance value of the sample at 260 nm. First-strand cDNA was synthesized from 1 μg total RNA, using an High-capacity cDNA Archive Kit (Applied Biosystems, Forster City, CA) and then analyzed by Real Time PCR, using Stepone Plus Real Time Systems (Applied Biosystems, Forster City, CA). TaqMan PCR was performed using an TaqMan® Universal PCR Master Mix (Applied Biosystems, Forster City, CA); TaqMan probe/primers specific for 18S (code number: Hs99999901_s1), SIRT1 (code number: Mm00490758_m1), PGC-1α (code number: Mm01208835_m1) and AMPK (code number: Mm01296700_m1) were purchased from Taqman® Assays-on-Demand™ Gene Expression Products (Applied Biosystems, Forster City, CA).
All data were normalized to 18S expression and quantification was performed using the ΔΔ CT method. The amount of target, normalized to an endogenous reference and relative to a calibrator, is given by 2-ΔΔCT.
Measurement of O2 consumption.
1-mL cell or tissue sample was incubated at 37°C in a gas-tight vessel that is equipped with a Clark-type O2 electrode (Rank Brothers, Bottisham, U.K.) connected to a chart recorder. Cellular O2 consumption was measured as described [80]. Protein content in both cell and tissue samples was determined by the bicinchoninic acid protein assay.
ATP measurements in cultured cells
The ATP content in myotubes was determined in 2.5% perchloric acid extracts neutralized with K2CO3, by reversed-phase HPLC. Lactate will be measured in the culture medium after deproteinization by using Ultra-4 centrifugal filter devices (cut-off, 10,000; Amicon) as well as in cells after extraction in cold perchloric acid.
ATP measurements in gastrocnemius muscle tissue
Frozen muscles were treated with perchloric acid and neutralized with K2CO3 on ice, and then levels were assayed by HPLC. ATP was quantified based on peak area compared with a standard curve and normalized to frozen tissue weight.
Statistical analysis
Data were analyzed using GraphPad Prism statistical software (GraphPad Software Inc., San Diego, CA). Data were analyzed using the univariate analysis of variance (One-Way ANOVA), followed by Bonferroni’s t-test for multiple comparisons. Statistical significance was accepted when P<0.05. All data are reported as mean ± standard error of the mean.
Gene expression of AMPK and PGC-1α and SIRT-1, in the C2C12 myotubes
To determine whether G115 affects gene expression in muscle cells, C2C12 myotubes were incubated in absence (control) or in presence of G115 at 10-50-100-200 mg/L for 24, 48 and 72 h.
G115, at concentration of 10 mg/L, significantly increased AMPK gene expression in myotubes after 72 h of incubation (p<0.001 vs controls). At concentration of 50 mg/L G115 significantly enhanced gene expression of AMPK at 24, 48 and 72 h (p<0.05, p<0.001 and p<0.001 vs controls, respectively) (Figure 2A). At concentration of 100 and 200 mg/L there was no observed effect: while at these concentrations not clear toxicity showed, the cells did not grow and showed morphological signs of apoptosis, suggesting a possible limiting toxicity at very high concentrations. We next examined the effect of G115 on gene expression of PGC-1α in myotubes. Unlike AMPK, G115 significantly increases the gene expression of PGC-1α only at a concentration of 50 mg/L after 48 and 72 h of incubation (Figure 2B). Also in this case it was not observed any effect at concentrations of 100 and 200 mg/L.
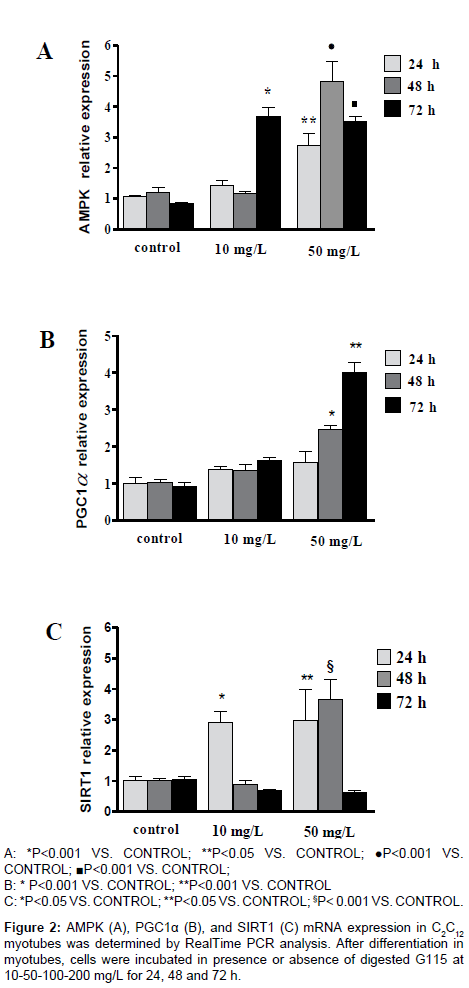
Figure 2: AMPK (A), PGC1α (B), and SIRT1 (C) mRNA expression in C2C12 myotubes was determined by RealTime PCR analysis. After differentiation in myotubes, cells were incubated in presence or absence of digested G115 at 10-50-100-200 mg/L for 24, 48 and 72 h.
Since the expression and activity of muscle PGC-1α is increased by the activation of SIRT1, we examined whether G115 may increase the gene expression of SIRT1. G115 significantly increased the expression of SIRT1 earlier. An increase was observed, at the concentration of 10 mg/L, after 24 h only, while at 50 mg/L concentration there was an increase after 24 and 48 h, returning to basal value after 72 h of incubation (Figure 2C).
Measurements of ATP production and O2 consumption
To examine whether the AMPK and PGC-1α gene expression was associated with an increased respiration and ATP synthesis, we measured the O2 consumption and the ATP production.
The treatment with G115 increased significantly cellular ATP levels (Figure 3) and O2 consumption (Figure 4) at the concentration of 50 mg/L only.
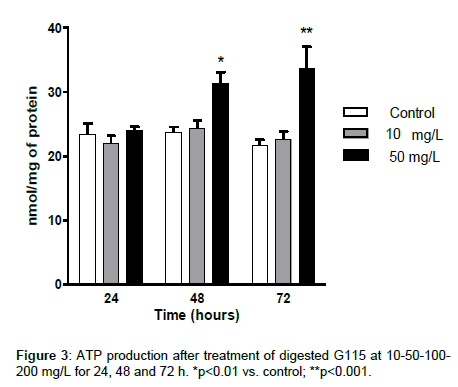
Figure 3: ATP production after treatment of digested G115 at 10-50-100- 200 mg/L for 24, 48 and 72 h. *p<0.01 vs. control; **p<0.001.
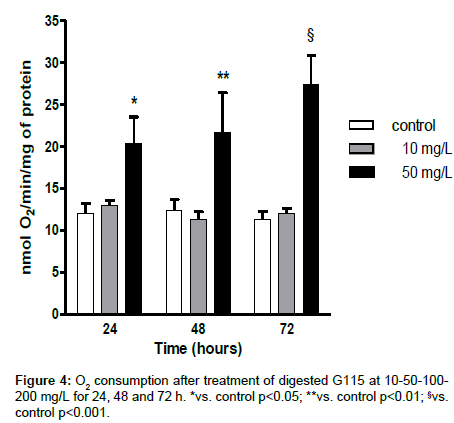
Figure 4: O2 consumption after treatment of digested G115 at 10-50-100- 200 mg/L for 24, 48 and 72 h. *vs. control p<0.05; **vs. control p<0.01; §vs. control p<0.001.
In vivo studies
Previous studies have indicated that P. ginseng may increase physical potency, especially in patients having medical conditions producing severe fatigue. Therefore, we have investigated whether AMPK, PGC-1α and SIRT1 are expressed in gastrocnemius muscle during acute and chronic physical stress in mice treated or untreated with G115. The gene expression of AMPK and PGC-1α shows the same behaviour. After acute stress exercise there is an increase of gene expression (p<0.001) while after chronic stress exercise there is a decrease of gene expression of both AMPK and PGC-1α (p<0.005 and p<0.01, respectively). The treatment with G115 shows no effect on acute stress exercise compared to stressed untreated mice, but after chronic stress exercise there is a significant increase of both AMPK and PGC-1α compared to both controls and chronic stressed mice (p<0.001 and p<0.01, respectively) (Figures 5A and B). The expression of SIRT1 shows a different pattern. A significant increase was observed in the gastrocnemius of treated animals’ undergone chronic exercises (Figure 5C). The O2 consumption and the ATP production measured in the gastrocnemius of mice have shown the same pattern observed for AMPK and PGC-1α (Figure 6).
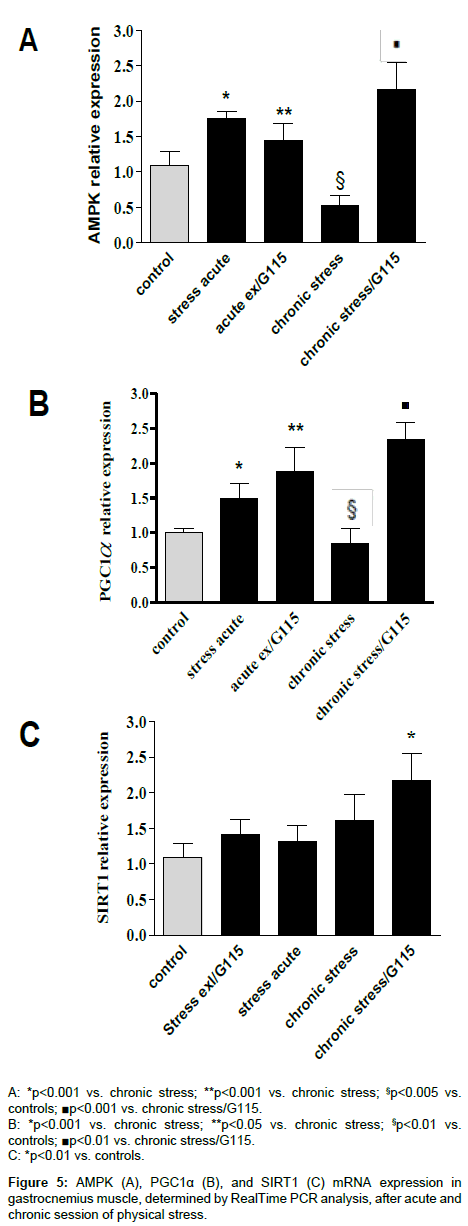
Figure 5: AMPK (A), PGC1α (B), and SIRT1 (C) mRNA expression in gastrocnemius muscle, determined by RealTime PCR analysis, after acute and chronic session of physical stress.
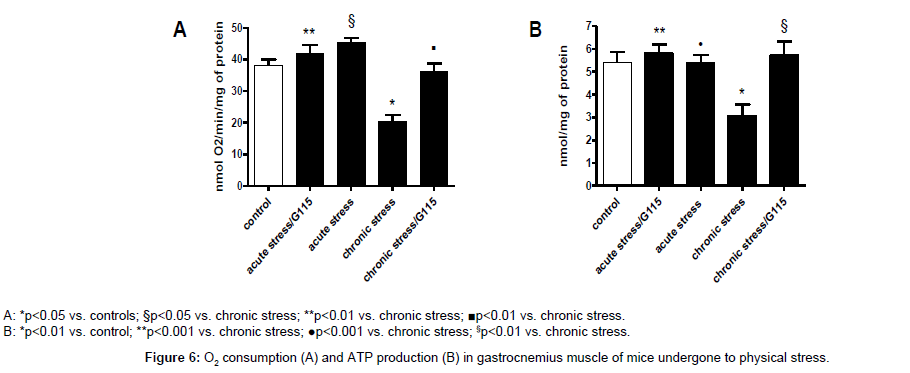
Figure 6: O2 consumption (A) and ATP production (B) in gastrocnemius muscle of mice undergone to physical stress.
Many investigators have found that ginseng may increase the duration of exercise to exhaustion during forced exercise trials [14,21]. In absence of experimental data, this was believed to be due to a general adaptogenic activity of ginseng supplementation [43]. P. ginseng has been investigated extensively for its stress-attenuating activity [44] and it is believed that this adaptation to stress may increase exercise time to exhaustion [45,46].
In untrained adults, it has been reported that consumption of 1,350 mg ginseng per day for 30 days improved their endurance time by more than 7 min and lowered their maximal mean blood pressure and maximal oxygen consumption (VO2max) at the 24th minute during endurance cycling exercise [47]. In another placebo controlled cross-over study [48], in which the subjects were administered with P. ginseng, Siberian ginseng or placebo supplements (1 g/day for 6 weeks for each supplement), it was reported that P. ginseng significantly increased maximal oxygen consumption, post-exercise recovery and pectoral and quadriceps strength. Furthermore, it has been shown that a single dose of 200 mg extract of P. ginseng standardized at 4% of ginsenosides, can modulate circulating blood glucose level, enhance cognitive performance on a mental arithmetic task and ameliorate the increase in subjective feeling of mental fatigue during sustained intense cognitive processing [49].
The results of many studies using ginseng in animal models showed improvements in exercise performance, but the use of large doses or parenteral administration (bioconversion of ginsenosides is known to occur in stomach acid and gut microbial actions before uptake) and the various methods of preparation weakens extrapolation of these data to humans [14].
In our study we used a P. ginseng standardized extract from ginseng roots that are harvested after 5 years of growth, containing invariably 4% ginsenosides (G115; Ginsana Products, Lugano, Switzerland). This extract is well characterised and supported by robust published data, obtained through rigorous quality steps starting from good agricultural and collection practices (GACP). In in vitro studies, the extract was previously digested in order to obtain active ingredients while in in vivo studies the extract was administered by oral route. Considering the support, albeit weak, that ginseng studies taken together provide in increasing physical performance, it appears relevant to highlight that our study identified clear indications that this specific extract has an important role in improving energy balance. Our findings provide an interpretation for previous clinical evidences of G115, potentially unlocking an important mechanism of action of ginseng extract.
The effectiveness of P. ginseng in increasing physical performance can be due to its effect on some key determinants of energy metabolism, rather than to a general “adaptogen” effect.
In this study, P. ginseng increased gene expression of AMPK in C2C12 myotubes in a dose and time-dependent manner. At 10 mg/L concentration the effect appears significant after 72 h incubation while at 50 mg/L concentrations the increase appears significant after 24 h, maintained for 72 h. Higher concentrations of 100 and 200 mg/L did not show any effect, probably because of cellular toxicity (the viability of cells appeared reduced- data not shown).
The gene expression of PGC-1α shows the same behaviour of AMPK. Previous studies have demonstrated that there is a strong overlap in the genes transcriptionally regulated by AMPK and those by PGC-1α, hence suggesting that PGC-1α might be an important mediator of AMPK-induced gene expression [50,51]. Supporting this hypothesis, we may assume that components of the ginseng extract are able to induce the gene expression of AMPK that leads to increased PGC-1α expression.
In this context SIRT1 could play an important role. Several lines of evidence indicate that SIRT1-mediated regulation of PGC-1α activity may play a major role in the metabolic adaptations to energy metabolism in different tissues [52-55].
Our results show, for the first time, that the expression of SIRT1 is early induced by G115 in C2C12 myotubes: either at 10 and 50 mg/L concentration of G115, the gene expression of SIRT1 significantly increased after 24 h of incubation and fell at basal value after 48 and 72 h respectively, indicating that SIRT1 could be an early target of P. ginseng action. It is possible to speculate that the increased expression of SIRT1 and AMPK induced by G115 increases expression of PGC-1α [32].
In any case, the induction by G115 on PGC-1α by AMPK and or SIRT1 may positively control energy production and expenditure.
The action of G115 on the gene expression of SIRT1, AMPK and PGC-1α appears more interesting in in vivo study. In exercise and sports science, ginseng is believed to be a physical performance enhancer; moreover, ginseng has been touted as possessing a stimulant effect and thus improves alertness and decreases fatigue and stress [15,43]. Nevertheless, it has been reported that its benefits were best seen in individuals in poor physical condition [14].
Based on these observations, we evaluated the effect of G115 on gene expression of AMPK, SIRT1 and PGC-1α in the gastrocnemius muscle of mice subjected to intensive acute exercise or to strenuous chronic physical exercise. Acute exercise significantly increases gene expression of AMPK and PGC-1α as reported by previous studies [56,57].
Treatment with G115 showed no change compared to untreated animals subjected to acute exercise. This result, apparently in contrast with the in vitro data, is agreement with a number of previous papers demonstrating that the requirements that ginseng root extracts must have to increase muscular strength and aerobic work capacity are sufficient daily dose (≥ 2000 mg P. ginseng root powder or an equivalent amount of 200 mg of root extract with standardized ginsenoside content), sufficient duration for effects to develop (≥ 8 weeks), and sufficient intensity of physical activity (especially in untrained or older subjects) [58,59]. Interesting results were observed in mice subjected to chronic strenuous exercise in which the expression of AMPK and PGC-1α was significantly depressed compared to the controls while the chronic treatment with G115 induced a significant increase of expression compared to animals untreated as well as to the controls. This result appears in contrast with previous studies demonstrating that PGC-1α as well as AMPK mRNA content was shown to be up-regulated in rodent skeletal muscle after a single swimming exercise bout [59] as well as in humans even in prolonged exercise [60-63].
The gene expression of SIRT1 shows a different pattern with a little non-significant increase after acute exercise both in treated and untreated animals. After strenuous chronic exercise there is an increase (not statistically significant) rather than a decrease, as observed for AMPK and PGC-1α in untreated animals, while in treated animals there is a significant increase compared to controls but not compared to untreated stressed animals.
There is an evident discrepancy in the different behaviour of AMPK, PGC-1α and SIRT1, that we cannot currently fully address. Clearly, AMPK, SIRT1 and PGC-1α do not function independently or linearly; also in previous study, following chronic exercise (multiple bouts of prolonged exercise) in rats, a negative correlation between SIRT1 and PGC-1α expression was identified [64]. In contrast, other studies have shown an increase in SIRT1 expression with exercise in old [65] or adult [66] rat skeletal muscle. Interestingly, one study [60] showed that SIRT1 expression decreased with age and recovered following a single, acute bout of intense exercise, while, in direct contrast, another [67] examined the same muscle in Wistar rats and found that there were elevated amounts of SIRT1 protein with aging, which were then reduced with chronic exercise training.
However, in humans, both an acute bout of sprint exercise [68], and high intensity training for 2 weeks [69] resulted in elevated SIRT1 protein content.
These different results may be consequent to the type of exercise in terms of strength and duration as well as how much stressful is the exercise. We must also consider that in almost all studies that have investigated the expression/activity of AMPK, PGC-1α and SIRT1 in muscle after exercise, were used physiological models of acute or prolonged exercise training not particularly stressful. Our model of stressful exercise [10] has never been used in other similar studies. Therefore, other factors involved in regulation of gene expression may be involved.
Thus, it is apparent from this diverse set of results that more clear evidence is required to understand the relationship between SIRT1 expression/activity, and exercise using standardized models.
Our data suggest that treatment with ginseng extract G115 results in induction of PGC-1α expression that could be related to the observed increase of AMPK and SIRT1. This translates into an improvement of the energy balance as demonstrated by increased production of ATP and by the increase in oxygen consumption. In addition, G115 is able to counteract the inhibition of gene expression of AMPK and PGC-1α due to the prolonged strenuous exercise. These results further support the consistency and effectiveness of the standardized ginseng extract G115, and help to explain the effects of P. ginseng in improving physical performance. We believe our findings are relevant to the many health conditions where energy balance is compromised such as Chronic Fatigue Syndrome, metabolic diseases and cancer, and deserve further scientific and clinical interest.
The work was supported by funds from the Department of Medical Biotechnology and Translational Medicine.
We wish to confirm that there has been no significant financial support for this work that could have influenced its outcome. Andrea Zangara has been a consultant to Ginsana SA. The other authors declare no conflicts of interest.