Cell & Developmental Biology
Open Access
ISSN: 2168-9296
ISSN: 2168-9296
Review Article - (2012) Volume 1, Issue 1
The larval transfer hypothesis states that larvae originated as adults in other taxa and their genomes were transferred by hybridization. It contests the view that larvae and corresponding adults evolved from common ancestors. The present paper reviews the life histories of chordates, and it interprets them in terms of the larval transfer hypothesis. It is the first paper to apply the hypothesis to craniates. I claim that the larvae of tunicates were acquired from adult larvaceans, the larvae of lampreys from adult cephalochordates, the larvae of lungfishes from adult craniate tadpoles, and the larvae of ray-finned fishes from other ray-finned fishes in different families. The occurrence of larvae in some fishes and their absence in others is correlated with reproductive behavior. Adult amphibians evolved from adult fishes, but larval amphibians did not evolve from either adult or larval fishes. I submit that [1] early amphibians had no larvae and that several families of urodeles and one subfamily of anurans have retained direct development, [2] the tadpole larvae of anurans and urodeles were acquired separately from different Mesozoic adult tadpoles, and [3] the post-tadpole larvae of salamanders were acquired from adults of other urodeles. Reptiles, birds and mammals probably evolved from amphibians that never acquired larvae. I claim that the larval transfer hypothesis offers the most credible, and in some cases the only, explanation of many anomalies in the development of chordates. Larval transfer is a type of hybridogenesis, which is discussed in relation to other types of evolution, including symbiogenesis and Darwinian ‘descent with modification’.
<Keywords: Amphibian; Amphioxus; Axolotl; Cyclostome; Fish; Jaw; Tadpole; Tunicate
The larval transfer hypothesis states that basic forms of all larvae originated as adults in other taxa and their genomes were transferred by hybridization [1]. I suggest that the first larva was a sexual hybrid between two distantly related species. Sperm of one species fertilized eggs of the other, which hatched as larvae resembling one parent, then metamorphosed into juveniles (small adults) resembling the other. All descendants of this cross were animals with larvae, in which one animal form followed another: they were sequential chimeras [2]. Corollaries of this hypothesis are that metamorphosis represents a change in taxon during development, larvae are later additions to the evolutionary history of species with larvae, and they do not represent evolutionary ancestors of such species. My 2003 book included cases ascribed to larval transfer from most phyla with larvae, but all the examples were drawn from invertebrates [1]. The present paper discusses the larvae of chordates, including vertebrates. It overlaps with previous publications on the tadpole larvae of urochordates [1-5], but it is my first paper to consider the origins of larvae of craniates and vertebrates. It presents no new observations on chordate development, but it views old observations from a new perspective. It interprets chordate development in terms of larval transfer. Firstly, however, I briefly review historical views on the status and phylogeny of larvae.
Darwin was convinced that all evolution is gradual, and he assumed that each larva and its corresponding adult had gradually evolved from a common ancestor [6]. Haeckel accepted the common ancestor theory, and he put forward his own ‘biogenetic law’ that ‘ontogeny recapitulates phylogeny’ [7]. He claimed that larvae represent ancestral adults and that major evolutionary changes are confined to adults. Balfour distinguished between ‘primary larvae’, which “are more or less modified ancestral forms”, and ‘secondary larvae’, which “have become introduced into the ontogeny of species” [8]. He regarded virtually all extant larvae as secondary, but he was unclear about the sources of introduced larvae. Garstang ignored Balfour’s work [9]. He updated Haeckel’s biogenetic law by proposing that modern larvae represent ancestral larvae rather than ancestral adults, and that, contrary to Haeckel, ontogeny creates phylogeny; it does not recapitulate it.
Haeckel’s and Garstang’s theories both assume that modern larvae represent ancestors, in some form. They prompt two questions: do animals without larvae have no evolutionary history, and, in animals with one larval form, what has happened to the other forms through which the animal must have evolved? My proposals on larval transfer were conceived independently of Balfour’s on introduced larvae, but they are similar. Balfour died in a climbing accident at the age of 30, in 1882, within a year of the publication of his seminal views on larvae, which have been largely ignored until recently.
Garstang drew attention to cases in which the larvae of one group of animals resemble the adults of another group, and he proposed that such adults are descendants of ‘persistent larvae’: animals that had matured in the larval state [9-11]. I regard such adults as surviving relatives of animals that were the sources of larvae.
I claim that ancestral chordates developed directly. In the present paper, I accept the current consensus on the evolution of adult chordates, I summarize development in taxa that contain species with larvae, I argue that such larvae were later additions to life histories, and I show that each type of larva has a credible adult source. My conclusions are in accord with the larval transfer hypothesis, but not with any of the theories that assume that larvae and adults evolved from common ancestors.
Chordates
Chordates are animals with notochords or vertebrae. A notochord (literally back-chord) is a flexible unjointed cartilaginous tube. In vertebrates, the notochord is replaced by a jointed column of bony or cartilaginous vertebrae. In animals with notochords, a major nerve cord always runs parallel to the notochord, but the two are not attached. In vertebrates the corresponding nerve cord runs through the dorsal part of each vertebra, the neural arch. The brain of vertebrates, at the anterior end of the dorsal nerve, is enclosed in a cartilaginous or bony skull, making them craniates.
The phylum Chordata consists of the classes Urochordata (‘tail chordates’), Cephalochordata (‘head chordates’), and Vertebrata (with vertebrae). Many authorities include the order Cyclostomata or Agnatha (lampreys and hagfishes) in the Vertebrata. Cyclostomes are craniates that have notochords rather than vertebrae. Undisputed vertebrates are the orders Pisces (fishes), Amphibia (amphibians), Reptilia (reptiles), Aves (birds), and Mammalia (mammals). Larvae occur in the life histories of most urochordates, some cyclostomes, some fishes, and most amphibians. A newly hatched cephalochordate shows some asymmetry, but it is questionable whether the asymmetrical form is a true larva.
A tadpole is traditionally the larva of a frog or toad. In this paper, the term is also used in a broader sense to include any aquatic adult or larva that lacks functional paired fins or limbs and swims by undulations of a flattened muscular post-anal tail that encloses a notochord and a separate nerve cord. A craniate tadpole has a cartilaginous skull, and a gnathostomatous craniate tadpole has a jaw hinged to the skull.
Urochordates: All urochordates are marine, and most of them, at some stage in their lives, are acraniate tadpoles (as defined above). The group is made up of tunicates, many of which have tadpole larvae, and larvaceans, which are tadpoles throughout life. In tunicates, the main body is enclosed in a ‘tunic’ with two openings, the inhalent and exhalent siphons.
Ascidians (Ascidiacea) (Figure 1A) are tunicates that attach to rocks, wrecks and other solid surfaces. They contract when disturbed, squirting seawater through their siphons, and, because of this, they are often called sea squirts. The mouth leads to a large pharynx with many perforations. Ascidians have tadpole larvae that do not feed (Figure 1B), in which the organs of the juvenile (young adult) start to grow within the body of the swimming tadpole. As a result, the animal has, for a time, a developing ascidian brain and nervous system and a separate tadpole brain and nervous system. The tadpole settles within a few days, and the organs of the tadpole die. They make no contribution to the organs of the developing ascidian.
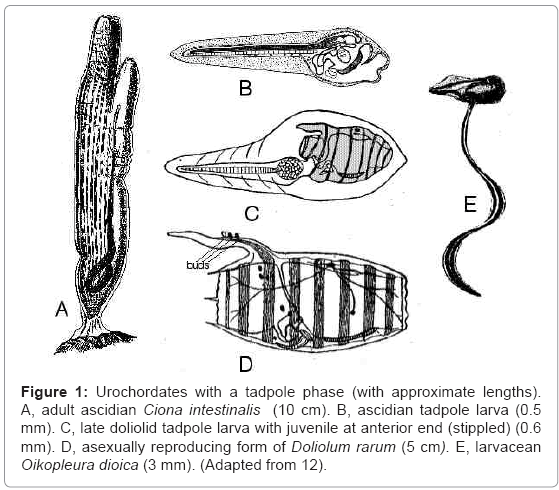
Figure 1: Urochordates with a tadpole phase (with approximate lengths). A, adult ascidian Ciona intestinalis (10 cm). B, ascidian tadpole larva (0.5 mm). C, late doliolid tadpole larva with juvenile at anterior end (stippled) (0.6 mm). D, asexually reproducing form of Doliolum rarum (5 cm). E, larvacean Oikopleura dioica (3 mm). (Adapted from 12).
The independence of the juvenile and the larva is more obvious in developing doliolids (Doliolida) (Figure 1C). Adult doliolids (Figure 1D) are barrel-shaped swimming tunicates with transverse muscles. The inhalent and exhalent siphons are at opposite ends, and contraction of the muscles provides a method of jet propulsion. In this group the juvenile grows as a swelling on the head of the tadpole larva (Figure 1C). In contrast to ascidians, the juvenile organs of doliolids are never near the larval organs, but doliolid and ascidian tadpoles are similar in other respects. Both are non-feeding, with no alimentary system. The juvenile doliolid will eventually break free while the tadpole is still swimming. The doliolid phase (Figure 1D) that follows the tadpole phase reproduces asexually by budding, and the buds migrate to the dorsal process at the rear end, known as the cadophore. Some of these buds develop into sexually reproducing doliolids, which lack cadophores.
Larvaceans (Larvacea or Appendicularia) (Figure 1E) are marine acraniate tadpoles that feed on small planktonic algae and bacteria, which they trap in a ‘house’ of secreted mucus. Larvaceans do not metamorphose. The tail is usually attached to the ventral side of the body. Its flexible skeleton is commonly called a notochord, but it is questionable whether the prefix ‘noto’ is appropriate. Larvaceans are placed in the same class as tunicates because of their resemblance to the tadpole larvae of ascidians and doliolids.
Salps (Salpida) (Figure 2A) and pyrosomes (Pyrosomida) (Figure 2B,C) are pelagic tunicates with no tadpole phase in their life histories. Like doliolids, they have inhalent and exhalent siphons at opposite ends of the body. The circular body muscles are incomplete ventrally in salps, but they nevertheless produce strong swimming movements. The corresponding muscles are feeble and restricted to the ends of the body in pyrosomes, which form large tubular colonies in which the inhalent siphons face outward and the exhalents face inward. Salps, like doliolids, have alternating sexual and asexual generations, but pyrosomes do not.The embryogeny of salps is similar to that of ascidians and doliolids, but that of pyrosomes is strikingly different [13,14].
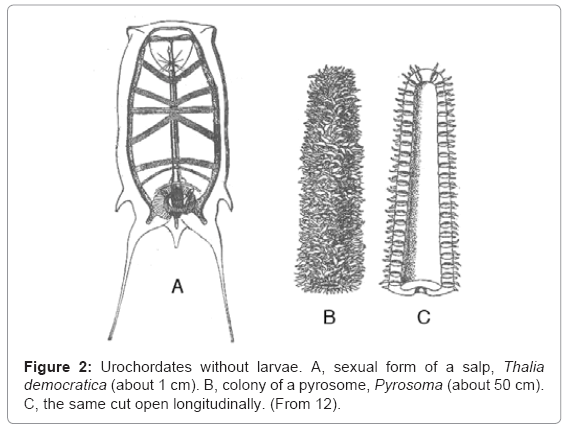
Figure 2: Urochordates without larvae. A, sexual form of a salp, Thalia democratica (about 1 cm). B, colony of a pyrosome, Pyrosoma (about 50 cm). C, the same cut open longitudinally. (From 12).
Remarks: The juvenile doliolid grows at the anterior end of the tadpole larva (Figure 1C), and it can break free while the larva is still swimming. This is an example of ‘overlapping metamorphosis’, in which the free-living juvenile and the larva co-exist for a time[1,15]. Comparable cases occur in echinoderms, polychaete worms, and nemerteans, and, in each case, the larva obviously does not ‘develop into’ the juvenile. As stated, “I know of no proffered explanation of how one animal, with one genome, might have evolved into two coexistent forms which are clearly neither twins nor clones. Such occurrences, however, are consistent with the suggestion that the basic forms of larvae were acquired by hybridization, and therefore two genomes are involved. The two coexistent body-forms represent those of the two animals that hybridized.” [15].
The alternation between sexually and asexually reproducing generations of doliolids may have resulted from hybridization between doliolids in different families, but I claim that the introduction of a tadpole phase into the life history of doliolids resulted from hybridization with a larvacean, with which doliolids are only remotely related [1,4].
Larvaceans are sufficiently like the non-feeding larvae of tunicates to convince several generations of zoologists that these two types of tadpoles must be related. Haeckel proposed that larvaceans were the original urochordates, and some larvaceans later evolved tunicate bodies [16]. Garstang suggested that larvaceans are descended from a persistent tunicate larva that did not metamorphose [10]. I theorize that larvaceans were originally unrelated to tunicates, but when one of them hybridized with a tunicate the result was a tunicate with tadpole larvae [1,4,5]. We now have draft genome sequences of both a larvacean, Oikopleura dioica [17], and an ascidian, Ciona intestinalis [18,19]. The 72 Mb genome of O. dioica is the smallest known in any chordate, and it shows genome architecture and organization patterns not seen in other animals [20]. The 160 Mb genome of C. intestinalis not only contains more than twice as many genes as O. dioica, but they are very differently arranged. I suggest that the small size and unique features of the genome of O. dioica are not consistent with Garstang’s hypothesis that larvaceans are descended from persistent tunicate larvae. I claimed that the genomic results are in accord with my proposal [5], but this was disputed by Hart and Grosberg who pointed out that C-values (weights of total DNA) paint a totally different picture [21]. C-values, however, vary enormously among animal species in general and even between congeneric species [22]. In most animals they contain a disproportionately large proportion (up to 99%) of non-coding DNA, and they bear no known relationship to the numbers of functional genes.
The similar embryogeny of direct-developing salps and indirectdeveloping ascidians and doliolids may be explained if former salps acquired tadpole larvae from the same source as ascidians and doliolids, but this larval form is suppressed in modern salps. Their embryos, however, show pre-tadpole features, just as the embryos of echinoderms that have secondarily lost their larval stage show bilateral features. I regard the direct development of salps as secondary, but I suggest that pyrosomes never acquired tadpole larvae, and their direct development is primary [1]. If so, the number of genes that code for proteins will be greater in salps and doliolids than in ascidians, because salps and doliolids not only hybridized with larvaceans to acquire tadpole genes but they also hybridized with closely related animals to acquire alternation of sexual and asexual generations. I predict that pyrosomes, with no hybridization events in their evolutionary history, will have many fewer genes that code for proteins than other tunicates.
Dominguez and Jefferies concluded that the Lower Cambrian genus Vetulicola “was almost certainly a tunicate and probably a stemgroup appendicularian.”[23]. Under my thesis, the Appendicularia (Larvacea) and the Tunicata were originally distantly related phyla, and the Ascidiacea, Doliolida and Salpida (but not the Pyrosomida) are descended from hybrids between former members of these phyla. These proposals cannot be shown in present methods of classification, but I urge geneticists to consider them in interpreting the genomes of species in these taxa. I question the assumption of several authors, including [20], that larvaceans are evolving rapidly.
Cephalochordates: The several species of lancelets that make up the genus Branchiostoma, better known as Amphioxus (Figure 3), are the only living cephalochordates. The name cephalochordate refers to the fact that the animal has a notochord in its head. Actually both the notochord and the dorsal nerve cord run practically from end to end. The name Branchiostoma refers to the tentacles, thought to be gills, surrounding the stoma (mouth). Amphioxus refers to the fact that the animal is pointed at both ends. The mouth opens into a large pharynx with 50 or more gill slits, leading to an intestine, which opens just in front of the tail fin. All cephalochordates are marine, and they spend most of the time partly buried in sand, with only the head showing.

Figure 3: The lancelet Branchiostoma (=Amphioxus) (about 5 cm), showing internal organs. (From 24).
Amphioxus hatches as a form in which the mouth is turned to the left and there is a single row of gill slits on the right side. It gradually becomes symmetrical as it grows. The left and right sides develop at different rates, but the asymmetrical form was probably not transferred from another taxon and is, therefore, not a true larva in terms of the larval transfer hypothesis [15].
The genome of Amphioxus has been sequenced [25]. It consists of about 520 megabase genes, and it is highly polymorphic.
Cyclostomes: Extant cyclostomes (Cyclostomata), also known as agnathans (Agnatha), are made up of hagfishes (Myxiniformes) and lampreys (Petromyzontiformes) (Figure 4A-C). As the names of the class imply, they have circular mouths without jaws. They are superficially eel-like, but they lack paired fins. They are craniates with short post-cloacal tails. They have notochords rather than vertebrae, but lampreys develop rudimentary vertebral elements dorsally to the notochord. Hagfishes have five to 15 pairs of gills, which share a common opening on each side in Myxinidae but open separately in Eptatretidae. There are seven gill pores on each side in lampreys. Lampreys differ from hagfishes in having disc-shaped or funnelshaped mouths, no barbels, well developed eyes, one or more dorsal fins separate from the caudal fin, and larvae. Adult lampreys live in the sea and in fresh waters, but marine lampreys migrate to fresh waters to spawn. All hagfishes are marine.
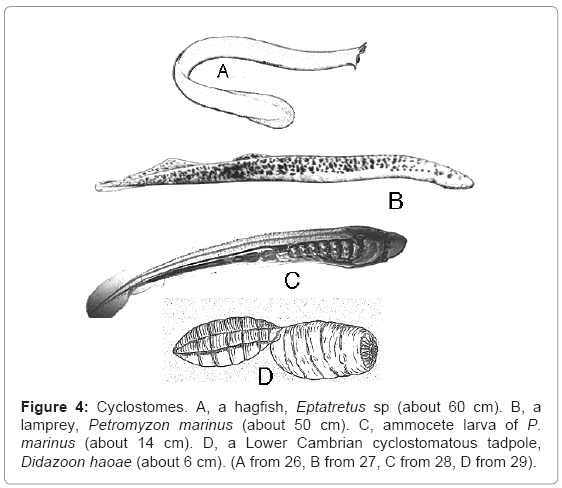
Figure 4: Cyclostomes. A, a hagfish, Eptatretus sp (about 60 cm). B, a lamprey, Petromyzon marinus (about 50 cm). C, ammocete larva of P. marinus (about 14 cm). D, a Lower Cambrian cyclostomatous tadpole, Didazoon haoae (about 6 cm). (A from 26, B from 27, C from 28, D from 29).
Lampreys spend five to seven years as ammocete larvae (Figure 4C), which resemble Amphioxus except that they have only seven gill slits on each side. They are like adult Amphioxus rather than the asymmetrical juvenile, and, like Amphioxus, ammocetes spend most of their time almost submerged in sand. This larva was given the name Ammocoetes before it was realized that it was a phase in the life history of a lamprey.
Didazoon haoae (Figure 4D), from the Lower Cambrian of southern China, has been described as a giant tadpole [30,31]. It was clearly a cyclostome.
Remarks: Cyclostomes have short post-cloacal tails, but, in other respects, they are adult carnivorous craniate agnathan tadpoles. Balfour noted that “the similarity of the mouth and other parts of Petromyzon to those of the Tadpole probably indicates that there existed a common ancestral form for the Cyclostomata and the Amphibia [8].” Didazoon probably resembled this common ancestor.
The ammocete larva of lampreys resembles Amphioxus. This is traditionally interpreted as evidence that lampreys evolved from cephalochordates resembling Amphioxus, but it raises the question, why do hagfishes lack similar larvae? I contend that lampreys and hagfishes had no larvae until an ancestor of modern lampreys acquired larvae by hybridizing with a cephalochordate. The limited fossil evidence is in accord with this view. “Fossil lampreys are admittedly extremely rare, but two of the earliest ones, Mayomyzon and Priscomyzon, are exquisitely preserved as imprints and extremely small, barely the size of an ammocoetes larva. Yet they show no indication of the typical larval anatomy of living lampreys.” [26]. Hagfishes lack larvae because, I claim, no hagfish crossed with a cephalochordate. If this is so, hagfishes will have fewer genes that code for proteins than lampreys, when adjustments have been made for repetition of gene sequences, but comparative figures are not yet available. Studies on cyclostome genomes have revealed unique features of that are not observed in the jawed vertebrates [32]. Comparisons with the known larvacean genome [17] would be welcomed.
Fishes: Fishes are aquatic vertebrates with jaws, scales, usually two sets of paired fins and several unpaired fins. They occur in most seas and fresh waters. The occurrence of larvae is restricted to lungfishes (Dipnoi) and most ray-finned fishes (Actinopterygii), but some ray-finned shore fishes, reef fishes and freshwater fishes lack larvae. No cartilaginous fishes (Chondrichthyes) or coelacanths (Coelacanthimorpha) have a larval phase. Cartilaginous fishes include sharks and skates. Coelacanths are classified with lung fishes as lobefinned fishes (Sarcopterygii).
The swim bladder of a lungfish functions as a lung, so these fishes can breathe air. The Australian lungfish Neoceratodus forsteri (Figure 5) has a single swimbladder, and it occurs in permanent rivers in Queensland. The South American Lepidosiren paradoxa and the four African species of Protopterus are found in swamps and slow moving rivers, and they have paired swimbladders. All extant lungfishes have persistent notochords, but Devonian fossil lungfishes had a full complement of ossified vertebrae [35].Lungfish larvae (Figure 5B) are craniate gnathostomatous tadpoles, each with a cartilaginous skull, a mandible and a notochord. They metamorphose gradually by developing paired fins, scales, lungs, ribs, and neural arches, but the notochord persists in the adult. The tadpole larvae of African and South American lungfishes have external branchiae (thin-walled extensions of the body wall that function as gills).
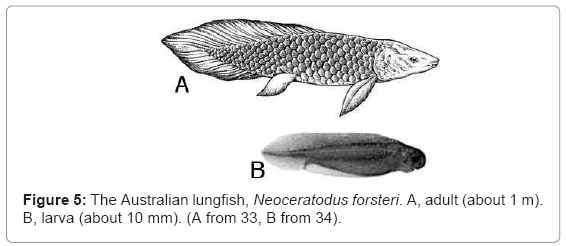
Figure 5: The Australian lungfish, Neoceratodus forsteri. A, adult (about 1 m). B, larva (about 10 mm). (A from 33, B from 34).
All larvae of ray-finned fishes are themselves ray-finned fishes, with cartilaginous vertebrae, skulls and jaws. This is illustrated by eels and their leptocephalus larvae (Figure 6A, B). Freshwater eels migrate to the sea to breed, and North Atlantic species breed in the Sargasso Sea. The larvae of the American eel, Anguilla rostrata, are carried by ocean currents for a year before they reach fresh waters and metamorphose, and it takes three years for the similar larvae of the European Anguilla anguilla to reach land and metamorphose. Leptocephalus larvae are found not only in eels (Anguillidae) but also in bonefish (Albulidae), tarpon (Megalopidae), and ladyfish (Elopidae). The name leptocephalus means small head, but fish larvae in general cover a wide range of head sizes and an assortment of shapes, so there is no ‘typical’ fish larva.

Figure 6: The American eel, Anguilla rostrata. A, adult (about 1 m). B, leptocephalus larva (about 55 mm). (From 36).
Remarks: “Devonian fossil lungfishes had a full complement of ossified vertebrae. These were lost in later lungfishes, and modern ones have a persistent notochord with cartilaginous arcualia and ribs enclosed in perichondral bone.” (Anne Kemp, University of Queensland, pers. comm.). If horizontal gene transfer is ignored, Kemp’s statement may be taken to imply that modern lungfishes, whose Devonian ancestors had evolved vertebrae to replace notochords, flouted Dollo’s law and reinstated notochords. Larval transfer, however, presents a solution to this anomaly that does not involve reverse mutation. I suggest that lungfishes did not have tadpole larvae until a post-Devonian ancestor of extant lungfishes acquired larvae by hybridizing with an adult craniate tadpole. The persistent notochord of modern species is an example of neoteny (retention of larval features by adults). Thomson et al. claimed that the Devonian fossil Palaeospondylus gunni was a larval lungfish [37], but this affiliation was disputed by Joss and Johanson, who listed the many differences between P. gunni and the tadpole larvae of modern lungfishes [38]. It is therefore doubtful whether P. gunni was a larval lungfish, and it was certainly not a tadpole larva. I agree with Joss and Johanson and several other authors that the persistent notochord of extant adult lungfishes is one of a number of neotenic features.
Because the larvae of ray-finned fishes are themselves ray-finned fishes and cover such a variety of shapes, they cannot represent the ancestor of all fishes, as Haeckel would have us believe, nor can they represent the larvae of this ancestor, in accord with Garstang’s theory. I suggest that fish larvae resulted from a series of larval transfers. Female ray-finned fishes lay unfertilized eggs, and I claim that larvae were acquired by many fishes when some of their eggs were fertilized by other species of ray-finned fish. In each case, one species became the larva and the other the adult of the resulting hybrid and its descendants. The occurrence of leptocephalus larvae in eels and in families not closely related to eels fits this explanation. Sharks, skates and coelacanths have no larvae, and I maintain that they are probably not descended from species with larvae. These fishes mate, and fertilization of the egg occurs within the female, so the possibility of hybridization is remote. I predict that fishes with larvae will have more protein-producing genes than those without, but all fishes investigated show much duplication of gene-sequences [39].
Amphibians: Amphibians are tetrapod vertebrates that are not amniotes. Adult amphibians can live on land or in fresh waters; there are no living marine amphibians. The three orders of extant amphibians are (a) Urodela or Caudata (salamanders (Figure 7AC), newts, mudpuppies, sirens (Figure 7D), and amphiumas), (b) Anura or Salientia (frogs and toads) (Figure 7E-G), and (c) Apoda or Gymnophiona (caecilians).

Figure 7: Some adult and larval amphibians (with approximate lengths). A, adult spotted salamander, Ambystoma maculatum (17 cm). B, sexually mature axolotl, A. mexicanum (23 cm). C, tadpole larva of A. mexicanum (9 mm). D, adult lesser siren, Siren intermedia (60 cm). E, adult American wood frog, Rana sylvatica (6 cm). F, larva of R. sylvatica (10 mm), with ventral view of mouth. G, larva of African clawed frog, Xenopus laevis (7 mm). (A from 40, B from 41, C from 42, D from 43, E from 44, F from 45, G from 46).
All urodeles have post-cloacal tails. Sirens (Sirenidae) have no rear legs and very short front legs. All other adult urodeles have two pairs of legs, but both pairs are very short in amphiumas (Amphiumidae). There is no consistent distinction between salamanders and newts.
Mating takes place in water in most urodeles but on land in some lungless salamanders (Plethodontidae). Some female Asiatic giant salamanders (Cryptobranchidae) lay eggs that are then fertilized by the male, who guards them until they hatch [47]. The eggs of other urodeles are fertilized within the female’s body. In most salamanders, including Ambystoma (Figure 7A, B), the male inserts a spermatophore directly into the cloaca of the female. In Lissotriton and some other genera, however, the male deposits a spermatophore in front of the female, who takes up the position previously occupied by the male and presses her cloaca onto the spermatophore.
Some lungless salamanders and the Alpine salamander (Salamandra atra) hatch in a form resembling the adult, without gills [48]. The first larva of most salamanders is a craniate gnathostomatous tadpole (Figure 7C), with a skull, a moveable jaw, external gills, a notochord, but no legs. The gut extends slightly into the posterior half of the body. The notochord and separate nerve cord are shown in photographs by Sobkow et al. [49]. The tadpole metamorphoses into a second larva, which has two pairs of legs and in which the notochord is replaced by a jointed backbone. Ambystoma mexicanum , a species of salamander, can mature in this second larval phase and remain aquatic, when it is known as an axolotl (Figure 7B). On the other hand, if the water is not well oxygenated, the axolotl may metamorphose again to become a terrestrial salamander, with neither gills nor an abdominal dorsal fin. Garstang, with poetic license, implied that axolotls are tadpoles [11], but I regard tadpoles and axolotls as separate developmental phases. Most species of salamander mature only after the second metamorphosis, as terrestrial forms, without gills. Mudpuppies (Proteidae), by contrast, hatch with gills and four legs, and do not metamorphose. They resemble the axolotl phase of A. mexicanum. A siren (Sirenidae) hatches as a tadpole larva, with a notochord, gills, and rudiments of forelegs. It metamorphoses into a form with vertebrae, gills, and small functional forelegs, and it matures in this state.
Adult anurans (Figure 7E) have large hind legs, no tail, and no gills. Fertilization is usually external, and the aquatic eggs hatch as agnathan tadpoles. Fertilization is internal in a small minority of frogs, but most of these also hatch as tadpoles. In Eleutherodactylus and related genera (Eleutherocactylinae, Leptodactylidae), however, fertilization is internal, and the eggs hatch as froglets [50]. In most anuran tadpoles, the anus opens in the anterior third of the body, and the tail occupies the posterior two-thirds (Figure 7F). The gut of some tadpoles, however, extends further back, and the body is less bulbous (Figure 7G). Young anuran tadpoles are without skulls and hinged jaws, as shown in serial transverse sections of Xenopus laevis tadpoles [51]. Later tadpoles have developing skulls and jaw cartilages, but there is usually no articulation between the lower jaw cartilages and the skull until metamorphosis [52]. The mouth of anuran tadpoles covers a wide range of form and is related to diet [53]. It is always much narrower than the head, and opening and closing mechanisms, when present, never involve a hinged lower jaw. When an anuran tadpole metamorphoses, legs, lungs, and a hinged jaw develop, and the small, enclosed mouth changes to a large mouth the same width as the head. The notochord is replaced by a jointed backbone, and the tail is resorbed.
Caecilians (apodans) have no legs and little or no post-cloacal tail. Most hatch in a form resembling the parent, but some, such as Typhlonectes, are born with enormous external gills, which are shed almost immediately.
Remarks: Garstang regarded the axolotl phase of Ambystoma mexicanum as a persistent larva [11], and several authors, including Hanken [54] and Hart and Wray [55], have attributed maturation of this species in the axolotl phase to heterochrony. My counter-view, in terms of larval transfer, proposes that, originally, jawed tadpoles, mudpuppies and salamanders were adult animals without larvae in discrete taxa. The present situation resulted, I suggest, from separate hybridzations between a tadpole and a salamander and between a mudpuppy and a salamander. Ambystoma has a large genome [56], which includes, I claim, a salamander genome, a mudpuppy (axolotl) genome, and a tadpole genome, and the animal can become sexually mature either as an axolotl or as a salamander. The fact that the tadpole phase precedes the axolotl phase in the ontogeny of salamanders is probably unrelated to the sequence in which the two types of larvae were transferred.
All tadpoles swim by undulations of a muscular tail enclosing a notochord and separate nerve cord, but urodeles and anurans have different types of tadpole larvae. Salamanders and sirens have gnathostomatous tadpoles, each with a moveable lower jaw hinged to the skull, while the frogs and toads have agnathan tadpoles, which lack hinged lower jaws. Adult urodeles have evolved less than adult anurans from their common labyrinthodont ancestors, but urodele tadpoles are more evolved than anuran tadpoles. Balfour pointed out the very striking resemblance between anuran tadpole larvae and adult lampreys, not only in the form of the mouth but also in the skull and gill arches [8]. I suggest that anurans had no larvae until some of them hybridized with adult agnathan tadpoles. Eleutherodactylus (discussed below) is a genus of frogs that probably never acquired larvae. Many urodeles also acquired tadpole larvae by hybridization, but in this case with adult gnathostomatous tadpoles. This explains why adult anurans, which are more evolved than adult urodeles, have less evolved tadpole larvae. Several authors (reviewed in [53]) have tried to derive anuran tadpoles from urodele tadpoles, but I suggest that the two types of tadpole larvae were acquired from dissimilar adult tadpoles.
If larvae and corresponding adults evolved from common ancestors, or if larvae were transferred to one ancestral adult species, the classification of larvae should broadly agree with the classification of adults, with small anomalies attributable to different rates of evolution in larvae and adults. Pugener et al. analyzed 43 larval characters and 73 adult characters of a selection of anurans, and they found general agreement between the larval and adult classifications of 20 of the 21 species investigated [57]. The 20 species formed two major clades: (a) the pipoids and (b) all other anurans. The anomalous species was Ascaphus truei, in which the male has a tail-like penis and the species practices internal fertilization. A. truei larvae showed clear affinities with the pipoids, while its adults showed equally clear affinities with the ‘other anurans’. This situation poses problems for the hypothesis that claims that larvae and adults evolved from common ancestors, but it is explicable if anurans acquired tadpole larvae by hybrid transfer from more than one species of adult tadpole. I postulate that early anurans, with no larvae, diverged into ancestral pipoids and ancestral ‘other anomurans’. The ancestral ‘other anomurans’ included ancestral ascaphids and ancestral elutherodactylins, but early elutherodactylins moved to drier habitats. Early pipoids and early ascaphids then acquired larvae by hybrid transfer from one species of adult agnathan tadpole. Early male ascaphids subsequently evolved penes, which led to internal fertilization and migration to turbulent streams. The remaining ‘other anomurans’ (without ascaphids and eleutherodactylins) acquired larvae by hybrid transfer from a different species of adult agnathan tadpole.
The seminal investigations by Pugener et al. covered 21 species of anurans [57]. Comparable investigations of other frogs and toads could reveal other species with incongruous larvae.
Many frogs have become independent of standing water for egglaying, but most of them have a brief tadpole stage in their development. On the other hand, Eleutherodactylus and related genera have no tadpole stage, and they hatch as small frogs. The embryonic development of the skull in E. coqui was studied by Hanken et al., who concluded that “many regions of the skull assume an adult, or postmetamorphic, morphology from the inception of their development”, and “precocious ossification of ……… jaw elements is an evolutionarily derived feature not found in metamorphosing anurans.” [58]. I hold that ancestral frogs had no larvae, and I suggest that an ancestor of Eleutherodactylus retained direct development as it became independent of standing water for reproduction. Early ossification of the jaw in Eleutherodactylus is, I suggest, not ‘evolutionarily derived’ but ancestral. Most other frogs are descended from ancestors whose aquatic eggs were fertilized by freshwater agnathan adult tadpoles.
According to Haeckel’s biogenetic law [7], tadpoles should represent adult forms in the evolution of amphibians, while according to Garstang [9] they should represent ancestral larvae. Modern amphibians, including salamanders and frogs, evolved from labyrinthodont amphibian ancestors, which, in turn, evolved from lobe-finned ichthyostegan fishes [59]. Both Haeckel’s and Garstang’s theories fail to explain why frogs have different tadpole larvae from salamanders, and why both types of tadpole larvae are invertebrate chordates, and therefore less evolved than fishes. Haeckel’s theory implies that, while a branch of lobe-finned fishes was evolving into amphibians, their larvae were evolving backwards, swapping backbones for notochords and, in the case of frogs and toads, losing their hinged jaws. Under Garstang’s theory, salamanders and frogs would have inherited their tadpole larvae from their labyrinthodont ancestors, but there is no explanation of why there are two types of tadpoles or why mudpuppies lack a tadpole phase. My larval transfer hypothesis claims that the two types of tadpoles, like all larvae, were transferred from animals in other taxa by hybridogenesis. Salamanders and frogs acquired their different tadpole larvae by hybridizing with different adult tadpoles, but no mudpuppy hybridized with an adult tadpole, nor did frogs of the subfamily Eleutherodactylinae (family Leptodactylidae).
The short-lived gills of some caecilians may be evidence of a suppressed tadpole larval stage, but this would have been a salamandertype tadpole, with skull and hinged mandible, whose cranial skeleton develops into the adult state with little modification.
I claim that the larval transfer hypothesis offers the most credible explanation, and in some cases the only explanation, of many anomalies in the development of chordates.
Urochordate larvaceans (e.g. Figure 1E) are extant marine adult tadpoles, and fossil tadpoles, including Didazoon (Figure 4D) and Vetulicola, have been described from Cambrian deposits in southern China [30,31]. These fossil tadpoles were about 6 cm long, and they were almost certainly adults. They could not have been larvae of fishes or amphibians because fishes and amphibians had not yet evolved. The larval transfer hypothesis asserts that every type of larva originated as an adult, and I suggest that descendants of these Cambrian tadpoles provided the adult sources of tadpole larvae. Larvaceans probably evolved from one branch of these tadpoles, while another branch evolved skulls and gave rise to cyclostomes. Some adult agnathan tadpoles moved into fresh waters and hybridized with early anurans to give frogs with tadpole larvae. Other craniate adult tadpoles evolved mandibles, and some of these moved into freshwaters and hybridized with lungfishes and salamanders to give jawed tadpole larvae to both groups.
Figure 8 is a simplified summary of the main evolutionary events in cyclostomes, fishes and amphibians, showing the proposed sequence of transfers of larvae by hybridogenesis. The dearth of larval fossils makes it impossible to give a precise geological chronology for larval transfers, but the suggested sequence of transfers fits established adult phylogeny. Figure 8 omits those ray-finned fishes that lack larva, and it implies that anurans acquired larvae in two acts of hybridization, but there may well have been more. It includes the suggestion that an ancestor of living salamanders acquired tadpole larvae by hybridizing with a jawed tadpole. A second possibility is that this ancestral salamander acquired tadpole larvae by hybridizing with a lungfish, one of whose ancestors had acquired a tadpole phase by hybridizing with a jawed tadpole. A third possibility is that African and South American lungfishes acquired tadpole larvae with external gills from salamanders.
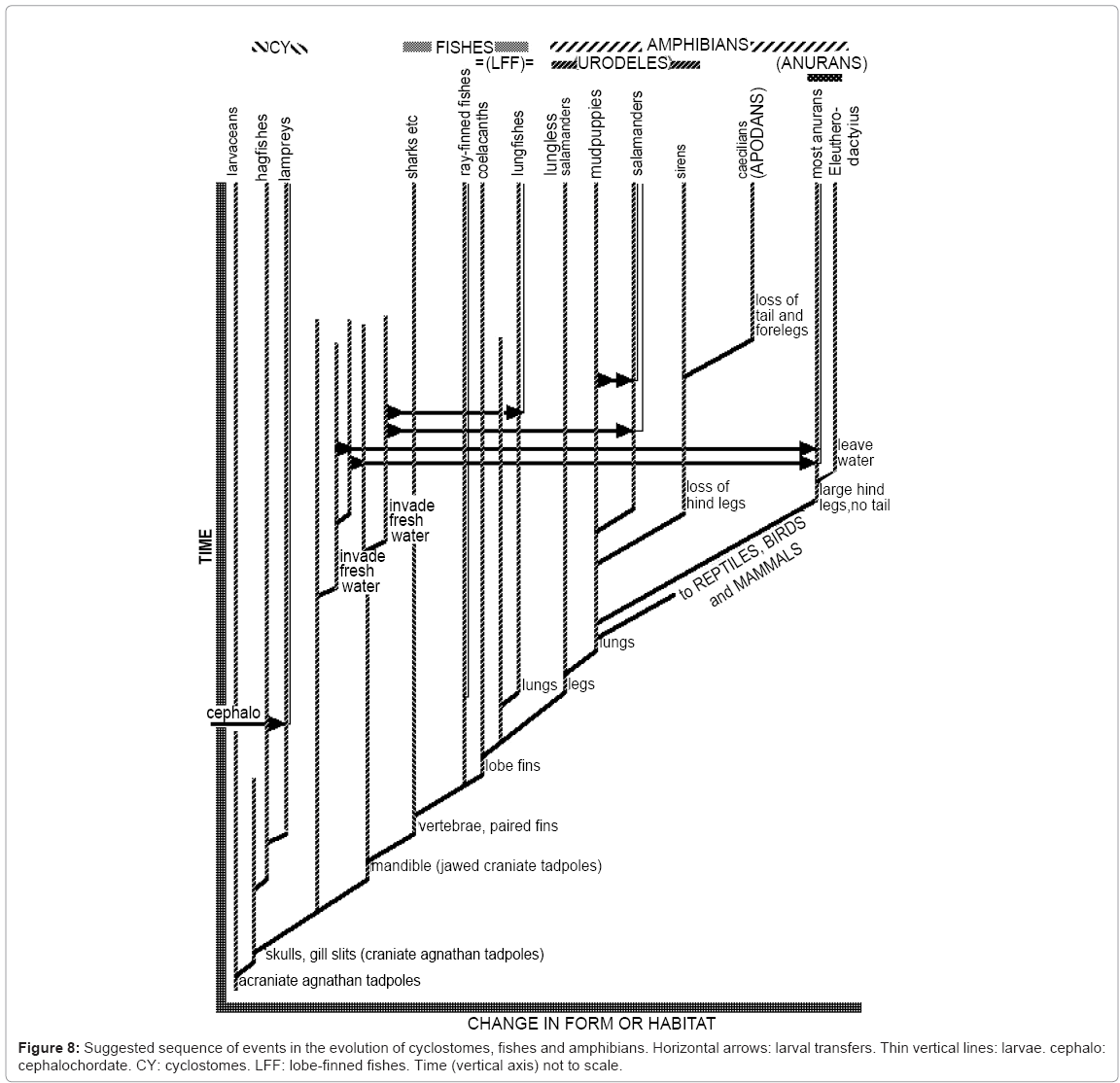
Figure 8: Suggested sequence of events in the evolution of cyclostomes, fishes and amphibians. Horizontal arrows: larval transfers. Thin vertical lines: larvae. cephalo: cephalochordate. CY: cyclostomes. LFF: lobe-finned fishes. Time (vertical axis) not to scale.
The title of the New Scientist article cited above on Cambrian tadpoles is, “The giant tadpole that spawned us all” [31]. This could have come about if some of these Cambrian tadpoles evolved into cyclostomes, which also have notochords and jawless mouths. Subsequent evolution then produced fishes from cyclostomes, amphibians from fishes, reptiles from amphibians, and mammals and birds from reptiles. This implies that tadpoles have a rather better claim than cephalochordates (like Amphioxus) to be ancestors of all vertebrates. My claim that descendants of these Cambrian tadpoles hybridized with some amphibians to produce amphibians with tadpole larvae have no direct bearing on our ancestry. We are probably descended from amphibians with no larvae, as indicated in Figure 8.
The larval transfer hypothesis challenges not only Darwin’s assumption that larvae and adults had a common ancestor, but also Haeckel’s biogenetic law that an animal recapitulates its phylogeny during its ontogeny, and Garstang’s claim that ancestry should be sought in larvae rather than in adults. Under larval transfer, larvae do not represent either adult or larval ancestors of modern species, nor have adults and larvae of the same animal gradually diverged from a common ancestor. Larvae, therefore, tell us nothing about how corresponding adults evolved. The fact that many tunicates have tadpole larvae does not imply that tunicates evolved from tadpoles or that tadpoles and tunicates evolved from a common ancestor. Tadpolelike larvaceans and tunicates were distantly related taxa without larvae, but hybrids between animals in these taxa were tunicates with tadpole larvae, as were descendants of these hybrids. Amphibians evolved from tadpoles (via cyclostomes and fishes), but the fact that many extant amphibians have tadpole larvae is not relevant to the evolutionary ancestry of adult amphibians. Early amphibians had no larvae, but some that had retained external fertilization acquired larvae by hybridizing with adult tadpoles.
The larval transfer hypothesis opposes Garstang’s concept of ‘persistent larvae’: adults that mature in a state that was originally larval. Under larval transfer, marine tadpoles did not originate as tunicate larvae, and freshwater tadpoles did not originate as amphibian larvae. The two sorts of tadpoles were animals in their own right before they came into contact with tunicates or amphibians, and there were tunicates and amphibians without larvae before some of them acquired larvae by hybridizing with adult tadpoles. Tadpole larvae, like larvae throughout the animal kingdom, were later additions to life histories [1,8]. Many authors follow Jägersten in claiming that most animals without larvae, particularly those that live in the sea, are descended from animals with larvae but have ‘lost’ their larvae [60]. Vertebrate examples include coelacanths, sharks and skates; invertebrate examples include cephalopod molluscs and clitellate annelids. I say that such animals never had larvae because they never hybridized with distantly related species, and the fact that they mate and have internal fertilization is fully consistent with this view. I predict that such animals without larvae will have fewer genes that code for proteins than related animals with larvae, when due adjustments for gene replication have been made.
Charles Darwin convinced biologists that plants and animals have evolved, and he described one type of evolution: gradual evolution within separate lineages, which he called ‘descent with modification through natural selection’ [6]. This explains the origins of new species, new genera and new families, but I am convinced that there are two other types of evolution, apart from epigenetics, both of which are rapid, and both of which involve fusion of genomes. They are symbiogenesis and hybridogenesis. Symbiogenesis is the generation of new forms of life by the merger of genomes of organisms in symbiotic association, and it explains the origin of eukaryotic cells (of protists, plants, animals and fungi) from prokaryotic cells (of archaeans and bacteria) [61- 63]. This theory is summed up in the aphorism ‘several prokaryotes make a eukaryote’. The genomes of the different bacteria merged to produce one genome of a totally new organism. Competition between organisms is an essential factor in Darwinian natural selection, but cooperation between organisms is an essential factor in the generation of nucleated cells by symbiogenesis. Hybridogenesis is the generation of new life forms by hybridization: sexual crossing of distinct species at all levels of relationship. The present paper describes how, in my opinion, some chordates acquired larvae by this process while others did not. The same principle explains the occurrence of larvae throughout the animal kingdom [1]. The genomes of different organisms have merged to produce new organisms, and the genomes of different animals have merged to produce animals with new life histories. The links between symbiogenesis and hybridogenesis are highlighted in [64].
I contend that none of the major animal phyla acquired larvae until the classes, and in some cases the orders, of that phylum were established [15]. If so, there must have been a time when animals had no larvae. Hybridization, however, is as old as sex and much older than larvae. I claim that the hybrid mergers of genomes in early animals were an essential factor in the Cambrian explosion, which was a rapid increase in numbers, size and complexity of animals about 550 million years ago. The first examples of animal phyla and classes were products of this melee [15]. Darwin said that the Cambrian explosion was impossible because it conflicted with his views on gradual evolution [6], but there is now a wealth of fossil evidence to confirm that it did take place. The merger of genomes provides explanations of the creation of eukaryotic cells, of the rapid increase in the size and complexity of animals, and of the acquisition of larvae by many animals. It occurs in conjunction with evolution within separate lineages.
Several people have kindly allowed me to reproduce their original illustrations or to quote from their letters. This should not necessarily be taken to imply that they agree with my views.
I wish to thank Lynn Margulis and Farley Fleming for encouragement and constructive criticism. Dedicated to Lynn Margulis (1938-2011), a champion of the larval transfer hypothesis.