Journal of Plant Biochemistry & Physiology
Open Access
ISSN: 2329-9029
ISSN: 2329-9029
Research Article - (2018) Volume 6, Issue 1
The main purpose of this research was to investigate the origin of isophorone related compounds, often called as norisoprenoids, in natural products such as honey. Norisoprenoids are volatile compounds and therefore may be identified and semi-quantified using gas chromatography mass spectrometry. In that sense, the reproducible and non-invasive method of headspace solid phase micro extraction (HS-SPME) coupled to gas chromatography-mass spectrometry (GS/MS), representing a targeted metabolomics procedure, was applied to 163 honey samples. Honeys belonged to four different floral types and were collected from sixteen different regions in Greece. Results showed that total isophorone content (TIC) was greatly affected by honey botanical origin and was positively correlated with moisture content using Pearson’s correlation (r=0.84) (p<0.05).
Keywords: Honey; Botanical origin; Norisoprenoids; Ripening; Moisture; Metabolomics
Terpenoids or isoprenoids, are a large family of compounds that are basically formed from C5-isoprene units [1]. This class of compounds includes carotenoids, tocopherol, phytol, sterols and hormones [2]. Depending on the number of carbon atoms terpenoids or terpenes are classified into monoterpene (C10), sesquiterpene (C15), diterpene (C20), sesterterpene (C25), triterpene (C30), tetraterpene (C40) and polyterpene (>C40) according to the multiple number of isoprenoid units. The class of compounds that originate from degradation of terpenes which have lost part of their carbon(s) are called norterpenes or norisoprenoids [3].
It has been reported that terpenoids may be thousands and likely many more that have not been yet identified [2]. Terpenoids play a vital role in all living organisms in respiration chain electron transport (ubiquinone and menaquinone), as well as in cell wall and membrane biosynthesis and stability (bactoprenol, hopanoids in bacteria and sterols in plants). Plants serve as the one of the major sources of terpenoid diversity. Terpenoids are vital for the growth and survival of photosynthetic organisms, since these compounds are responsible of conversion of light into chemical energy and for organization and function of photosynthetic reaction centers including chlorophylls, bacteriochlorophylls, rhodopsins and carotenoids [2]. Other known functions of plant terpenoids include important roles in stress response or in defense mechanisms by combining with reactive oxygen species [4] and so could be protected against internal oxidative damage [5].
Terpenoids are all made from the same five-carbon building blocks, isopentenyl diphosphate (IDP) and dimethylallyl diphoshate (DMADP). IDP and DMADP originate from already known pathways: The mevalonate (MEV) pathway uses acetyl-CoA as a substrate and converts this in six steps, where mevalonate is one intermediate, to IDP, which can be interconverted to DMADP by an isomerase [6]. This pathway operates in eukaryotes, archaic and some bacteria and believed to be the dominant pathway of the formation of terpenoids in all organisms [2]. In was 1993, when an alternative pathway for the biosynthesis of terpenoids in bacteria was reported, which uses pyruvate and glyceraldehyde-3-phosphate as substrates to form both IDP and DMADP [2]. The new pathway is now commonly referred to as the MEP pathway, for the intermediate methylerythritol-4- phosphate.
Honey, the product of Apis mellifera and Apis dorsata honeybees via the collection of floral nectar or honeydew after the addition of enzymes (mainly invertase and glucose oxidase), is a highly concentrated water solution of two simple sugars, dextrose and levulose, followed by small amounts of other more complex sugars and numerous minor components including minerals, polyphenols, organic acids, proteins, vitamins, volatile compounds, etc.. Among volatile compounds, terpenoids and norisoprenoids vary greatly according to floral origin of honey [7-13].
Targeted metabolomic approaches typically aim to quantify the measured metabolites using internal or external standards. Targeted methods with absolute or semi-quantification have the advantage that measurements from different studies can be compared and brought into excessive discussion [14]. Maximum coverage and elucidation of the metabolites (or metabolome’s) chemical space can be achieved by utilizing, among other techniques (high performance liquid chromatography, nuclear magnetic resonance, liquid chromatography mass spectrometry, etc.) gas chromatography coupled to mass spectrometry.
Based on the aforementioned, the aim of the present study was to investigate whether the content of isophorone related compounds could be affected by botanical origin of honey with respect to its moisture content by using a targeted metabolomic procedure on the basis of headspace solid phase micro extraction coupled to gas chromatography mass spectrometry (HS-SPME/GC-MS).
Honey samples
One hundred and sixty three honey samples (N=163) were included in the present study. Honeys belonged to the four dominant varieties harvested in Greece including thyme (N=54), pine (N=50), fir (N=36) and citrus honeys (N=23) and were collected from 16 different regions. All samples maintained at 4 ± 1°C until HS-SPME-GC/MS analysis.
Determination of moisture content
Moisture content (%) was determined using an ATC (Bellingham +Stanley, UK) refractometer. The refractometer was calibrated, prior to use, with a reference material [extra virgin olive oil served as the reference material (moisture of 27%)]. All honey samples were homogenized at room temperature and were directly deposited on the prism of the refractometer.
Headspace extraction
Headspace isophorone related compounds were extracted from honey, using a divinyl benzene/carboxen/polydimethylsiloxane (DVB/CAR/PDMS) fiber 50/30 μm (Supelco, Bellefonte, PA, USA). The fibre, prior headspace extraction, was conditioned by following the manufacturer’s recommendations. Honey samples of 2 g dissolved in 2 mL of distilled water containing 0.20 g NaCl and 20 μL of internal standard (benzophenone, 100 μg/mL), were placed in 15 mL screw-cap vials equipped with PTFE/silicone septa. Each vial was maintained at 45 °C in a water bath under stirring at 600 rpm for the entire headspace extraction. A magnetic stirrer (cross-shaped PTFE-coated, diameter 10 mm) (Semadeni, Ostermundigen–Bern, Switzerland) was placed inside the vials. The optimum sample preparation conditions are given in previous works [10,13]. The temperature of 45°C was fixed in order to keep at minimum the formation of thermal artefacts like furan derivatives, furfural, etc. that could affect the isolation of isophorone related compounds. To avoid any source of contamination of the fibre used for the collection of isophorones, blank runs were carried out before sample analysis. The fibre was cleaned (blank run) after the analysis of every five honey samples.
GC/MS instrumentation and conditions
The GC unit Agilent 7890 A coupled to the MS detector Agilent 5975, was used to for the analysis of honey samples. A DB-5MS (crosslinked 5% PH ME siloxane) capillary column, having dimensions of 60 m × 320 μm i.d., × 1 μm film thickness, was used, with helium as the carrier gas (purity 99.999%), at 1.5 mL/min flow rate. Analysis conditions were the same as described in relevant works [10,11,13].
Mass spectral data processing-identification of volatile metabolites
The identification of isophorones was achieved using the Wiley 7, NIST 2005 mass spectral library. For the determination of linear retention indices a mixture of n-alkanes (C8-C20) dissolved in nhexane, was employed. The standard mixture was purchased from Supelco (Bellefonte, PA, USA). Calculation of retention time indices was carried out for components eluting between n-octane and neicosane. Benzophenone (Molecular weight=182.2179; Chemical formula: C13H10O) was chosen as an internal standard since it did not naturally occur in any of the identified isophorones. The characteristic fragment ions (m/z) of the internal standard were: 51/77/105/182. Isophorone related compounds having ≥ 90% similarity with the Wiley mass spectral library were tentatively identified using GC-MS spectra. For semi-quantification, isophorone content was expressed as μg/kg based on the ratio: peak area of analyte to peak area of the internal standard [11].
The average values of the isophorone related compounds identified in honey samples were compared using T-test at the confidence level p<0.05. Correlation was obtained using the Pearson’s bivariate correlation (p<0.05). Statistical treatment was accomplished using the statistics software SPSS v.20 for Windows.
Isophorone related compounds content
The most characteristic isophorone related compounds identified in fir, thyme and orange blossom honeys were: alpha-isophorone, betaisophorone, 2-hydroxy-isophorone and 4-oxo-isophorone (Table 1). The higher total average content of isophorones (μg/kg) was recorded for fir (417.83) followed by thyme (105.99) and orange blossom (13.34) honeys (p<0.05) (Figure 1). Figures 2a and 2b represent the typical mass spectra and gas chromatogram of fir and thyme honeys, respectively, indicating the isophorone related compounds that were identified. To great surprise, isophorone related compounds were not identified in pine honey samples. This observation could serve as an additional criterion in honey botanical origin differentiation efforts when using that is specific volatile metabolites [7-13].
| Isophorone related compounds |
RT (min) | RIexp/RIlit | Thyme honey (N=54) | Fir honey (N=36) | Citrus honey (N=23) | Pine honey (N=50) | Dominant fragment ions |
Molecular weight (MW) | Chemical formula (CF) |
|---|---|---|---|---|---|---|---|---|---|
| 3,5,5-trimethyl-2-cyclohexen-1-one (alpha-Isophorone) | 20.49 | 1143/1144 | I | I | I | NI | 82/138 | 138.2069 | C9H14O |
| 3,5,5-trimethyl-3-cyclohexen-1-one (beta-Isophorone) | 20.49 | 1143/1144 | NI | I | NI | NI | 82/138 | 138.2069 | C9H14O |
| 2,6,6-trimethyl-2-cyclohexene-1,4-dione (4-oxo-Isophorone) | 20.84 | 1160/1159 | I | NI | NI | NI | 68/96/152 | 152.1904 | C9H12O2 |
| 2-hydroxy-3,5,5-trimethyl-cyclohex-2-enone (2-hydroxy-Isophorone) | 20.99 | 1168/- | NI | I | NI | NI | 70/98/139/154 | 154.2063 | C9H14O2 |
Table 1: Isophorone related compounds identified in Greek unifloral honeys. Where, N: number of honey samples analyzed in the study, RT: retention time in minutes (min), RIexp: experimental retention indices values calculated using the Kovats equation based on the retention time values of standard alkanes. RIlit: literature cited data of retention time indices, obtained from Wiley NIST 7 MS library, I: identified, NI: not identified. Every value is the average of two independent determinations (n=2).
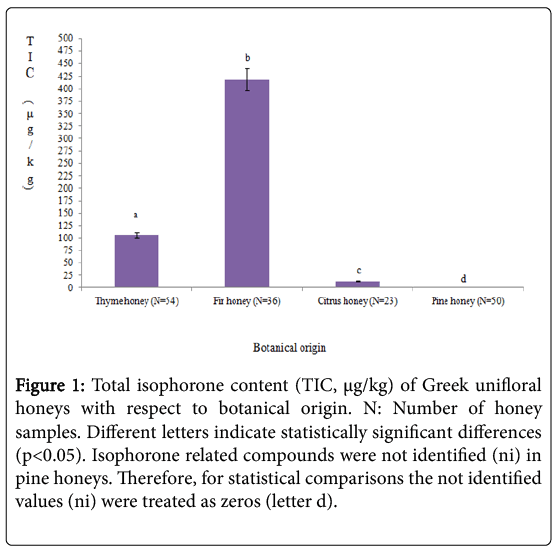
Figure 1: Total isophorone content (TIC, μg/kg) of Greek unifloral honeys with respect to botanical origin. N: Number of honey samples. Different letters indicate statistically significant differences (p<0.05). Isophorone related compounds were not identified (ni) in pine honeys. Therefore, for statistical comparisons the not identified values (ni) were treated as zeros (letter d).
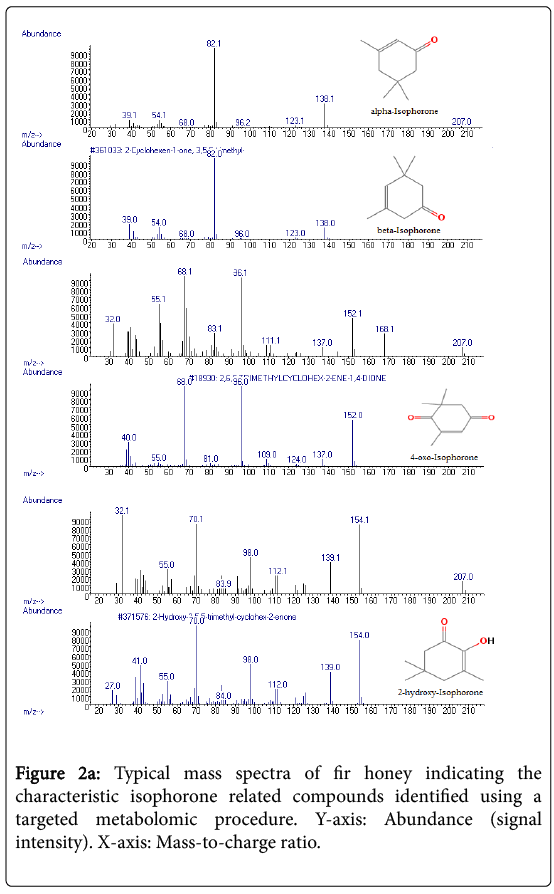
Figure 2a: Typical mass spectra of fir honey indicating the characteristic isophorone related compounds identified using a targeted metabolomic procedure. Y-axis: Abundance (signal intensity). X-axis: Mass-to-charge ratio.
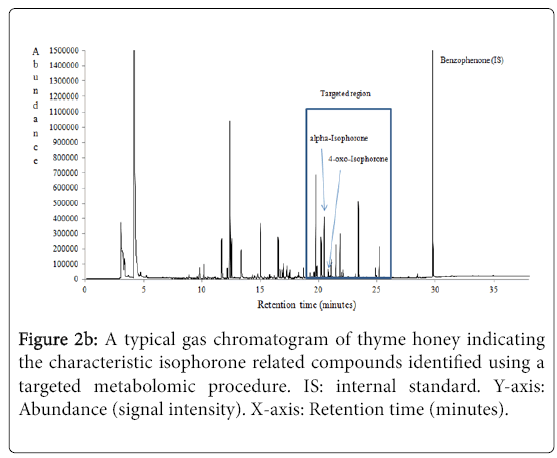
Figure 2b: A typical gas chromatogram of thyme honey indicating the characteristic isophorone related compounds identified using a targeted metabolomic procedure. IS: internal standard. Y-axis: Abundance (signal intensity). X-axis: Retention time (minutes).
Alpha-isophorone recorded a much higher average content value (μg/kg) in fir (1540) compared to thyme (81.27) and orange blossom (53.37) honeys. Beta-isophorone was identified only in fir honey samples. Respective average content value was 21.30 μg/kg. 4-oxo- Isophorone was identified only in thyme honey samples, recording an average content value equal to 236.70 μg/kg. Finally, 2-hydroxyisophorone was identified only in fir honey samples, at an average content value equal to 110 μg/kg. This volatile metabolite could also serve as a principal differentiation marker compound between thyme and fir, pine or orange blossom honeys. Results also showed that the dominant isomer of isophorone in honey samples analysed was found to be isomer I (alpha-isophorone).
Fuente et al. [7] reported that alpha-isophorone recorded higher relative standard concentration of the % total area of the chromatograms obtained using SPME methodology coupled to gas chromatography/mass spectrometry in heather, followed by rosemary, eucalyptus and citrus honeys. Respective findings for 4-oxoisophorone followed the order: heather> rosemary>citrus>eucalyptus honeys. In a previous work dealing with the volatile characteristics of Turkish and Greek pine honeys the authors applied a purge and trap system coupled to GC/MS for the isolation of volatile and semi-volatile compounds. Tananaki et al. [8] reported that the presence of isophorone (% of the total volatile compounds identified) in Greek pine honeys was much lower than that of Turkish pine honeys. Spanish Εucalyptus honeys contained higher amounts (μg/kg) of 4- oxoisophorone compared to thyme honeys or citrus honeys. Respective values were 178, 29.7 and 28.9 μg/kg. The methodology applied included the micro-scale simultaneous distillation-extraction coupled to gas chromatography/mass spectrometry [9]. Sardinian Eucalyptus honeys were also characterized by the presence of 4-oxoisophorone (or 4-ketoisophorone). The relative per cent of 4-oxoisophorone was increased using dynamic headspace extraction instead of solid phase micro extraction, in combination with gas chromatography /mass spectrometry [12]. Therefore, the botanical origin of honey, harvesting year and extraction methods used for the isolation of volatile norisoprenoids may affect the content of isophorone related compounds.
Another important issue to discuss is that, in the present study, there was a positive Pearson’s correlation (correlation coefficient r=0.84) (p<0.05) between TIC and moisture content of honeys. Respective average moisture content values (g/100 g) followed the text sequence: fir (16.99)>thyme (16.51)>pine (15.03)>citrus (14.60). As it can be observed, the higher the moisture content of honey is, the higher the total isophorone related compound content.
A possible theorisis that elucidates the presence of isophorone related compounds in honey
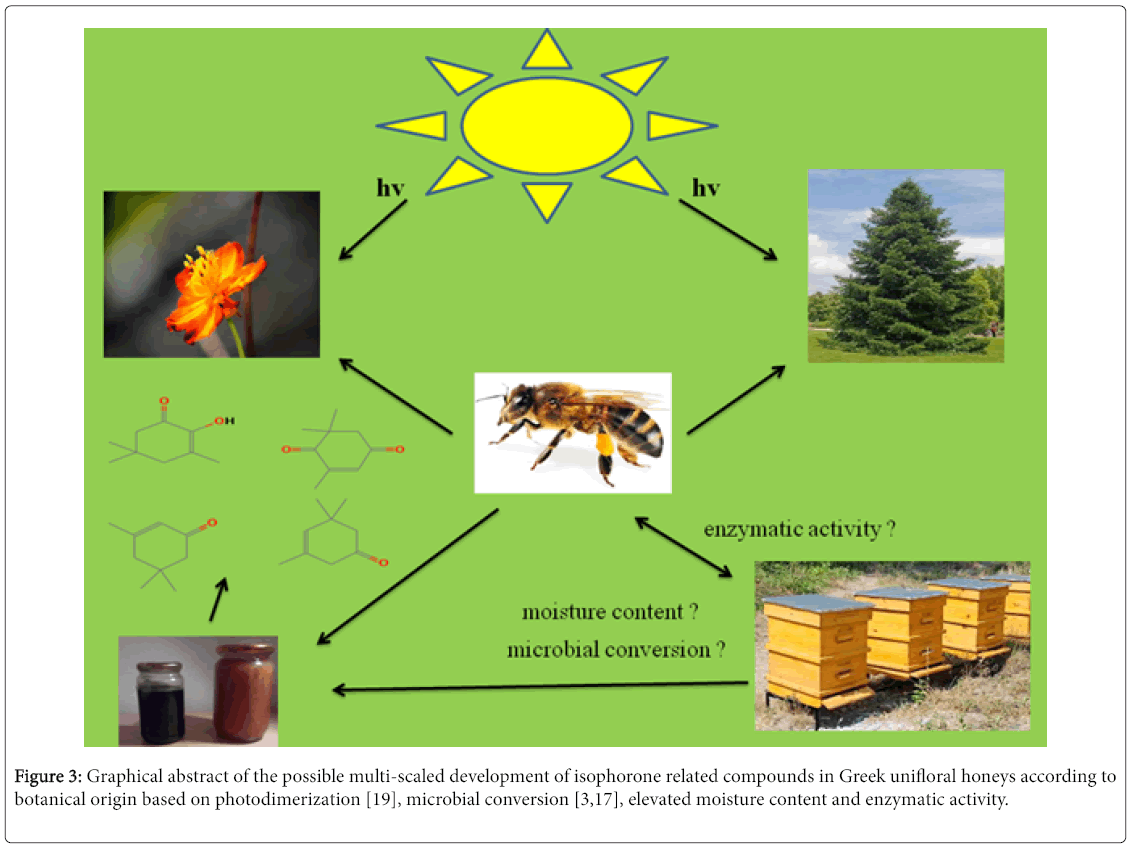
Let’s take a look through a possible theorisis that implies the presence of isophorone related compounds in honey. Nectar is the sugary liquid that contains ca. 80% water, but that is a convenient average that depends on the specific plants and the conditions under which they are growing. Water can be as low as 30% or as high as 92% of the nectar [15]. On the other hand, honeydew is a sugar-rich sticky liquid, secreted by aphids, whiteflies and some scale insects as they feed on plant sap (phloem sap), in order to obtain sufficient nitrogen. Honeydew differs from nectar in containing oligosaccharides synthesized by the insects from the dietary sugars [15].
Previous studies have demonstrated that nectar and pollen contain many microorganisms that can be transmitted to honey [16]. Some secondary sources of the presence of microorganisms in honey are humans, beekeepers equipment, recipients, wind, dust, insects, animal and water. But what type of microorganisms is present in honey? A recent study carried out on honey samples of different botanical and geographical origins showed that the major microorganisms found in honey could be classified into three groups: Bacteria (Bacillus spp., Lactococcus lactis , Aeroccoccus viridans , etc.), Filamentus fungi (Alternaria alternata , Arthrinium spp., Aspergillus niger , etc.) and Yeasts (Zygosaccharomyces mellis, Debaromyces hansenii , etc. [17]. For example, Aspergillus niger JTS 191 proved to be able to convert isophorone. Four products were found and identified as: (4S)-4- hydroxy-3,5,5-trimethyl-2-cyclohexen-1-one, 3,5,5-trimethyl-2- cyclohexene-1,4-dione, 3,5,5-trimethyl-cyclohexane-1,4-dione and 3- hydroxymethyl-5,5-dimethyl-2-cyclohexen-1-one. However, the major products were (4S)-4-hydroxy-3,5,5-trimethyl-2-cyclohexen-1-one and 3-hydroxymethyl-5,5-dimethyl-2-cyclohexen-1-one. The main reactions that took place were oxidation [3].
The same fungus could also reduce 4-oxoisophorone [18]. Sinacori et al. [17] reported the presence of Aspergillus niger in Italian honeydew honeys. Therefore, the microbial conversion of isophorone through the action of that is Aspergillus spp. is probably favoured in honeydew honeys. This is in agreement with present results that showed a higher total isophorone content (TIC, μg/kg) in fir honeys compared to thyme or orange blossom honeys. Another important finding, worthy of discussion, is that TIC was strongly correlated (Pearson’s coefficient r=0.84) with the moisture content (g/100 g) of honeys. In that sense, the increased moisture level in different honey types may affect their total isophorone content or specific isophorone related compounds’ content, since it generates microbial growth.
An additional factor that should be considered (and has been already reported) is the sunlight exposure of flowers or trees in which bees collect nectar or honeydew. An extended sunlight or exposure of flowers and trees to sunlight may result in the development of higher amounts of carotenoids and subsequently increase the level of norisoprenoids (that is isophorones) in plant based products [19]. Therefore, climatic conditions (sunlight, rainfall, humidity, etc.) in a given region may also contribute to the overall development of isophorone related compounds in honey, since apart from the degree of degradation of carotenoids, may also affect the overall production of nectar and honeydew from flowers and trees, respectively.
Finally, the action of invertase in honey which breaks down sucrose to fructose and glucose units could be also taken into account. If we consider that initially isophorone related compounds are bounded to sugar units, rendering them aromatically inactive, then during the action of invertase added by the bees, are released from sugars and turn into active volatile compounds such as that is norisoprenoids. The characteristic odour of isophorone has been reported to be as that of tobacco [3]. Further research upon the role of invertase or other enzymes in the development of isophorone related in honey will validate present theorisis. A possible and multi-optional theorisis that elucidates the presence of isophorone related compounds in honey is shown in Figure 3.
Exploitation of isophorone related compounds
Isophorone is produced on industrial scale by the aldol condensation of acetone using potassium hydroxide (KOH). Diacetone alcohol, mistily oxide, and 3-hydroxy-3,5,5-trimethylcyclohexan-1-one are the intermediates. A side product is beta-isophorone, where the C=C group is not conjugated with the ketone [20]. Before going any further it is of great interest to discuss some valuable information. As shown in the present study the dominant natural isomer of isophorone found in honey is alpha-isophorone followed partially by betaisophorone. In organic synthesis, beta-isophorone is probably favoured. The hydrogenated derivative trimethyl-cyclohexanone is used in production of polycarbonates [20]. Another important application of isophorone is that this compound is easily subjected to hydrocyanation. Hydrocyanation offers the nitrile group and if followed by reductive animation then the final product is isophorone diamine. This diamine is used to produce isophorone diisocyanate which has certain niche applications [20]. Isophorone diisocyanate is also used to produce special polyurethanes.
Since terpenoids cover a wide range of biological functions, have extensive applications in the fields of pharmaceuticals, cosmetics, colorants, disinfectants, fragrances, flavourings and agrochemicals, the exploitation of natural norisoprenoids, as natural breakdown products of carotenoids [21], in pharmaceuticals, biopolymers of food packaging (as antimicrobials), etc. could be of great interest to research community. The LD50 value of isophorone in rats and rabbits by oral exposure is ca. 2.00 g/kg [22].
The presence of isophorone related compounds in honey is a complex phenomenon. Present results showed that it may be affected from moisture content and botanical origin of honey considering, however, the microbial conversion of specific microorganisms [3,17,18]. In addition, factors such as: beekeeping practices, harvesting year, climatic conditions [19], processing practices along with the analytical techniques applied for the determination of such compounds [7-13], should be also considered. Targeted metabolomics analysis in combination with the genetic characterization of honey micro biota may provide in future work useful information upon this research topic.
The author is grateful to Prof. Michael G. Kontominas for the access he provided to the laboratory of Food Chemistry located at the Department of Chemistry in the University Campus of Ioannina, 45110, Greece. Attiki Bee Culturing Co.-Alex. Pittas S.A. (Protomagias str. 9, Kryoneri 14568, Attica, Athens) and local beekeepers from the regions of Messinia, Lakonia, Arkadia, Evia, Aitolokarnania, Arta and Symi, are also greatly acknowledged for the honey samples they donated.