Anatomy & Physiology: Current Research
Open Access
ISSN: 2161-0940
ISSN: 2161-0940
Review Article - (2016) Volume 6, Issue 3
Mouse models are important in Biomedical- and Life Sciences. Recently it has been stated that the genetically engineered mouse models for contemporary Obesity and/or Type-2 diabetes (DM2) will in most cases deliver heavily metabolically abnormal animals due to changed gut microbiotics with increased anaerobic CO2 production. Therefore, it can be questioned if indirect calorimetry which it at this moment “the golden standard” in all mouse laboratories over the world, still can be practiced because it is dependent on the conversion factor Respiratory-Quotient (RQ-value: CO2/O2) for expressing energy expenditure into Heat production (HP) in [mW]. Direct calorimetry, however, can accurately quantify HP and thus Basal Metabolic Rate (BMR) in both metabolically normal and abnormal states. In this first manuscript we determined physical properties and characteristics of a former aquatic flow-through twin detection Setaram calorimeter used for measuring Basal Metabolic Rate of fish under adverse environmental conditions (anoxic-, hypoxic-, acid- environmental water). Our objective of this study was to determine if this Setaram calorimetric system with stainless steel vessel of 1 liter could be used in the physical medium air in order to measure energy expenditure in mouse models. This first [Technical note] describes the physical properties and characteristics in the medium water and air of this calorimetric set up. Characteristics of the calorimetric system in air were for the initial baseline over 80 minutes a heat flow of 13.44 ± 1.611 (mW) and for the end baseline over 175 minutes 14.95 ± 1.79 (mW) so the ∆-Baseline (initial –end) was 13.44-14.95 = -1.51 (mW). The drifting baseline over the total experiment of 33h was (14.95 / 13.44) * 100% = 1.1124% with a drifting baseline per hour was 0.035 % per hour which are common values for twin detection Calvin calorimeters. We calculated based on the heat transfer characteristics from water to stainless steel (aquatic calorimeter) in combination with the specific heat capacity of water Cp-water of 4,1884 [kJ K-1] vs Cp-air is 0.001293 [kJ K-1] an aquatic time-constant τwater of 30.48 min vs τair or = 0.019605 min. Because the τair was so small deconvolution techniques (earlier successfully applied in water), correcting for the lag time of the system had no effect. This study also includes the first successful “principle of proof” pilot-experiment with one single mouse locked up for ≈ 14 h proves that measuring the energy expenditure with a single mouse in the 1 liter stainless steel calorimetric vessel in the medium air corresponds to measured values of wild strain mouse used in indirect calorimetry (IMC) (of ≈10 mW/g). A precondition was that in order to prevent suffocating of the mouse in the one liter vessel both vessel (Measurement and Reference: twin detection measurement were supplied with one liter of air per minute- with this former aquatic flow-through twin detection Setaram calorimeter. Although there is in Bio-calorimetry a switch to cell-, (sub)cellular, molecular and biomolecular interactions, this study with “whole animal” calorimetry proves that at the organismal level important fundamental questions are still unanswered and open related to the pathogenesis of human diseases like Obesity and metabolic related disorders like Type-2 Diabetes and more fundamental Life Sciences related processes like ‘ageing’. In addition, also the research area of nutraceuticals can be tested with DMC.
Keywords: Biological calorimetry; Physical properties medium; Mouse; Energy expenditure
The Basal metabolic rate (BMR) which can be considered as the ‘minimal metabolism’≈ ‘the Energy Costs to stay alive’, and the closely related resting metabolic rate (RMR), is the amount of daily energy expended while at rest in a neutrally temperate environment, in the postabsorptive state (meaning that the digestive system is inactive, which requires about eight hours of fasting in murents=mouse models). The release of energy in this state is sufficient only for the functioning of the vital organs, the heart, lungs, nervous system, kidneys, liver, intestine, sex organs, muscles, and skin [1]. The common house mouse (Mus musculus L.) is the most used laboratory animal in medical research [2]. Besides being genetically similar to humans, mice are small and inexpensive to maintain. Their short life span and rapid reproductive rate make it possible to study disease processes in many individuals throughout their life cycle. Above of all following publication of the sequence and analysis of a mouse strain in December 2001 [3], the mouse became the animal model of choice for most laboratory experiments. The potential of mice for genetic manipulation means that their use is now often favored over rats and other rodents. In addition, the mouse makes an excellent model for human disease because the organization of their DNA and way their genes are expressed is very similar to humans. Their reproductive and nervous systems are like those of humans, and they suffer from many of the same diseases such as cancer, diabetes and even anxiety. Manipulating their genes can lead them to develop other diseases that do not naturally affect them, and as a result research on mice has helped understanding of both human physiology and the causes of disease. The last 10-15 years, genetically engineered mouse, including transgenic mouse, knock-out mouse, knock-in mouse, and knock-down mouse, have been cultured. At this moment, up to 478 inbred strains have been cultured and more than 2,000 mutant strains have been established by either spontaneous, inducible methods or transgenic technology and have been conserved for laboratory purposes [2]. This could be established because in correspondence with the “Human Genome Project (HGP) [4,5], the DNA sequencing of the entire genome of mouse has been completed (Mouse Genome Sequencing Consortium 2002) [3]. As an animal model different genetic variants and strains have been developed e.g. for human related disease like e.g. for the maturity onset of the young (MODY) syndrome [6], syndromes of severe Insulin Resistance [7], mitochondrial diabetes [8], the different obese mouse strains like ob/ob, db/db, and fa/fa [9], but also for “fundamental processes of Life Sciences” like ageing. In order to give an answer of the point mutations and deletions of mitochondrial DNA (mtDNA) accumulate in a variety of tissues during ageing in humans and this increase in somatic mtDNA mutations is associated with reduced lifespan [10-14] created a homozygous knock-in mice that expresses a proof-reading-deficient version of POlgA, the nucleusencoded catalytic subunit of mtDNA polymerase. Their study with the knock-in mice strain showed that the animal developed an mtDNA mutator phenotype with a threefold to fivefold increase in the levels of the point mutations, as well as increased amounts of deleted mtDNA. This increase in somatic mtDNA mutations is associated with reduced lifespan and premature onset of ageing related phenotypes. Their results clearly provide a causative link between mtDNA mutations and ageing phenotypes in mammals [14]. An example of a knock-out mouse strain for the senescence marker protein-30 (SPM-30) had after three months an increased mortality rate, and many alterations in triacylglycerol-, cholesterol- and phospolipid levels compared to wild mice. The researchers suggested that the SPM-30 gene has an anti-apoptotic (apoptotic=programmed cell death) function with a wide spectrum and that this strain might be an useful tool for ageing and biological monitoring [15]. These different murent-models thus provide us with a valuable highly appreciated and valuable tool to deepen our view and give perspectives via e.g. a reductionistic approach to unravel the fundamental mechanisms or to test several hypotheses which may play a causative role in the induction of the pathogenesis of certain human diseases e.g. Obesity, Type-2 diabetes (DM2) and the mechanism(s) behind processes of Life Sciences like e.g. ageing. Until this moment the only applied method to measure the BMR in mouse-models is by indirect calorimetry, Jenesen et al. [16], in some cases complemented by telemetric monitoring of body core temperature, sometimes in combination with locomotion activity registration in mice and rats [17,18]. The indirect calorimetric method has until this moment largely been supplanted in determining the BMR in mouse models owing to the relative ease and ready commercial availability of the latter technique. This technique involves measurement of CO2 produced and O2 consumed by an organism. These measurements are then used of the ratio of O2 consumed and CO2 produced, which gives the respiratory quotient (RQ), a relative assessment of protein, carbohydrate and lipid oxidation. Each substrate has its own characteristic RQ value, for the substrates protein, carbohydrate and lipids these are respectively, 0.809, 1.000 and 0.711 [1]. However new insights nowadays based on a) Methodological grounds; b) Physiological information regarding the “created” transgenic or obese mouse models; c) Phenotypical variations; d) Not optimal husbandry/housing conditions like a temperature out of the thermo-neutral zone of murents, can be critically reviewed. Therefore the question can be posed if the indirect calorimetric method (based on the conversion factor RQ) is a correct approach to calculate the heat equivalent of energy output, routine metabolic rate (RMR) or BMR. According to researchers Kiyala and Ramsey [19], direct calorimetry with mouse models is at this moment “the golden standard” –an undeniable prerequisite to prevent tremendous experimental flawsfor reliable Biomedical and Life-Sciences related experimental work with mouse models in nearby future. Accordingly, it seems obvious that new research employing simultaneous direct and indirect calorimetry (total calorimetry) will be essential to validate respirometric BMR and RMR phenotyping in existing and future pharmacological and genetic models of Obesity and Type-2 Diabetes [19]. The initial hypothesis we wanted to test in this pilot experiment was if it was possible by direct calorimetry the measure energy expenditure, the RMR and BMR of a single mouse. If this pilot would give positive results the combination of direct and indirect calorimetry has to be possible to determine the exact order of magnitude of experimental errors in obtained dataset using indirect calorimetry in the traditional “mouse-respirometry” cages all around over the world applied at this moment.
Twin-detection Setaram microcalorimeter for whole animal studies
In collaboration with a calorimeter manufacturer, Setaram, Caluire, France we developed in the past an aquatic twin detection flow-through microcalorimetric system with stainless steel calorimetric vessels of 1 litre for mainly experiments with fish in the range of 20-120 gr [20-28]. Because in Biomedical Science the need for direct calorimetry, especially with obese and transgenic mouse models was very recently stipulated [19] we hoped that this from origin aquatic flow-through calorimeter could be rebuild to a mouse calorimeter by eliminating the aquatic pre-microcalorimeter (LKB-bioactivity monitor). An advantage was that the 1 litre vessel was large enough to house individual adult mouse in the range from 20-40 g bodyweight. The temperature of the Setaram calorimeter is roughly stabilized by an external loop filled by dodecane and an internal loop for fine-tuning of the temperature. The signal resolution is 50 μWatts with a drift < 0.1 mWatts per 24 hours. Success was not guaranteed at forehand due to physical determined differences in heat capacity of the different media (water vs. air) and consequently different time constants. Therefore we were lucky that the developed (original aquatic) calorimetric set up was developed in such a way that for a resolution of <50 μW the ΔT between the heat sink and the animal chamber must be <10-4°C. This temperature stability and heat flow sensitivity can only be reached with a twin detection system. Furthermore a low heat loss of <5% was reached by using 3 counter current heat exchangers, two within the calorimeter systems and one on top of the animal chamber within the measuring system. The calorimetric block contains identical measurement and reference vessels. The signals of both vessels are affected by environmental (=temperature) changes in an equal way such that subtraction of both signals results in a stable baseline. After introduction of the single mouse of 40.1 g in this pilot experiment the baseline stability was reached within 4 hours. The calorimetric vessels are suitable for a constant flow through of air so that simultaneous indirect calorimetry can be applied if gas analysing equipment for O2 (inlet) and CO2 (outlet) would be available in future studies. For this pilot experiment we were only curious if we could measure under calibrated conditions the heat production in air in a by a and a continuous flow rate of air through the 1 liter stainless vessels of ≈ 1 liter per minute (heat loss!), in this way refreshing the aerial environment for the mouse in the 1 litre calorimetric vessel. The specific heat capacity of water respectively air is: Cp-water of 1 litre of water = 4,1884 [kJ K-1] and for water.
Calibration was performed by electrical calibration by means of an electrical current through a circular resistor (1000 Ω) mounted in the M-vessel during a period of 5 hours. By calculating the Area under the Curve (AUC) this delivers us a sensitivity coefficient (μV/mW ), which transforms the by the thermopiles in the calorimetric block Voltage signal into a heat production signal (power in mW) (Figure 1).

Figure 1: Principle of the twin-detection method in the period of an aquatic flow-through microcalorimeter: signals of both vessels are affected by environmental (=temperature) changes in an equal way such that subtraction of both signals results in a stable baseline. The experiment starts and finishes with an electrical calibration. During a defined time-interval (here 6 hours) an electrical current of 0.315 A and a Voltage of 3.164 V is applied to a resistor of 1000 Ω mounted inside the measurement vessel.
Miscellaneous equipment twin-detection Setaram microcalorimeter
The following miscellaneous equipment and electronic devices for this air-based micro-calorimeter to measure the energy expenditure of a single mouse: i) Twin detection micro calorimeter (Setaram GF108); final temperature stabilization by means of three heat exchangers, two inside the calorimeter and one on top of the 1 litre calorimetric vessel. ii) Three 1 litre stainless steel vessels. In one (reserve) is at the top build in a receiver plate of Data Sciences International (DSI), St. Paul, USA, for the telemetry transmitters build in the peritoneal cavity of the mouse to be able to measure in addition via this invasive way via telemetry: a) Body core temperature, b) Heart-rate frequency and produce an c. Electrocardiogram (ECG) [corresponding like performed for goldfish during metabolic depression in: [26] iii) Automatic alternating Brückert three-way valve controlled by the computer for separately gas-analysing equipment for indirect calorimetry. iv) Miscelaneous electronic equipment for the calorimetric set up like: amplifier (Setaram A85), voltmeter (Setaram VN), thermometer (Setaram TN2) temperature programmer and controller (Setaram), thermal safety unit (Setaram TS1); v). Special developed software and interface for this Setaram twin-detection micro-calorimeter.
Experimental set up
Direct calorimetry experiment (pilot) with a white albino male house mouse (Mus musculus L.) of 40 g in a Setaram GF108 twin detection micro-calorimeter at a room and calorimeter temperature of 21C and a continuous flow rate of air through the 1 liter stainless vessel of ≈ 1 liter per minute (both M- and Reference Vessel). The experiments starts with an electrical calibration by means of an electrical current through a circular resistor (1000 Ω) mounted in the M-vessel during a period of 5 hours. With sensitivity coefficient of 144 μV/mW in the software the produced heat in the M-vessel corresponds to 100 mW => calibration. Thereafter the mouse is placed in the Setaram calorimeter, in this pilot with a piece of apple to avoid desiccation and starvation. For six hours we don’t get a signal (large spikes) because the Setaram calorimeter needs to reach a new thermal equilibrium after opening it and the 1 liter stainless M-vessel (=animal chamber) to insert the mouse in the calorimetric system. Thereafter we get a usable Heatproduction signal of the 40 g mouse for a period of ≈12.5 hr. After taken out the mouse out of the calorimeter and a stabilization period of approximately 5 hours the experiment is finished with an electrical calibration by means of an electrical current through a circular resistor (1000 Ω) mounted in the M-vessel during a period of 5 hours.
Experimental mouse for this pilot experiment and handling of the animal
A Swiss male wild-type albino mouse (age 8-12 weeks) of 40.1 g was bought in a local pet shop. The pilot experiment with the mouse was performed outside the University. The animal was carefully handled and was locked up in a 1 liter vessel with sufficient air (1 liter per minute), food and water and after 14 hours released and brought back at a the local pet shop where it was bought.
Calculations
Calculation of the time constant τair in air and comparison with the time constant in water τwater: In correspondence with a previous manuscript [22] the one litre stainless vessel filled solely with water without animals (at that time an aquatic flow-through calorimeter) it was concluded that there was an ideal pattern of mixing in the vessel. Therefore it was concluded that the time constant τwater of the timecontinuous system was determined mainly by the heat capacity of water and the heat transfer coefficient from water to stainless steel. Also the the time constant τwater could be calculated from:
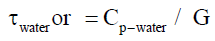 (1)
(1)
Where the heat capacity Cp of 1 litre of water (0.998 kg) was with a specific heat (in the range 293-373 K) = 4.18 * 103 [J kg-1 K]. So the specific heat capacity Cp-water of 1 litre of water = 4,1884 [kJ K-1].
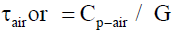 (2)
(2)
Similar for air (mixture) the density is 1.293 [kg m-3] => 1000 litre = 1 m3 so 1 litre of air = 0.001293 [kg] and 1 kg air = 1/0.001293 = 773,3952049 [litre]. The heat capacity Cp of 1 litre of air (at constant pressure Cp and T=273 K) = 1,00 * 103 [J kg-1 K] = 1,00 [kJ kg-1 K]. So 773,3952049 [litre] of air = 1,00 kJ K-1. So the specific heat capacity Cp-air of 1 litre of air =
1,00 / 773,3952049 [kJ K-1] = 0.001293 [kJ K-1].
The thermal conductance G sees (1) is described by
G = α * A (3).
Where α is the transfer coefficient i) From water to stainless steel (αwater = 50 W K-1 m-2) and ii) From air to stainless steel (αair = 24 * 10-3 W K-1 m-2) and A is the exchanging surface of the stainless steel 1 litre vessel which can be described by
A = (2*π* r * h) + π* r2 (4)
Where r = 5.4 [cm] and h = 10.8 [cm]. Thus A corresponds to (2*π * 5.4 * 10.8) + π * 5.42 = 0.0458 m2. Consequently, the thermal conductance Gwater = 50 * 0.0458 = 2.29 [W K-1] and the thermal conductance Gair = 24 * 10-3 * 0.0458 = 0.0010992 [W K-1]
Following for water equation (0). (τwater or = Cp-water / G = 4188 [J K-1] / 2.29 [W K-1] = 1829 s. which is 30.48 min. Similar following the air equation (1).
τair or = Cp-air / G = 0.001293 [kJ K-1] / = 0.0010992 [W K-1] = 1.176310 s. ≈ 0.019605 min.
Deconvolution techniques, desmearing of calorimetric data: The time constant τwater of the developed 1 liter differential flow-through calorimeter (Sétaram GF 108) was estimated for the medium water to be 30.48 minutes [22]. While τair for the medium air was only 0.019605 min. Data for an electrical calibration in air were desmeared using the time constant τair of 0.019605 min. based on the method of [29], with one time constant.
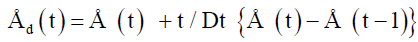 (5)
(5)
With Åd(t) is deconvoluted signal, Å (t) is original data point to be corrected, Å(t-1) is data point registered one minute prior to Å (t) and τ is the time-constant corresponding to 33 minutes, Δt is the sampling rate equal to 1 minute. In addition to the method of [29] data were filtered in the spreadsheet. For every new smoothed point the mean value of ten data points prior to this point were used according to:
With Åa(t) = the mean value of ten data points.
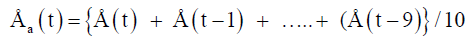 (6)
(6)
Results
Our pilot experiment had to meet three demands:
1. Are we able in the (physical) medium air with a heat capacity of 0.001293 [kJ K-1] in comparison to the heat capacity of water of 4,1884 [kJ K-1] and varying time constants of water vs. air, respectively 30.48 min. vs. 0.019605 min. to:
• Gain stable thermic calorimetric conditions and
• Measure the heat production of a warm-blooded mammal single mouse
• Are we able to perform long lasting experiments (≈ 12 hours or longer) with one single mouse in this one liter stainless steel calorimetric vessel and -to prevent suffocation of the animal- lead continuous a constant air flow of 1 liter per minute through the two vessels (twin detection) without affecting the stability of the baseline.
2. Can we get in a reliable way data - by means of electrical calibration using a resistor of 1000 Ω giving a calibration factor expressed in μV/mW - which would allow us to quantify the energy expenditure of a single mouse in order to measure the BMR. The results of this pilot experiment determining the BMR of a single mouse by direct calorimetry are given in Figure 1.
• Results considering the direct calorimetric set up are given in Figure 1. Comparing the drift between the initial baseline and at the end of the experiment the total drift over the experimental period of 33 h. corresponds to 1.1124% which is at a Δ-(baseline) of 1.51 corresponding to the RMR of the mouse of 40.1 gr 810.74 ± 192.483 (mW) only 0.0014%. Or compared to the Maximal Metabolic Rate (MMR) of 1457.59 (mW) only 0.00010% or compared to the SMR of 411.35 (mW) only 0.0037% (Table 1).
| No. | Method | Details experiment | RMR or BMR/g tissue | Reference |
|---|---|---|---|---|
| 1. | Direct | Mouse 40.1 g, 21ºC 14 h mouse in calorimeter |
BMR≈10.26 mW/g | Results in this research article |
| 2. | Indirect | Food intake, 28ºC | RMR≈10.98 mW/g | Speakman and McQueenie [32] |
| 3. | Indirect | 5º,15º,25º,30ºC | RMR males≈10.30 mW/g RMR-females≈12.03 mW/g |
Selman et al. [33] |
| 4. | Indirect | N=265, 30ºC, RQ=0.95, flow rate500-700 ml/min | C57BL/6 BMR=8.05 mW/g |
Johnston et al. [34] |
| 5. | Indirect | 12.0-19.6 g T=25-30ºC, RQ=0.82 |
BMR=10.81 mW/g | Hussein [35] |
Table 1: Basal Metabolic Rate (BMR) of this pilot direct calorimetric experiment in comparison to indirect BMR calorimetric literature data. Details of the different conditions of the several experiments like Temperature, Respiratory Quotient (RQ), and flow rate through the indirect calorimetric cages are given.
• Table 2 shows the Characteristics of the direct calorimetric system obtained during a direct calorimetric experiment of 33 hours like baseline stability and/or drift and experimental error in comparison to the finally measured heat production of the mouse (Table 3). Characteristics of the calorimetric system in air were for the initial baseline over 80 minutes a heat flow of 13.44 ± 1.611 (mW) and for the end baseline over 175 minutes 14.95 ± 1.79 (mW) so the Δ-Baseline (initial –end) was 13.44-14.95 = -1.51 mW. The drifting baseline over the total experiment of 33h was (14.95 / 13.44) * 100% = 1.1124% with a drifting baseline per hour was 0.035 % per hour which are common values for twin detection Calvin microcalorimeters.
| Parameter | Measured Value |
|---|---|
| Baseline start | |
| *interval | 80 minutes |
| *Heat flow (mW) | 13.44 ± 1.611 |
| Baseline end | |
| *interval | 175 minutes |
| *Heat flow (mW) | 14.95 ± 1.79 |
| ∆-Baseline (initial –end) | 13.44-14.95 = -1.51 mW |
| Drifting Baseline over total experiment 33 h | (14.95 / 13.44) * 100% = 1.1124% |
| Drifting Baseline per hour | 0.035 % per hour |
| Electrical calibration with resistor 100 Ω | |
| sensitivity coefficient of 144 µV/mW | |
| First calibration for 5 h. with resistor 100 Ω | Stable plateau value: 99.71 ± 0.333 (mW) |
| Second calibration for 5 h. resistor 100 Ω | Stable plateau: 99.67 ± 0.307 (mW) |
Table 2: Characteristics of the direct calorimetric system obtained during a direct calorimetric experiment of 33 hours like baseline stability and/or drift and experimental error in comparison to the finally measured heat production of the mouse (Table 3).
• The total direct calorimetric experiment including initial and final calibration is depicted in Figure 2 and lasted ≈33 h. The mouse was from 5-18 hours (X-axis) => 13.55 h (≈14 h) in the one liter calorimetric vessel. The heat production data of this single mouse are depicted in Figure 3 and Table 3. The micro-calorimeter regained its thermal equilibrium within 5 hours after putting the animal in the 1 liter calorimetric vessel so actually the animal was for ≈19 hours in the 1 liter calorimetric vessel.

Figure 2: Direct calorimetric experiment in the medium air with a single mouse of 40.4 g. The experiment starts with an electrical calibration until 100 Ω and finishes with a similar electrical calibration. Time without signal due to opening the calorimeter and putting the mouse in the 1 liter stainless steel calorimetric vessel an also taken the animal out gives during a period of five hours no signal (≈heat production data) because the calorimeter has to regain its thermal equilibrium. The mouse was for approximately 12.5 hours in the calorimeter and reaches its lowest level of ≈10 mW/g at the end of the experimental period.

Figure 3: Energy expenditure during the 14 hours the single mouse of 40.1 g was in the Direct Setaram twin-detection calorimeter at 21°C.
| Parameter | Measured Value |
|---|---|
| Heat Production mouse of 20.4 g | |
| -5 h (Mean ± stdev) (note exception: 48 min.) | 1347.1 ± 70.81 (mW) (Figure 3) |
| -6 h (Mean ± stdev) (60 minutes) | 1109.5 ± 99.57 (mW) (Figure 3) |
| -7 h (Mean ± stdev) | 874.0 ± 113.66 (mW) (Figure 3) |
| -8 h (Mean ± stdev) | 753.8 ± 28.25 (mW) (Figure 3) |
| -9 h (Mean ± stdev) | 829.4 ± 65.52 (mW) (Figure 3) |
| -10 h (Mean ± stdev) | 706.9 ± 29.87 (mW) (Figure 3) |
| -11 h (Mean ± stdev) | 677.7 ± 17.79 (mW) (Figure 3) |
| -12 h (Mean ± stdev) | 743.9 ± 52.94 (mW) (Figure 3) |
| -13 h (Mean ± stdev) | 694.9 ± 15.37 (mW) (Figure 3) |
| -14 h (Mean ± stdev) | 721.0 ± 59.64 (mW) (Figure 3) |
| -15 h (Mean ± stdev) | 888.4 ± 23.67 (mW) (Figure 3) |
| -16 h (Mean ± stdev) | 792.9 ± 84.04 (mW) (Figure 3) |
| -17 h (Mean ± stdev) | 668.8 ± 29.75 (mW) (Figure 3) |
| -18h (Mean ± stdev) (note exception: 32 min) | 630.3 ± 21.40 (mW) (Figure 3) |
| -Total (RMR): 5 - 18 h (Mean ± stdev) | 810.74 ± 192.483 (mW) |
| -Maximal Metabolic Rate (MMR) (1 min.) | 1457.59 (mW) |
| -lowest BMR-1 (interval 1 minute) | 411.35 (mW) |
| -mean BMR-2 (interval 8 minutes) | 428.6 ± 17.14 (mW) |
| RMR :BMR-1 | 1.97 fold |
| MMR : RMR | 1.80 fold |
| MMR : BMR-1 | 3.54 fold |
Table 3: Biological values obtained by the direct calorimetric experiment (in air) of a single mouse of 40.1 g where the metabolic rate had been measured for 13,55 hours (≈14 h) at a room and calorimetric temperature of 21°C.
In Figure 2, we see this pilot direct calorimetric experiment in a calibrated (switched to air) Setaram twin-detection calorimeter with a single mouse of 40.1 g. reflecting its energy expenditure.
In Figure 3 and Table 3 some biological characteristics of this direct calorimetric experiment with a mouse of 40.1 g at a room and calorimeter temperature of 21°C and a continuous flow rate of air through the 1 liter stainless vessel of ≈ 1 liter per minute (both M- and Reference Vessel) are mentioned.
From Figure 3 we can see that (although the mouse was already for 4-5 hours in the 1 liter calorimetric vessel before the calorimeter reached its thermal equilibrium) that the animal had for three hours (hour 5,6,7) still an increased heat production of >1000 mW. Thereafter during the following period of 10 hours (8-18 hours) it fluctuated roughly between 600-900 mW. A detailed description of this experiment in terms of energy expenditure is given in Table 3.
The minimal level (BMR) over a time interval of one minute of this pilot we found for a single mouse in our air-based Seteram twin detection micro-calorimeter corresponded to ≈ 411.35 mW/ 40.1 g (≈10.26 mW/g). This agrees in large extent to the values we found for BMR of mouse determined by indirect calorimetry given in Table 1.
3. This pilot experiment (Figure 3) proves that a single mouse supplied with food and water can survive for at least 23 hours (Figure 3: 5-18 hours + precondition period of 5 hours) in the 1 liter stainless steel vessel, if supplied with a continuously air flow of 1 liter per minute. The selected temperature of room and calorimetric set up was 21°C, which is rather low because according to some authors the thermo-neutral zone for murents is ≈28-32°C.
4. Not clearly visible in Figure 1 (but rough data in our .PRN file) demonstrate that the electrical calibration and the calculated AUC result in a sensitivity coefficient of 144 μV/mW.
When this value is inserted in the software before the start of the pilot experiment the calibration procedures at the start and at the end of the experiment correspond to ≈100 mW.
We calculated in the M and M two different time constants for (in the past as an aquatic flow through calorimeter for mainly fish [21-28] the physical medium water The time constant τwater of the developed 1 litre differential flow-through calorimeter (Sétaram GF 108) was estimated for the medium water to be 30.48 minutes [22]. With this time constant we could perform deconvolution techniques to correct afterwards a direct biological calorimetric dataset for the lag-time of the system (heat-transfer from water to stainless steel in combination could in the past desmeared the datasets of goldfish (Carrassius auratus L) under metabolic depression during anoxia and describe the dynamic response of this “hibernation” process which was estimated to take place on a time scale of ≈ 20 minutes [22]. An example of an electrical calibration in the medium water with a time constant of 30.48 minutes is given in Figure 4.

Figure 4: An example of an electrical calibration in the medium water with a time constant τwater of 30.48 minutes with the original signal, the deconvoluted signal and the per 10 points (sample frequency once a minute) smoothed signal.
However our time constant (τair ) we calculated for the physical medium air is only 0.019605 min. We desmeared, using deconvolution techniques, the calorimetric dataset of an electrical calibration in air with a resistor of 1000 Ω using the time constant τair of 0.019605 min. and in addition also calculated the smoothed curve based on the method of Hand and Gnaiger [29]. This implies that we take per sample point with a sample frequency of 1 sample per minute the mean over an time interval of 10 minutes (10 sample points), all along the curve. The three graphs we obtained are depicted in Figure 5.

Figure 5: Three graphs are depicted in this figure in a calorimetric vessel filled with air and refreshed with a continuous air flow of 1 liter per minute (through both vessels: twin detection): A). The original dataset after electrical calibration during a period of 6 hours with a resistor of 1000 Ω mounted in the Mesurement calorimetric vessel; B). The deconvoluted dataset corrected for the time constant τair of 0.019605 min; C). The deconvoluted and smoothed dataset according to 29). Air has probably a low specific heat capacity and is therefore very sensitive to a temperature change so there is nearly no lag-time.
Central message of this research manuscript was that there are several reasons [19], that indirect calorimetry doesn’t work for nowadays determination of BMR and energy expenditure of mouse models (wildtype-, High-Fat diet induced obese or transgenic-) based on a) Methodological grounds, b) Physiological information regarding the “created” transgenic mouse models, c) Genotypical variations can be critically reviewed if the indirect calorimetric method (based on the conversion factor RQ) is a correct approach to calculate the heat equivalent of energy output, routine metabolic rate (RMR) or BMR. d) Phenotypical variations like gender, ageing, housing conditions. We therefore performed a pilot experiment with an existing (from origin aquatic) twin detection flow-through Setaram calorimeter with three objectives:
1. Are we able in the (physical) medium air to measure the heat production of a single mouse? [Note that this from origin aquatic flowthrough calorimeter was specific developed for these conditions which implies: a) Heat-transfer from water to stainless steel in combination with mainly. b) The large specific heat capacity of the medium water with as consequence being rather insensitive to a temperature change].
2. Are we able to perform long lasting experiments (≈ 12 hours or longer) with one single mouse in this one liter stainless steel calorimetric vessel and -to prevent suffocation of the animal- lead continuous a constant air flow of 1 liter per minute through the two vessels (twin detection) which would result in a continuous heat loss but also wouldn’t affect the stability of the baseline;
3. By electrical calibration using a resistor of 1000 Ω mounted in the experimental vessel would give (via a calibration factor expressed in μV/mW quantify in a reliable way the heat production of the mouse in order to measure the BMR. In the Results paragraph we gave our findings and we can conclude based on this pilot that this flowthrough (air) twin detection Seteram micro-calorimeter with stainless steel vessels of 1 liter can be used as a mouse calorimeter to measure the energy expenditure and BMR in murent models. This is urgently needed as we will emphasize with the following arguments:
• The answer is YES based on characteristics of the calorimetric set-up under air (Table 1) with during a direct calorimetric experiment of 33 hours a baseline stability of 1.1124%, Δ-(baseline) 1.51 mW which is in comparison to the RMR (Table 3) only an error in the range of 0.0014%. The minimal level (BMR) over a time interval of one minute of this pilot we found for a single mouse in our air-based Seteram twin detection micro-calorimeter corresponded to ≈ 411.35 mW/ 40.1 g (≈10.26 mW/g). This agrees in large extent to the values we found for BMR of mouse determined by indirect calorimetry given in Table 1. Four different authors [30-35] gave in general a uniform pattern that the by indirect calorimetry determined BMR of mouse is around ≈ of around 10 mW/g in the temperature range of 25-30°C (Table 1, third column details experiment). It has to be remarked that because this is the first direct calorimetric study at the BMR of mouse we can only compare our BMR value with these of indirect calorimetric studies. Hopefully these studies were not performed with “metabollically abnormal” animals which was our premises for our arguments that direct calorimetry has to come back as measurement tool to determine the BMR of mouse models [19,20].
• Future studies for e.g. with starvation animals (but with unlimited supply of water) will with mouse probably last a maximum of 48 hours because in previous experiments with localized in vivo MRI-imaging with a C57/BL6 mouse model studying the process hepatic steatosis and its lipid composition in the selected liver voxel [31] revealed that 48 hours is at the physical limits a mouse can handle. This statement was based on observations we performed during that experiment like: continuously shivering, curled position of the body, no activity patterns and it seems a continuously state of “metabolic arrest”, unable to handle to anaesthetize the animals with 4% isoflurane in air (50%) and O2 (50%) and anaesthetics was maintained with ~1.5% isoflurane to put them in the “mouse-probe” for this special for mouse developed in vivo 1H MRS localized 9.4 T spectrometer with vertical bore and imaging system (Bruker Biospin, Karlsruhe, Germany) [31]. So in principle, when fed, the amount of available food and water enclosed in the 1 liter calorimetric vessel determine the time period of a direct calorimetric experiment. Furthermore in Figure 1 we see over the time-period 20-29 hours a flat stable baseline.
• The answer is yes seeing ‘Results’ inter alia Figures 2 and 3 and Table 3. In addition regarding the time constant of this calorimeter in the physical medium water (previous experiments) [21-28], and now in air. We calculated in the M and M of this research manuscript two different time constants which values clearly demonstrate the totally (physically determined) different environmental conditions this Seteram calorimeter now how has to operate. These time constants were an aquatic time-constant τwater of 30.48 min in comparison to a time-constant in air of τair or = 0.019605 min. Because the τair was so small deconvolution techniques (earlier successfully applied in water [22]), correcting for the lag time of the system had no effect.
In general, direct calorimetric studies at the energy expenditure of mouse models are at this moment urgently needed for the following scientific reasons.
Methodology
By far the most difficult aspect of indirect calorimetry is measurement of O2. Moreover, the abundance of O2 (20.95%) relative to CO2 (0.03%) in ambient conditions dictates that measurement errors of O2 have greater implications on calculations of RMR or BMR and RQ. Because compressed air is not feasible for use with animals in long-term experiments, changes in ambient conditions are nearly unavoidable Jensen et al. [16].
Physiological
Relying on metabolically abnormal animals in transgenic mouse models or pharmacologically-induced alterations in mouse strains: The major disadvantages of indirect calorimetry (up to this moment only worldwide applied in (also genetically engineered mouse models) are the following. Genetically engineered mouse models for contemporary Obesity and/or Type-2 diabetes (DM2) will in most cases deliver heavily metabolically abnormal animals. This may probably reflect itself also at the cellular level e.g. in case of genetically induced i). Changes at the mitochondria of a transgenic mouse model, ii) Or in case of a shift in the by the “metabolically machinery” preferred substrate e.g. lipids instead of carbohydrates (or a diminished “metabolic flexibility” ≈ the capacity of an organism to switch between substrates, mainly glucose vs. FFA); iii) But also at the level of the organism in Obese and Type-2 diabetic animals the gut microbiotics (≈prokaryotic and for 99% anaerobic) outnumber eukaryotic cells in the body by ≈10-fold and; iv) This raises important questions about transforming aerobic gas exchange into Heat Production based on a conversion factor the Respiratory-Quotient (RQ-value: CO2/O2) and if this is allowed in genetically engineered mouse strains and will result in valuable values of the energy expenditure of this genetically engineered animal in comparison to “traditional” wild-bred strains which rely nearly totally on an aerobic metabolism [19,20]. Direct calorimetry, however, can accurately quantify heat production (HP) and thus RMR or BMR in both metabolically normal and abnormal states, whereas respirometric indirect calorimetry relies on important assumptions that apparently have never been tested in animals with genetic or pharmacologicallyinduced alterations that dysregulate metabolic fuel partitioning and storage (≈ maintain “Energy Homeostasis”) in metabolically normal and abnormal states. An example is the genetically engineered mouse models for contemporary Obesity and/or Type-2 diabetes (DM2) [36]. Which rely heavily on metabolically abnormal animals? Recent data implicating individual and group variation in the gut microbiome in obesity and diabetes raise important questions about transforming aerobic gas exchange into HP because 99% of gut bacteria are anaerobic and they outnumber eukaryotic cells in the body by ∼10- fold [19]. Recent credible work in non-standard laboratory animals documents substantial errors in respirometry-based estimates of HP of pharmacological and genetic murent models of Obesity and Type-2 Diabetes [19]. Recent studies demonstrated that diet-induced Obesity was linked to changes in the gut microbial ecology, resulting in an increased capacity of the distal gut (colon) microbiota to promote host adiposity due to an increased energy harvest from the diet. Food ingredients like fructose improve metabolic alterations associated with Obesity, including dyslipidemia, impaired gut permeability, endotoxemia, inflammation due to a changed innate immune system resulting in the pathogenesis of metabolic disorders including DM2 in increasing amounts among the young e.g. children at an age of 7 years are already obese!!. Transplantation experiments of caecal microbiota from lean and obese mice in the gut of germ-free mice definitely demonstrated that altered gut microbiota composition is a cause and not a consequence of obesity or altered dietary habits [37].
Genotypical variation
A typical example is the SMA1 mice which carry a missense mutation in the growth hormone gene that leads to semi-dominant dwarfism and obesity. The thermal properties of these SMA1 mice were examined by indirect calorimetry and this strain has proportionally a lower BMR and RMR and a higher lower critical temperature [17].
Phenotypical variations
i) Gender, ii) Ageing, iii) Housing conditions like group or solitaire, iv) Exposure to an exercise protocol of e.g. obese mouse strains followed in time.
Gender
Sexes differences in RMR of mouse models have also been observed in mouse models at different temperatures (5˚, 15˚, 25˚, 30˚C) based on indirect calorimetry by Selman et al. [33] giving a RMR for males of 10.30 Mw/g and a RMR for females of 12.03 mW/g. But also clear differences between females and males C57BL/6 mouse were observed in the study of [38] after voluntary running on an angled running track during an exercise protocol of 36 days (see iv.) resulting in a higher activity and better performance of the females.
Ageing
It may be clear that the knock-in-mice model of Trifunovic [14] with an increase in somatic mtDNA mutations, which are associated with reduced lifespan of the organism, also in time probably, will result in a changing BMR. Also the knock-out mouse strain for the senescence marker protein-30 (SPM-30) of [15], for which it is suggested that SPM-30 the SPM-30 gene has an anti-apoptotic function, probably also may result during time course in a changing BMR.
Groups or solitaire housing
Male mouse are strongly territory bounded and housed in a group have a strongly hierarchy with social dominance over each other, which can probably affect hormones like the stress hormone in mouse: corticosterone, in this way strongly affecting the BMR of each individual animal.
Exercise
Exposure to an exercise protocol of 36 days of a C57Bl/6 mouse model, both males and females running voluntary on an angled running track, gave an improvement in time of certain physical parameters including parameters for exercise performance during the test period but also demonstrated a clear differences between sexes with a better exercise performance and higher activity level of the females. While the time of daily running expressed in km was for males at the start of the experiment 2 km and increased towards 8 km after 36 days these numbers for females were 5 km => 15 km after 36 days. Also the time spent running over that 36 days periods was for males initially 2 h and increased towards 3 h while this was for females 3.9 h going towards 6 h [38,39]. The effect of exercise in obese mouse strains will have via an altered RQ value an effect on the BMR. Also a clear day-night rhythm was observed with the highest activity during night. In addition it has been reported that voluntary exercise improved insulin sensitivity and adipose tissue inflammation in diet induced obese mice [40-42].
In conclusion we reviewed the literature regarding mouse models and came to the conclusions coinciding the reviews of [19] that measuring the energy expenditure and consequently the BMR by direct calorimetry is nowadays a prerequisite in modern Medical Bioscience and Life-sciences with (transgenic and/or obesity induced) mouse models. This pilot experiment has demonstrated that this is technical feasible and reasonable. In this study we determined successfully for the first time the BMR of a common mouse by direct calorimetry ≈ 10.26 mW/g bodyweight.
Although Bio-calorimetric research is more and more switching towards the cell-, (sub)cellular components like organelles as mitochondria, bio-molecular interactions (to give some examples: DNA-protein interactions, ribonuclease-ligand screening, the effect of growth-factors on human fibroblasts) [41]. Despite this “tendency to the small” this study clearly demonstrates that direct calorimetric studies at the level of the whole animal (organism) are urgently needed in Biomedical Research and Life Sciences. For the Biomedical Physiologist tremendous opportunities are offered with: a) The new genetically engineered mouse models; b) “New state of the art techniques” to study via mouse models the pathogenesis of human diseases like Obesity and metabolic related disorders like Type-2 Diabetes or more fundamental Life Sciences related processes like ‘ageing’. Regarding these “new state of the art techniques”, it is our intention to extend this already unique sophisticated sensitive Setaram twin-detection direct calorimetric system (with also unique a vessel with a size of 1 litre), with gas-analysing equipment for simultaneously indirect calorimetry. Furthermore, the special for mice developed LABORAS animal behaviour and motion analysis system (company METRIS, Hoofddorp) which based on tremendous small vibrations caused by the different motion patterns caused by varying behaviour components of the mouse (like: grooming, eating, drinking, resting, rearing, locomotion and climbing), “recognizes” these different motion patterns. But also the SONOTRACK ultrasonic vocalization system of the same company which based on vocalizations provide an index of the emotional state and affective state of the animal in the 1 litre calorimetric vessel. In addition, at the same time and synchronous with the via direct calorimetry obtained energy expenditure data, we intend to build in the peritoneal cavity of the mouse small invasive radio-telemetry transmitters of ≈3.3 g (type: TA 10ETA-F20-L20) to deliver extra physiological information like: i) Body core temperature, ii) Heart-rate frequency (HRF) and produce an iii) Electrocardiogram (ECG) [Data Sciences International (DSI), St. Paul, USA]. The combination of all these “state of the art techniques” like: a) LABORAS: to correct for motion and determine the “real” BMR); b) SONOTRACK: to determine based on ultrasonic vocalization the emotional state and affective state of the animal e.g. is the animal stressed?; c) Invasive radio-telemetry transmitters for possibly fundamental physiological processes like e.g.: a) The relation to the hypothalamic regulated body-core temperature and by direct calorimetry measured energy expenditure; b) The relation between emotional state and affective state of the animal (SONOTRACK), HRF, body-core temperature (DSI) and energy expenditure (Setaram indirect and direct-calorimetry) at the BMR-level (LABORAS) opens a broad range of new opportunities to elucidate questions from Biomedical-Science regarding the pathogenesis of some diseases and Life-Science related processes like ageing. Also in order to combat obesity the rather new research area of a specially treated food, vitamin, mineral, herb, etc., that you eat or drink in order to improve your health can be investigated using this new research tool (DMC) [42].
Vincent van Ginneken kindly acknowledge the Staff of the Biology Department, Leiden University, the Netherlands for unrestricted use of the Setaram twin-detection calorimetric set up which will be helpful to build up an own Biomedical line of research for our Blue Green Technologies Foundation www. bluegreentechnologies.nl