Journal of Horticulture
Open Access
ISSN: 2376-0354
ISSN: 2376-0354
Research Article - (2015) Volume 2, Issue 1
American cranberry (Vaccinium macrocarpon) is naturally adapted to environments with high concentrations of soluble iron. Yet, there is a need to further explore iron nutrition in cranberry given concerns of toxicity problems from irrigation with iron-rich water. This study investigated the threat of iron toxicity by evaluating its effects on shoot growth total shoot weight, length (primary shoot and axillary growth), and number of axillary shoots in cranberry plants exposed to varying levels of a commonly used chelated iron, specifically Sequestrene 330 (ferric diethylenetriamine penta-acetate; Fe-DTPA). Cranberry plants were grown under controlled greenhouse conditions and received varying concentrations of Fe-DTPA in their irrigation water solutions. Four treatments of Fe-DTPA were applied (0, 14 ppm, 28 ppm, 56 ppm Fe), and shoot growth measurements were taken weekly over the course of 20 weeks. At the end of the course, total fresh shoot weight was recorded for each plant and tissues were sampled for nutrient analysis. Overall, shoot length (primary shoot and axillary) was significantly (P<0.05) reduced with increasing Fe-DTPA concentrations and plant symptoms included leaf drop, necrosis, and mortality in the higher concentration treatments. The number of axillary shoots per upright significantly (P<0.05) increased with treatment intensification. Nutrient analysis revealed increasing amounts of iron with increasing Fe-DTPA dosages. However, iron tissue levels were within the normal range found in healthy field plants (142.8-880.7 ppm) in all treatments. We hypothesized that the toxicity symptoms observed in cranberry plants treated with Fe-DTPA are likely not due to iron, but to a specific toxicity of cranberry to the chelators or other unknown components used in Fe-DTPA.
<Keywords: Vaccinium macrocarpon; Iron toxicity; Chelated iron toxicity; Iron chelates; Sodium ferric diethylenetriamine pentaacetate; Cranberry nutrition
The Ericaceous American cranberry (Vacinium macrocarpon Ait.) is an acidophile plant species indigenous to peaty, sandy, acidic soils characteristic of bog environments, which are regularly wetted or waterlogged and possess high organic matter content. These bog environments typically have abundant concentrations of soluble iron and other metals that may be toxic to non-adapted plant species. Although the physiological mechanisms are not fully understood, the family Ericaceae has a reported tolerance to iron [1]. In fact, V. macrocarpon has the capacity to regulate iron supply, uptake and translocation across a wide range of environmental concentrations [2]. Thus, cranberries are also well adapted to low-buffered, sandy environments where mercurial iron species constantly oscillate amid rapidly shifting pH conditions. For example, cranberry cultivation is done in sandy soils, which have been found to be particularly variable in iron concentrations, spatially from region to region, bed to bed, and within beds and also temporally throughout the growing season [3]. V. macrocarpon is also mycoheterotrophic and lacks root hairs, but possess very fine lateral roots termed “hair roots” that help cranberries regulate iron uptake and likely play a role in the successful colonization of cranberry in sandy acidic soils and bog environments [1,4-7].
Indeed, the ability of V. macrocarpon to tolerate high iron concentrations qualifies it as a wetland species [2]. Although cranberry is evolutionarily accustomed to unstable, nutrient-poor, iron-rich conditions, there may be a threshold beyond which iron concentrations may become toxic to the species. Cranberry thrives in an optimal pH range of 4.2 to 5.5, with water being a dominant agent in both natural ecosystems and commercial cranberry production systems, potentially altering pH and iron concentrations [8]. A number of Wisconsin cranberry marshes may be subject to abnormally high iron concentrations, resulting in conditions which might be conducive to iron toxicity risks in the commercial beds. These beds are flooded several times annually and typically overhead irrigated via sprinkler systems. Water drawn for flooding and irrigation sometimes have high concentrations of iron, which may expose both the soil and plants to elevated iron concentrations and deposit “rust” residues on plant vines, flowers, fruit, and litter. Some growers have noticed decreasing yields with an increasing presence of iron on their marshes. Heightened exogenous and/or endogenous iron levels may influence photosynthetic capabilities, discourage pollination, and hinder overall plant growth and performance. The objective of this study was to evaluate the effect of iron fertilizer on total shoot biomass, length (primary shoot and axillary growth), and number of axillary shoots in cranberry plants grown under controlled greenhouse conditions.
Plant material
Cuttings of uprights from cranberry cultivar ‘Stevens’ were collected from two randomly selected beds at a cranberry marsh in Juneau County, WI, in late March 2011. Cuttings were visually sorted according to quality (vigor, incidence of leaves, and uniformity), then trimmed to 9.5-10 cm lengths prior to refrigeration.
Rooting
Selected uprights were rooted using a hydroponic system in controlled conditions at the Walnut Street Greenhouse, University of Wisconsin-Madison. Plastic washtubs covered with aluminum foil were filled with deionized water and tubes connected to fish aquarium pumps were placed within the bins to maintain water oxygen levels. Uprights were suspended in water, having been positioned in holes made in the lids of pink foam insulation board that was approximately 2.5 cm thick. Approximately 2.5-3 cm of the stem base was submerged in the water and levels were replenished as necessary, including a weekly supplement of a slightly adapted Hoagland’s nutrient solution. Plants were subject to a sixteen-hour day under high-pressure sodium lighting at a temperature of approximately 27°C. Clear plastic lids were positioned over the uprights to minimize transpiration and uprights were misted daily. After one week, the plastic lids were elevated 1 cm to allow air circulation and harden off the plants.
Potting
Small pots were prepared for transplanting and these pots were filled with uniform, medium-texture, pH-neutral sand. Uprights were transplanted into small individual pots May 19, seven weeks after being planted in the hydroponic system. A layer of black gravel was spread around the upright in each pot; white gravel was then overlaid in order to algal growth.
Experimental design
Uprights were arranged in a four block randomized complete block design with four treatments and ten uprights per treatment, totaling 160 uprights. Application of treatments began after plants where established in pots and extended for 143 days. Treatments were applied daily and consisted of an iron-containing solution with the following concentrations of iron: 0 ppm (none), 14 ppm (low), 28 ppm (medium), and 56 ppm (high). Each treatment was applied in solution with daily applications of 800-mL deionized irrigation water per potted plant, which was applied directly on the gravel pot surface and varied only in the concentration of iron in solution. Treatment solutions were made from the chelated iron product, Sequestrene 330 (Novartis Crop Protection; Greensboro, IL; ferric diethylenetriamine penta-acetate; Fe-DTPA). In addition to the iron treatments, all plants received the same weekly fertigation using the original hydroponic rooting nutrient solution.
Data collection
To gather natural cranberry iron baseline data, plants were collected from 15 Wisconsin marshes and iron levels were assessed using all fresh vegetative shoot material, which was weighed and submitted for tissue nutrient analysis (AgSource Laboratories, Bonduel, WI). Experimental greenhouse plants were assessed for treatment effects on shoot growth. Shoot growth variables consisted of total shoot biomass, length (primary shoot and axillary growth), and number of axillary shoots. Shoot growth measurements began first week of June and ended the last week of October when overall plant vigor began to decline. Measurements were recorded on twelve dates over a 143-day span. The first nine dates were eight to ten days apart and the last three occurred approximately one month apart. Following plant harvest, all fresh vegetative shoot material was weighed and submitted for tissue nutrient analysis (AgSource Laboratories, Bonduel, WI).
Statistical analysis
Data were analyzed using a repeated measures analysis variance (ANOVA) to determine the main effects of Fe-DTPA treatment on cranberry growth variables. Calculations were performed using the general linear model (mixed) procedure of SAS (ver. 9.2, SAS Institute, Cary, N.C.). The repeated measures model accounted for block, treatment, day and interaction between days and treatments. Where appropriate, means were separated by Tukey’s multiple comparison test between treatments using a value of P<0.05.
Analysis of Variance
An ANOVA with repeated measures was performed to test for effects between treatments Fe-DTPA application on cranberry plant growth variables (Table 1). The best covariance structure for both shoot growth and number of axils was the heterogeneous first-order autoregressive (ARH1), and the best fit model was the linear model, AIC=11103 and AIC=5456, respectively. Tukey’s tests of significance showed significant (P<0.01) differences between treatments for both shoot growth and number of axils (Table 1).
| Treatments | Plant height (cm)90 days |
No. of stems per plant | Fresh weight of plant (g) | Dry weight of plant (g) | Days required for opening of first spike |
|---|---|---|---|---|---|
| T1 | 86.07 | 2.08 | 200.34 | 61.58 | 74.03 |
| T2 | 87.19 | 1.77 | 182.70 | 50.40 | 79.12 |
| T3 | 91.88 | 2.20 | 208.35 | 65.59 | 72.10 |
| T4 | 88.97 | 2.03 | 189.37 | 62.70 | 74.42 |
| T5 | 87.83 | 1.67 | 194.26 | 56.34 | 76.11 |
| T6 | 84.98 | 2.10 | 177.37 | 59.40 | 78.18 |
| T7 | 85.61 | 1.43 | 180.36 | 54.38 | 82.01 |
| T8 | 84.32 | 2.00 | 173.39 | 57.81 | 75.14 |
| T9 | 84.90 | 1.90 | 186.57 | 52.39 | 77.19 |
| T10 | 82.75 | 1.33 | 173.72 | 48.52 | 83.21 |
| T11 | 80.95 | 1.52 | 161.36 | 46.37 | 81.04 |
| T12 | 83.82 | 1.55 | 172.44 | 48.03 | 79.17 |
| S.Em.± | 2.208 | 0.119 | 8.756 | 2.927 | 1.994 |
| C.D. at 5% | 6.48 | 0.35 | 25.68 | 8.58 | 5.85 |
| C.V.% | 7.68 | 11.50 | 8.27 | 9.17 | 8.12 |
Table 1: Effect of organic, inorganic and biofertilizer on days to vegetative parameters in gladiolus cv. Psittacinus hybrid.
Plant growth
The Fe-DTPA concentration gradient visibly impacted cranberry plant shoots. There was a noticeable decrease in plant health in the high and medium Fe-DTPA treatments resulting from diminishing plant vigor and leaf loss. A significant (P<0.05) negative relationship between shoot growth and increasing chelated iron concentrations was evident mid-way through the experiment (day 64) and even more pronounced in the final growth measurement (Figures 1 and 2). Treatment concentrations also significantly (P<0.05) increased the number of axillary shoots generated per upright terminal shoot (Figure 1). Overall, there was negative relation between terminal shoot growth and axillary shoot growth. The treatments receiving the highest concentration of Fe-DTPA fertilizer displaying the least amount of terminal shoot growth, yet highest number of axillary shoot growth per upright across measurement events (Figures 1 and 2).
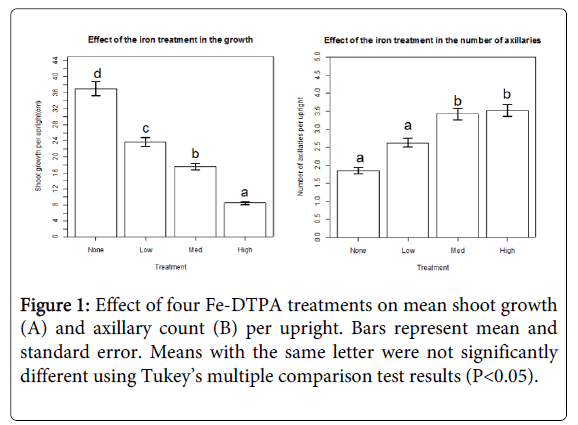
Figure 1: Effect of four Fe-DTPA treatments on mean shoot growth (A) and axillary count (B) per upright. Bars represent mean and standard error. Means with the same letter were not significantly different using Tukey’s multiple comparison test results (P<0.05).
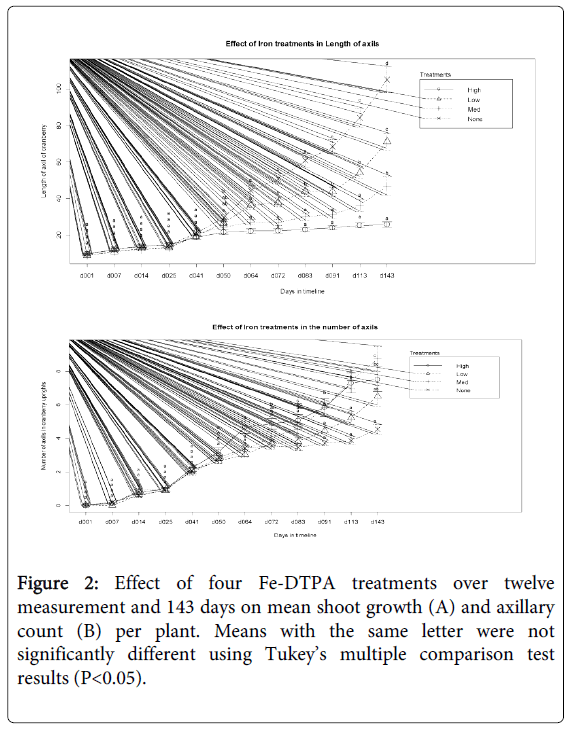
Figure 2: Effect of four Fe-DTPA treatments over twelve measurement and 143 days on mean shoot growth (A) and axillary count (B) per plant. Means with the same letter were not significantly different using Tukey’s multiple comparison test results (P<0.05).
There was a significant (P<0.05) decrease in plant biomass from uprights grown with the high Fe-DTPA concentration, with total mean fresh weight (10.8 g) 3.4 and 2.8 fold below the control and low treatments, respectively (Figure 3). Final shoot biomass for high Fe-DTPA concentration treatment plants was 56%, whereas other treatments averaged 80-96%. Overall, there was greater fluctuation detected in shoot fresh weights among treatments than shoot growth measurements. Both variables are important and could impact a plant’s ability to sustain berry growth and development.
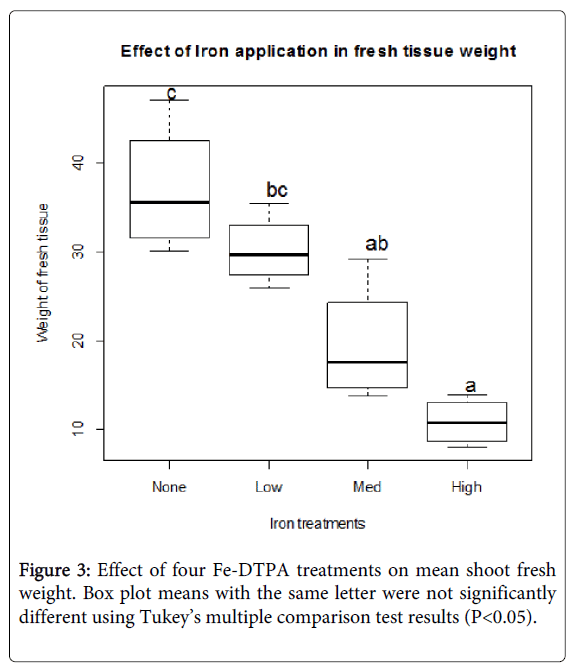
Figure 3: Effect of four Fe-DTPA treatments on mean shoot fresh weight. Box plot means with the same letter were not significantly different using Tukey’s multiple comparison test results (P<0.05).
Although no data were recorded, root systems also appeared to be impacted by the high Fe-DTPA concentration treatments. Acute stunting, discoloration, and injury to plant roots were consistently observed at higher Fe-DTPA concentrations, as well as substantial reduction in root mass. Photographs representative of root injury patterns are presented in (Figure 4).
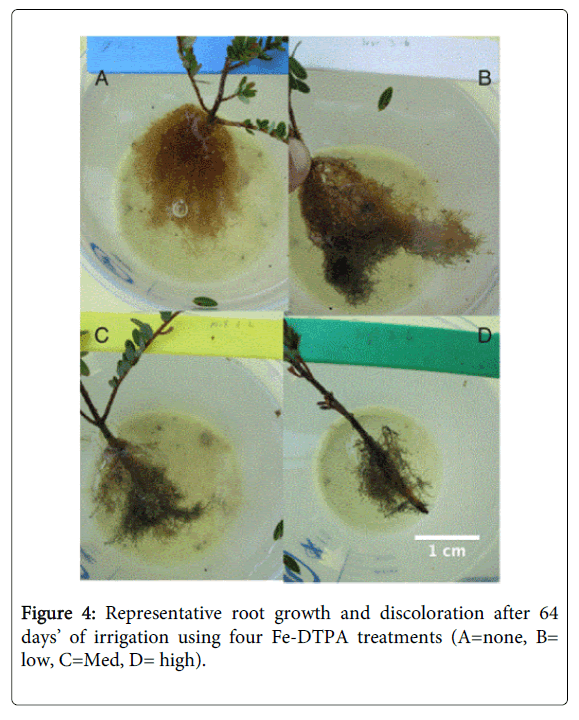
Figure 4: Representative root growth and discoloration after 64 days’ of irrigation using four Fe-DTPA treatments (A=none, B= low, C=Med, D= high).
Nutrient analysis
Nutrient analysis showed the following overall mean nutrient level: nitrogen (1.4-1.70%), phosphorous (0.12-0.16%), potassium (0.67-0.88%), calcium (0.33-0.53%), magnesium (0.14-0.20%), sulfur (0.09-0.16%), manganese (45.0-76.5 ppm), Fe (76-625 ppm), zinc (14.7-19.7 ppm), aluminum (56.5-81.5 ppm), copper (5.8-8.3 ppm), sodium (358-1583 ppm), and boron (47-59 ppm) (Table 2). Natural iron in field plants collected from 15 marshes with no reported history of iron toxicity ranged from 142.8 ppm to 880.7 ppm (Table 2).
| Treatments | Flowering span (days) |
Fresh weight of whole spike (g) |
Diameter of floret (cm) |
Length of floret rachis (cm) |
Longevity of spikes (days) |
Vase life of spikes (days) |
|---|---|---|---|---|---|---|
| T1 | 23.10 | 88.14 | 7.20 | 54.03 | 13.83 | 13.17 |
| T2 | 21.33 | 80.21 | 6.38 | 45.25 | 12.85 | 11.41 |
| T3 | 21.47 | 85.32 | 6.73 | 50.29 | 13.52 | 12.90 |
| T4 | 22.12 | 86.19 | 7.39 | 51.18 | 14.29 | 13.67 |
| T5 | 21.50 | 82.15 | 6.36 | 50.73 | 13.20 | 12.83 |
| T6 | 24.17 | 90.03 | 7.98 | 55.24 | 15.17 | 14.32 |
| T7 | 19.65 | 78.15 | 6.20 | 44.61 | 10.95 | 10.38 |
| T8 | 21.97 | 83.89 | 6.95 | 52.10 | 12.03 | 11.65 |
| T9 | 21.73 | 80.07 | 6.66 | 49.55 | 12.35 | 12.29 |
| T10 | 20.98 | 78.23 | 5.96 | 40.94 | 12.00 | 11.17 |
| T11 | 19.48 | 75.10 | 6.02 | 43.62 | 10.03 | 9.28 |
| T12 | 20.07 | 79.50 | 6.27 | 43.01 | 11.33 | 11.19 |
| S.Em.± | 1.302 | 2.996 | 0.333 | 2.492 | 0.713 | 0.632 |
| C.D. at 5% | NS | 8.79 | 0.98 | 7.31 | 2.09 | 1.85 |
| C.V.% | 10.51 | 8.31 | 8.66 | 8.92 | 9.79 | 9.11 |
Table 2: Effect of organic, inorganic and biofertilizer on flowering and flower quality in gladiolus cv. Psittacinus hybrid.
Controlled experiments using Sequestrene 330 Fe-DTPA produced clear toxicity on vegetative growth and plant death with increased concentrations (Figures 1-4). Nutrient analysis (Table 2) showed that most shoot nutrient treatment means remained close to the sufficiency ranges recommended by Davenport et al. [3]. However, both iron and sodium increased significantly (P<0.05) with increased Fe-DTPA concentrations (Table 2). Iron ranged from 76 ppm (none) to 627 ppm (high), while sodium ranged from 359 ppm (none) to 1583 ppm (high), both of which increased in a linear fashion with increasing irrigation concentrations with Fe-DTPA. While iron was 8.3 times higher in the high Fe-DTPA treatment than in the control treatment, sodium increased 4.4 times in the high treatment versus the control treatment. An increase in iron concentrations was expected over the 143-day span of the experiment, but the sharp sodium increase was unexpected in the cranberry tissues tested. In fact, sodium levels actually increased in tandem with iron concentration, perhaps owing to its presence in the Fe-DTPA. Indeed, commonly used synthetic iron chelates such as Sequesterne 330 (Novartis Crop Protection; Greensboro, IL) usually consist of Fe3+ ions, EDTA or DTPA, and sodium (Na+) or ammonium (NH4+) ions [9]. In this case, Sequestrene 330 is iron salt containing sodium.
The only direct study thus far of iron tolerance in cranberry is Medappa and Dana [10] who investigated the effects of wide range of ferric ethylenediaminetetraacetic (Fe-EDTA) concentrations on shoot and root growth. They reported iron toxicity in cranberry and plant death at treatments of 50 ppm over five weeks, but they noted that plant death could be attributed to interactions with the chelate rather than a direct consequence of iron concentration [10]. Experiments conducted using a different iron sources such as ferrous sulfate instead of chelated forms [11] have demonstrated that Fe is not particularly toxic in several plant species previously deemed highly susceptible [12,13]. Thus, research on Fe toxicity based only on chelated iron forms may have led researchers to the conclusion that Fe itself is highly toxic in many species [11]. In the present study, field iron tissue ranged from 143 to 881 ppm among 15 Wisconsin cultivated marshes with no apparent iron adverse reaction (Table 3).
| Treatments | No. of spikes per plant | No. of spikes per net plot | No. of spikes per ha(lakh no.) |
|---|---|---|---|
| T1 | 2.08 | 18.75 | 2.57 |
| T2 | 1.77 | 15.91 | 2.21 |
| T3 | 1.73 | 15.60 | 2.16 |
| T4 | 2.03 | 18.43 | 2.53 |
| T5 | 1.67 | 15.07 | 2.09 |
| T6 | 2.20 | 19.82 | 2.87 |
| T7 | 1.43 | 12.90 | 1.79 |
| T8 | 2.00 | 18.01 | 2.50 |
| T9 | 1.90 | 16.88 | 2.34 |
| T10 | 1.55 | 13.95 | 1.94 |
| T11 | 1.28 | 11.55 | 1.60 |
| T12 | 1.52 | 13.65 | 1.90 |
| S.Em.± | 0.106 | 0.977 | 0.139 |
| C.D. at 5% | 0.31 | 2.86 | 0.41 |
| C.V.% | 10.46 | 10.66 | 10.97 |
Table 3: Effect of organic, inorganic and biofertilizer on spike yield in gladiolus cv. Psittacinus hybrid.
This range is greater than the recommended optimal ranges in cranberry, 40-80 ppm, “normal concentration” level of >20 ppm [2,4,14], and higher than the ranges observed by Doughty [15], who considered iron levels of 279 ppm to be very high. Also, the highest level of iron detected under cultivated conditions (881 ppm) was higher than the iron levels (627 ppm) detected in the high iron treatment regime of the current experiments, which resulted from irrigation with water at 56 ppm Fe (Tables 2 and 3) and was similar to death eliciting treatments of Medappa and Dana [10]. Therefore, the high range of iron detected in the current commercial field survey indicates that cranberries may tolerate higher concentrations of iron than previously believed. Albeit, some of the high Fe readings in field tissues could be due surface deposition, our data showing high iron in tissue of healthy plants grown under commercial conditions (Table 3) do not support the idea that phytotoxicity in our cranberry study was caused iron accumulation.
On the other hand, sodium levels in the cranberry tissues treated using chelated iron in the current experiment were high in the medium and high treatments (898 and 1583 ppm, respectively) compared to observed range (37-488 ppm) in the field (Tables 2 and 3). Salt water damage has been documented during hurricanes [16]. Similarly, visible damage attributed to road salt has also been recorded in cranberry near highways and from direct salt deposition or vehicle overspray in marshes [17]. However, the sodium toxicity work of DeMoranville et al. [18] did not show drastic reduction on shoot weight, abnormal axillaries and roots, and plant death associated with similar sodium tissue concentrations (i.e., 1583 ppm) as observed in our study. High Na caused only minor growth reduction (~10%) that actually stimulated axillary growth into runners. Moreover, DeMoranville et al. [18] routinely observed tissue Na between 1000-3000 ppm with no observable leaf browning or drop and no tip dieback. Therefore, the severe reactions observed in our cranberry study could not be attributed to sodium toxicity.
In order to further assess the threat of iron in cranberry, and in light of the possible toxicity of cranberry to chelators or other components in chelated iron sources, a systematic study of iron levels in cranberry and its effects needs to be undertaken using both natural and controlled systems using an alternative of iron sources (e.g., sodium free chelates, ferrous sulfate, ferric citrate, ferric glucoheptonate, or ferrous ammonium sulfate). However, it might be difficult to measure iron under field and greenhouse due to its instability and negligible effects due to natural fluctuation [19], and as demonstrated in the current experiment, observed toxicities may not be due to iron.
Controlled experiments using Sequestrene 330 Fe-DTPA produced clear toxicity with increased concentrations. Iron tissue levels increased with greater Sequestrene 330 concentrations, but the greatest iron concentration observed in greenhouse experimental conditions was lower than the naturally occurring high levels of iron observed in field-grown plants. However, high sodium was also detected in tissues treated with Sequestrene 330, but the observed ranges likely could not produce toxicity in cranberry. In conclusion, the toxicity observed is likely due to specific toxicity of cranberry to the chelators in the Sequestrene 330. At this moment, is not clear if iron toxicity in cranberry is a common or even real. More systematic nutrient studies in cranberry need to be conducted. Elucidating the benefits or risks to particular accumulations of iron and other micronutrients are needed in order to establish sufficiency ranges, thresholds, and help growers better understand toxicity risks.
Rom. 8:31-39. Special thanks to Carolyn DeMoranville for her comments about this research. This research was supported by Wisconsin State Cranberry Growers Association, Cranberry Institute, and USDA-ARS (project no. 3655-21220-001-00) provided to J.Z. and S.S.