Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Review Article - (2014) Volume 3, Issue 2
Malignant carcinoma is one of the most threatening diseases to our human being’s life and health. However, chemotherapy, as one of the principal modes of treatment, especially for metastatic cancers, has been blocked by drug resistance, which contributing to disease progression and leading to a high mortality rate. Thus, research on both mechanisms and resolutions to chemo resistance seems extremely indispensable and urgent. MicroRNAs (miRNAs), the recently emerging class of novel molecules, have been reported to have close correlation with cancer. They act as oncogene or tumor suppressor gene, and growing evidence indicates that the desregulation of miRNAs involves in drug resistance. Alteration of miRNA’s expression leads to obviously different responsiveness to anticancer drugs. It implicates that miRNA may be a promising therapeutic target for improvement of chemoresistance, and it is inevitable that the settlement of drug resistance will be a giant leap and milestone in treatment of malignance. So in this review, we’ll elaborate the potential and promising role miRNAs playing in clinical treatment and progress.
<Keywords: miRNA; Cancer; Chemosensitivity
MiRNAs are evolutionary conserved endogenous non-coding RNAs with approximately 22 nucleotides in length, that generally negatively regulate gene expression by perfect (always appearing in plants) or imperfect (appearing in animals) complementary basepairing on the 3’-untranslated region (3’-UTR) of the sequences of target mRNAs, resulting in their cleavage or translational repression [1]. It was reported that miRNA can also bind to the 5’UTR of target genes to repress the target mRNAs and can induce translation upregulation of target mRNAs in specific conditions (such as serumstarved conditions) [2]. Alike to protein-coding RNA, miRNAs are also transcribed by RNA polymerase II enzyme inside the nucleus, producing primary-miRNA (pri-miRNA), more than with a cap structure at the 5’ end and a poly-adenylated at the 3’ end, specifically with characteristic hairpin-shaped stem-loop structures. With the cleavage of Drosha, the mature-miRNA molecule pre-miRNA is formed, then pumping out into cytoplasm by Exportin 5, being integrated into RNA-Induced Silencing Complex (RISC), mature and participate into cleavage or translational repression of mRNA (Figure 1) [3]. Since the discovery of the two initial miRNAs lin-4 and let-7, which regulate the timing of stem-cell proliferation and differentiation in nematode Caenorhabditis elegans, [4-6] and let-7 was subsequently found as the first known human miRNA [7], to date thousands of miRNAs have been identified in a wide range of plants and animals. Investigations have identified that miRNAs are implicated in critical functions across various biological processes in cells as diverse as developmental timing, cell development proliferation and death, metabolism like fat metabolism, hematopoiesis, patterning of the nervous system, tissue morphogenesis and even leaf development and floral patterning in plants [8-10]. Over and above the referring functions about development proliferation differentiation and apoptosis, some miRNAs are always elucidated to involved in pathological conditions, for instance, cardiovascular diseases [11-15], diabetes [16-18], viral infection [19,20], and cancer.
Chemotherapy, as one of the primary treatments especially for advanced cancers, is blocked by drug resistance. Research on drug resistance in cancer has focused on cellular resistance due to both the specific nature and genetic background of the cancer cell itself, and the genetic or epigenetic changes that follow toxic chemotherapy, namely intrinsic resistance and acquired resistance [21]. At present, the anticancer drug resistance is considered to be a multifactorial phenomenon involving three major mechanisms, firstly, decreased uptake of water-soluble drugs, such as folate antagonists, nucleoside analogues and cisplatin, which require transporters to enter cells; secondly, various changes in cells that affect the capacity of cytotoxic drugs to kill cells, including alterations in cell cycle, increased repair of DNA damage, reduced apoptosis and altered metabolism of drugs, changes in glutathione transferase expression and topoisomerase II and etc.; and thirdly, increased energy-dependent efflux of hydrophobic cytotoxic drugs that can easily enter the cells by diffusion through the plasma membrane leading to diminish the ability to kill cancer cell [22,23] (Figure 1).
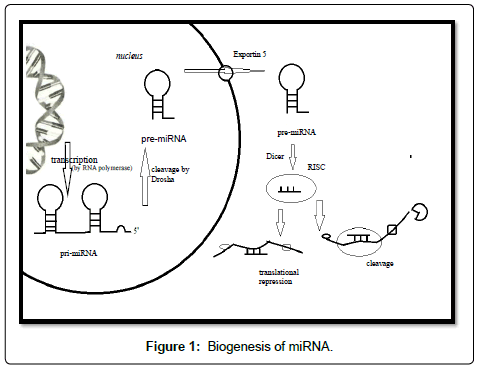
Figure 1: Biogenesis of miRNA.
Relationship between miRNAs and cancer
MiRNA was firstly demonstrated linking with cancer in 2002 [24], as is said the deletion and down-regulation of miR-15 and miR-16 was always found in Chronic Lymphocytic Leukemia (CLL). From then on, mounting investigations and evidences demonstrated the robustly implication between miRNAs and cancer, varying in tumorigenesis, progression (invasion and metastasis), diagnosis, treatment and prognosis [25,26]. To date, over 1000 miRNAs have been reported in humans, and it is also proved that 98 of 186 (52.5%) of miRNAs genes are in cancer-associated genomic regions or in fragile sites [27]. A single miRNA can always impact hundreds of targets, and single target can always be affected by multiple miRNAs [28,29]. However, miR-155 firstly showed us that the deregulation of a single miRNA gene can also lead to cancer (CLL) [25].
MiRNAs function as oncogene or tumor suppressor gene during the tumorigenesis and progression as is reported, repressing the translation of their targeted genes or directly inducing their degradation. Since the negative regulation of targeted genes expression, once the expression of miRNA itself changed, aberrant gene expression would be emerging. Those being deemed to play a crucial role in the initiation and progression of human cancer are designated as oncogenic miRNAs (oncomiRs), with over expression level in tumor cells, such as miR21, miR372/373, miR155, miR221, miR191 and miR17-92 etc. While let-7, miR15, miR16, miR98, miR127, miR143 always act as tumor suppressor gene, with low expression level in tumor cells [21,22,30-33]. Examples in Table 1 may give us better understanding.
| miRNA | Human cancer | Targets | Researchers | References |
|---|---|---|---|---|
| miR-21↑ | gastric cancer | PTEN | Zhang BG | [28] |
| miR-15a miR-16↓ | prostate cancer | Bcl-2, CCND1 and WN T3A | De´sire´eBonci | [24] |
| miR-21 | prostate cancer | MARCKS | Tao Li | [14] |
| miR-27a↓ | gastric cancer | P-gp↓p21↑ | Zhao X | [37] |
| miR-520b↓ | hepatocellular carcinoma | MEKK2↑ Cyclin D1↑ | WY Zhang | [63] |
| miR-125b | breast cancer | ENPEP, CK2-α, CCNJ, MEGF9 | Feliciano A, | [75] |
Table 1: Examples of tumor suppressors.
Since resistance of cancer cells to chemotherapy continues to be a major clinical obstacle to the successful treatment of cancer, there have been accumulating researches evaluating the mechanisms of resistance and the biological factors involved, they all put forward that miRNAs could regulate factors to affect chemotherapy efficacy.
Anti-oncogenes or oncogenes
Anti-oncogenes and precursor of oncogenes, functioning in cell progress as well as normal genes, are well known to play a critical role in cancers [34-39]. On the way exploring effective gene therapeutics about malignant tumors, Evidences demonstrate close relations between miRNAs and them. As the target of miRNAs, they would accompany with the desregulation of miRNAs. The abnormal expression of antioncogene and oncogene are more than associated with tumorigenesis, there is also association with chemotherapy, bridging by miRNA. There are some examples lists in Table 2.
| miRNA | Human cancer | Drugs | Targets | Researchers | References |
|---|---|---|---|---|---|
| miR-21↑ | breast cancer | ADR | PTEN↓ | Wang ZX | [44] |
| miR-122↑ | acutaneous T-cell lymphoma | _ | Akt↑ p53↓ | Nalls D | [56] |
| miR-34a↑ | medulloblastoma | mitomycin C cisplatin | MAGE-A↓ p53↑ | Shi L | [57] |
| miR-19↑ | Breast cancer | PTEN↓ | Liang Z | [65] | |
| miR-130a↑ | Non-small cell lung cancer | _ | p27(kip1), PTEN and TIMP3↓ | Acunzo M | [66] |
| miR-21↑ | Breast cancer | trastuzumab | PTEN↓ | Gong C | [67] |
| miR-19a | Bladder cancer | arsenic trioxide | PTEN | Cao Y | [68] |
| miR-92b | Non-small cell lung cancer | CDDP | PTEN | Li Y | [76] |
Table 2: Example of Anti-oncogenes or oncogenes.
Multidrug resistance (MDR), simultaneous resistance to different drugs with a variety of chemical structures and diverse chemotherapeutic functions, which is most commonly encountered in the laboratory, is always mediated by an altered expression family of energy dependent transporters that leads to drug efflux from cancer cells. There are three commonly proteins known as P-glycoprotein (P-gp; from MDR1 and ABCB1 genes), MDR-associated protein (MRP1; from ABCC1 gene), and breast cancer-resistant protein (BCRP; from ABCG2 gene), facilitating drug transport across the cell membrane [40].
P-gp, an efflux pump, generating from the MDR1 gene in the human, was one of the first members described of a large family of ATP-dependent transporters known as the ATP-Binding Cassette (ABC) family. It is low-level expressed in normal cells, involved not only in efflux of drugs but in moving nutrients and other biologically important molecules into, out of, and across plasma membranes and intracellular membranes in cells, but widely expressed in many human cancers, being conferred drug resistance. Previous review has given an exhaustive summary of interrelationship between ABC transporters from ABC subfamilies and chemotherapeutic agents in vitro as well as in clinical trials [41-49]. Current ongoing researches throw further light on the correlations. Some examples have given in Table 3.
| miRNA | Human cancer | Drugs | Researchers | References |
|---|---|---|---|---|
| MiR-200c↑ | breast cancer | epirubicin | Chen J | [35] |
| miR-122↑ | HCC | ADM or VCR | XU Y | [36] |
| miR-27a↓ | ovarian cancer | paclitaxel | Li Z | [38] |
| MiR-138↑ | leukemia | VCR | Zhao X | [39] |
| miR-331-5p↓miR-27a↓ | leukaemia | doxorubicin | Feng DD | [40] |
| miR-27a↓ | hepatocellular carcinoma | 5-fluorouracil | Chen Z | [8] |
Table 3: Some examples of current ongoing researches throw further light on the correlations.
Cancer stem cells
Stem cells are defined as cells that have the ability to perpetuate themselves through self-renewal and to generate mature cells of a particular tissue through differentiation [50]. Dysregulation of stem cell self-renewal is a requirement for the initiation and formation of cancer, besides, it is also a very likely cause of resistance to current cancer treatments, as well as relapse in cancer patients. As is firstly identified in leukemia [51], Cancer Stem Cells (CSC) are defined as a small subpopulation of cells in a variety of tumors that are capable of unlimited asymmetric self-renewal, differentiation, generating the heterogeneous lineages of tumor cells and initiating or maintaining the tumor [52]. S. D. Hatfield and his colleagues have found that MirRNAs play important roles on stem cell continuous division by downregulating Dap, a negative regulator in transition between the G1 and S phases of the cell cycle [53]. Recent evidences have also paid attention to “niche”, a kind of local tissue microenvironment that maintains and regulates stem cells [54]. It is tempting to speculate that miRNAs could play an important role in keeping up the microenvironment. As is referred above, CSC is one of the possible cause of relapse and resistance to cancer treatment, cancer stem cells may be highly resistant to radiation and chemotherapy; therefore, the development of more effective therapies for cancer requires effective targeting of this cell population to be maximally effective, cancer therapy must also be directed against both the resting cancer stem cells and the proliferating cancer cells [55]. This may be possible if specific stem cell signals are inhibited using molecular therapy, while at the same time attacking proliferating cells by conventional therapies [56-59]. Some list in Table 4.
| mirna | Cancers | Drugs | Researchers | References |
|---|---|---|---|---|
| miR-21↑ | Glioblastoma | Temozolomide | Zhang S | [48] |
| miR-451 | colon carcinoma | irinotecan | Bitarte N | [49] |
| miR-34a↓ | pancreatic cancer | 5-Aza-dC Vorinostat | Nalls D | [56] |
| miR-238a | colorectal cancer | Xu XT | [69] | |
| miR-128 | Breast cancer | doxorubicin | Zhu Y | [70] |
Table 4: List of Cancer stem cells.
Apoptosis-associated genes
Apoptosis, known as programmed cell death, is an important physiological process, which occurs in cells during development and normal cellular processes. As the major mechanism of chemotherapy, which induced by chemotherapeutic agents, joins into most of the war fighting with cancer, evidence from increasing number of investigations on this has shown that miRNA may bridge them together. In fact, mechanisms on chemotherapy, we have mentioned above, are always associated with apoptosis. The Table 5 lists some examples.
| miRNA | Human cancer | Drugs | Targets or signaling pathway | Researchers | References |
|---|---|---|---|---|---|
| miR-122↑ | hepatoblastoma | 5-FU | Bcl-2 Bcl-XL | Yin J | [42] |
| mir-15↑ mir-16↑ |
gastric cancer | VCR | Bcl-2 | Xia L | [41] |
| miR-21↑ | glioblastoma | Temozolomide | Bax/Bcl-2 | Shi L | [57] |
| miR-181a miR-630 | non–small cell lung cancer | cisplatin | mitochondrial/ postmitochondrial steps of apoptosis | Lorenzo Galluzzi | [77] |
Table 5: List of circulating miRNAs
MiRNAs are ubiquitous in most of the body fluid types. A survey Weber etc. conducted, about the miRNA distribution in human body fluids, shows the presence and distinct compositions in different fluid types and the association with various physiopathological conditions including cancer [60]. Previous research shows that miRNA expression profile reflects the developmental lineage and differentiation state of tumors [61]. It suggests that miRNAs may be used as biomarkers for diagnosis of cancer and prediction of prognosis for clinic, as is mentioned above, that the quantity of miRNAs even presence or not is associated with the tumorigenesis as well as outcomes of therapy on cancer. Besides, it may be also possible evidence for individual therapy. To date, much investigations closely concern about circulating miRNAs. Some are listed in Table 6. Although several challenges remain to be addressed, the exciting prospective of using miRNAs in human body fluids as powerful and non-invasive cancer biomarkers, has the necessity to research for the use for the diagnosis and prognosis of cancer diseases. Since the non-specificity of ectopic expression of miRNA in tumors, a pan-human microRNA, and high density microarray was introduce to detect and analyze the expression pattern of much miRNAs, in order of discriminating cancer patient from normal ones (Table 6) [62-74].
| Mirna | Cancer | References |
|---|---|---|
| MiR-125b↑ miR-181a↓ miR-205, miR-342; miR-21↑ |
Breast Cancer | [13,70,74] |
| miR-21↑ miR-210, miR-31 and miR-182↑ miR-126, miR-145↓ |
Lung cancer | [62,73] |
| miR-15b, miR-23a, miR-133a, miR-150, miR-197, miR-497, miR-548b-5p↓ | malignant astrocytomas | [72] |
Table 6: List of pan-human microRNA
Complex and tendency of relapse of cancers contribute to poor efforts of chemotherapy and formation of some chemoresistance. Researches on therapies of cancers are multifarious, and more investigations should be conduct. Since miRNA discovered, much studies have been placed onto the regulation it conducted, as well as the association with diseases like cancer. A growing number of investigations and evidence have pointed out the essential role it plays in cancers’ tumorigenesis, progression (invasion and metastasis), diagnosis, treatment and prognosis. For the reason that a single miRNA can always impact hundreds of targets, and single target can always be affected by multiple miRNAs, it is obvious that of ectopic expression of miRNA in tumors is non-specific. Thus, it is necessary to identify the exquisite associations between miRNAs and cancers. A high density microarray of miRNAs may be useful for cancers vary in diagnosis, individual treatments, and estimation of prognosis. And treatments on the level of miRNAs, may be a solution of chemoresistance and relapse, considering its network with anti-oncogenes or oncogenes, MDR, CSCs, apoptosis-associated genes.