Cell & Developmental Biology
Open Access
ISSN: 2168-9296
ISSN: 2168-9296
Review Article - (2014) Volume 3, Issue 2
Annual fishes are found in both Africa and South America occupying ephemeral ponds that dried seasonally. Neotropical annual fishes are members of the family Rivulidae that consist of both annual and non-annual fishes. Annual species are characterized by a prolonged embryonic development and a relatively short adult life. Males and females show striking sexual dimorphisms, complex courtship, and mating behaviors. The prolonged embryonic stage has several traits including embryos that are resistant to desiccation and undergo up to three reversible developmental arrests until hatching. These unique developmental adaptations are closely related to the annual fish life cycle and are the key to the survival of the species. Most of the available data on Neotropical annual fishes derive from studies on the genus Austrolebias. Herein, we review and summarize the available data on the evolution, reproduction strategy, developmental biology and conservation status of these Neotropical fishes.
<Zebrafish (Danio rerio) and medaka (Oryzias latipes) have become widely used fish models because they are oviparous, exhibit long breeding periods, transparency of eggs and embryos, and they are relatively easy to maintain in the laboratory [1,2]. Strikingly different to those model organisms is a group of teleost for which Myers [3] used the term ‘annual fishes’. Annual fishes are found in ephemeral pools of water in Africa and Neotropical regions and complete their life cycle within a year at the end of which they die leaving behind fertilized eggs that typically survive in the soil and hatch during the next rainy season.
Neotropical annual killifishes share the general features of the widely used teleost models mentioned above. However, they are unique in having a very short lifespan, up to 9 months from hatching to death [4]. Ecologically, they are exposed to an extremely variable environment inhabiting ephemeral ponds; these habitat conditions lead to the annual death of the entire adult population.
Life in seasonal and ephemeral aquatic habitats presents numerous challenges. Among vertebrates, the African (Protopterus) and South American (Lepidosiren) lungfishes, face seasonal habitat dry outs. Protopterus avoids desiccation and aestivates during the dry season by burrowing in the mud and forming a cocoon over its entire body, except the mouth [5,6], whereas Lepidosiren do no form cocoon and aestivates in burrows [7]. Among teleosts, the African and Asian clariids catfishes survive dry season by burying in soft sand or in burrows with water-air interface [8]. Burrowing and relatively impermeable cocoons have also been reported in salamanders and anurans; these cocoons consist of accumulation of multiple layers of shed-keratinized skin detached from, but that remains enveloping, the individual. The North American salamander genus Siren inhabits seasonally drying swamps. They survive dry periods by burying in the mud and forming cocoons [9-12]. Among frogs inhabiting deserts or environments with prolonged dry season, cocoon formation evolved independently and has been reported in distantly related genera of North America (e.g., Scaphiopus), Australia (e.g., Cyclorana, Neobatrachus), Africa (e.g., Pyxicephalus, Leptopelis), and South and Central America (e.g. Ceratophrys, Lepidobatrachus, Smilisca) [13-20].
Neotropical annual killifishes are members of the diverse freshwater family Rivulidae (Cyprinodontiformes, Aplocheiloidei) distributed from Mexico and the Caribbean Islands to Southern Argentina [21] and includes both species with annual and non-annual life-history strategies. Within this family, two major clades have been recognized: Cynolebiatinae and Rivulinae [22-24]. The maintenance over evolutionary time of an annual life cycle is coupled with inherently related adaptations in embryonic development, ecological, and behavioral traits [23]. The most striking difference between non-annual and annual killifishes is that the later exhibit arrest development (a diapausing egg). Annualism evolved independently in the African family Aplocheilidae [24]. However, the single or multiple evolution of annualism in Rivulidae is not fully resolved [25,26]. The most recent analyses provide support for two independent origins of annualism in the Rivulidae [27]. Furthermore, 1) the evolution of annualism seems to be favored by the prolonged development of cyprinodontiform eggs and species occurrence in geographical “harsh” environmental conditions [27], and 2) annual species exhibit a noticeable increase in the molecular rate of base substitutions relative to non-annual rivulids [26].
Most of the information on the evolutionary and developmental biology available for the Neotropical annual killifishes derives from studies on the genus Austrolebias Costa, 1998; a genus consisting of 39 currently recognized species and distributed across South-Eastern South America [25,28].
Herein, we reviewed different aspect of the evolutionary biology of the annual fishes with emphasis in the genus Austrolebias.
Throughout the rainy season, adults of Austrolebias inhabit flooded small and medium size temporal pond found in grasslands habitats. These ponds are shallow (50-100 cm deep) with a surface of about 100-500 m2, have aquatic vegetation, turbid water, and soft muddy substrates. Adults are found in the fall, winter and spring months, when the ponds are flooded. They engage in elaborate courtship behaviors that result in the male and female burying in the substrate. While buried they lay desiccation-resistant eggs. These eggs will survive the disappearance of the ponds during the dry season (Figure 1). In the subsequent rainy season, once the ponds are flooded, most eggs hatch. Consequently, the embryos spend their life buried in the mud. Neotropical annual killifish grow fast after hatching and reach sexual maturity fast in about 8-12 weeks and a new reproductive cycle begins [29-32]. In contrast with Neotropical species, most of the African annual species, Nothobranchius, lay their eggs on the surface of the substrate and reach sexual maturity earlier in about 4 weeks [33].

Figure 1: Habitat (ephemeral pond) and biological cycle of annual fishes. (A) Environment during the wet season with flooded ponds when the adults reproduce. Dimorphic colorations of Austrolebias charrua male and female are shown. (B) Environment during the dry season, ponds have disappeared and only embryos remain in the substrate in a pre-hatching state of diapause III. An embryo in this stage and of the same species is shown.
Courtship, mating, and sexual selection
Annual fish life occurs in geographical isolated, discrete, and relative small populations. Consequently, they provide unique opportunities to study mating behavior and in particular to examine the role of sexual selection and the evolution of male traits, male competition, and female choice. Differences, even small ones, in any of these traits have a significant impact on reproductive success. Reproductive isolation would then reinforce their geographical isolation resulting in rapid speciation in this clade of annual fishes.
Striking sexual dimorphism has been reported for several species of rivulids fish and most Neotropical annual killifishes. In Austrolebias, males are usually larger than females. A recent study demonstrated that intra- and intersexual selection on male body size favors larger size in males of A. charrua [34]. Moreover, while females are overall brownish with sparse darker dots on body and fins, males display bright coloration, particularly on the opercular region, unpaired fins, and body flanks. The coloration ranges from bright dots, sometimes disperse over the body flanks and fins (e.g., A. affinis), dispersed over the body flanks and arranged as horizontal bands on dorsal and ventral fins (e.g., A. nigrippinis), to body flanks with distinct vertical bright bands (e.g., A. charrua). Whereas tail fins in Austrolebias can range from dots scattered over the entire fin to a line of dots close to the edge of the rounded fin and a few other dots scattered over the surfaces of the fins. A recent study suggested that that ancestral coloration for both sexes of killifishes was relatively plain [35].
Male coloration plays a crucial role in male reproductive success and male displays are used both in male-male interactions as well as courtship of females. Early courtship studies described a pattern of male displays in A. bellottii, A. nigripinnis, A. viarius, A. luteoflammulatus, and A. cheradophilus [29,36]. These displays consist of an overall pattern of six behavioral units: 1) males fully extend and vibrate dorsal, ventral, and anal fins in front of the females; 2) male quick bursts of swimming alternating with sudden stops or shift on direction; 3) male placing himself in a 45-90 degree angle with the substrate, sometimes introducing the snout into the substrate; throughout these early steps male coloration intensifies. After a while, 4) the female aligns and contacts with the side of the male; 5) the mating couple then buries themselves in the substrate by means of strong movements of the dorsal, ventral, and anal fins; 6) the pair remains buried for variable periods of time, up to 45 seconds, and the individuals emerge from the substrate at separate times and places. This basic behavioral pattern (with the addition and/or modification of behavioral units/displays) has been reported for other species (e.g., A. cyaneus, A. nigripinnis, and A. charrua) [37]. More elaborate analyses of the sequence of behavioral units, as well as frequency and duration of male and female behavioral units were reported for A. reicherti [38]. Undoubtedly, these elaborate and sequentially structured courtships play a role in species recognition and as barriers to hybridization that are evolutionary critical given that more than one species occur sympatrically in the small and ephemeral pools. Indeed, a recent study [38] suggested that the high conservancy in the sequence of courtship displays among species of Austrolebias is involved in avoiding hybridization. Moreover, a recent study demonstrates that A. reicherti responds to chemical cues of potential mates and suggest that chemical cues are essential in the muddy ponds in which this species reproduces [39].
Little is known, particularly under wild conditions, about the natural history and biology of annual Neotropical killifishes, data such as life span, diet, growth rate, egg production, life cycle, and ecology are scarce. This is particularly the case for the genus Austrolebias where the available data is limited to: A. adloffi [40,41], A. bellottii [42,43], A. viarius [44,45], A. cheradophilus [45], A. luteoflammulatus [45], A. nigrofasciatus [46], A. toba [47], A. reicherti [38], A. vandenbergi [48,49], and A. quirogai [50].
Reproduction and sexual strategy
Different sexual strategies are found among teleost fishes. Diversity ranges from gonochorism (testes and ovaries in separate individuals) to hermaphroditism (functional male and female tissues in the same individual) [51]. The only data referring to sex differentiation pattern were reported for A. charrua. According to Yamamoto’s [52] classification, this species corresponds to ‘differentiated gonochoric’ pattern where the early gonads directly develop into an ovary or a testis without intermediate stages [32]
Austrolebias shows a high reproductive potential. After hatching, most species analyzed reach sexual maturity in 8–12 weeks [29,53]. Under laboratory conditions, females spawn daily for about 32 weeks until senescence. Similar data were reported for Cynopoecilus melanotaenia [54].
Previous studies have shown that in most gonochoric teleosts sex differentiation occurs during post-hatching stages and earlier in females than males [55,56]. The first, and up to now only, histological study about sex differentiation and gametogenesis in annual fishes were reported for A. charrua [32]. This histological study reported germ cells proliferation in presumptive ovaries before hatching, demonstrating that sex differentiation occurs at pre-hatching stages. This is a remarkable difference with most non-annual gonochoric fish species where sex differentiation has been reported at various post-hatching stages [56]. However, sex differentiation before hatching has also been suggested for two closely related non-annual fishes Oryzias latipes (Beloniformes) and Gambusia affinis (Cyprinodontiformes) [57,58].
Gamete morphology and fertilization
Fish oocytes develop within the ovarian follicle (oocyte surrounded by follicular cells) and are released during ovulation [59]. The oocyte is surrounded by the vitelline envelope (eggshell, zona pellucida, zona radiata or chorion) that is found between the oocyte and the follicular cells. This structure is deposited in successive layers concomitant with vitellogenesis [60,61]. Teleost fishes, as well as insects and cephalopods, have a special structure in the form of a narrow channel-like structure, the micropyle, which perforates the chorion at the animal pole [62]. This funnel-shaped structure is the only site for the male gamete entry and it is formed during the deposition of the egg envelope. Furthermore, the micropyle opening has taxonomic and systematic value since it shows morphological differences among species and sperm attraction to its opening is species specific [60,62]. The teleost oocyte envelope has been shown to be a sensitive biomarker to adverse pollutants. Furthermore, changes in the organization of egg envelope and in its proteins synthesis have been reported as a consequence of aquatic contaminants (particularly xenoestrogens) [63,64].
Austrolebias oocytes exhibit the general characteristics of other telolecithal eggs (i.e., large amount of yolk, nucleus in the animal pole and just below the micropyle). However Austrolebias’ egg envelope is thicker, a feature that may contribute to prevent dehydration during dry season. In addition, the envelope surface of annual fishes is ornamented with filaments of different sizes, thickness, and distribution patterns among species [32,65,66] (Figure 2A). In freshwater fishes in general [67], and annual fishes in particular [65,68], the ornamentation of the egg envelope is species-specific and a potential source of taxonomic and phylogenetic information.
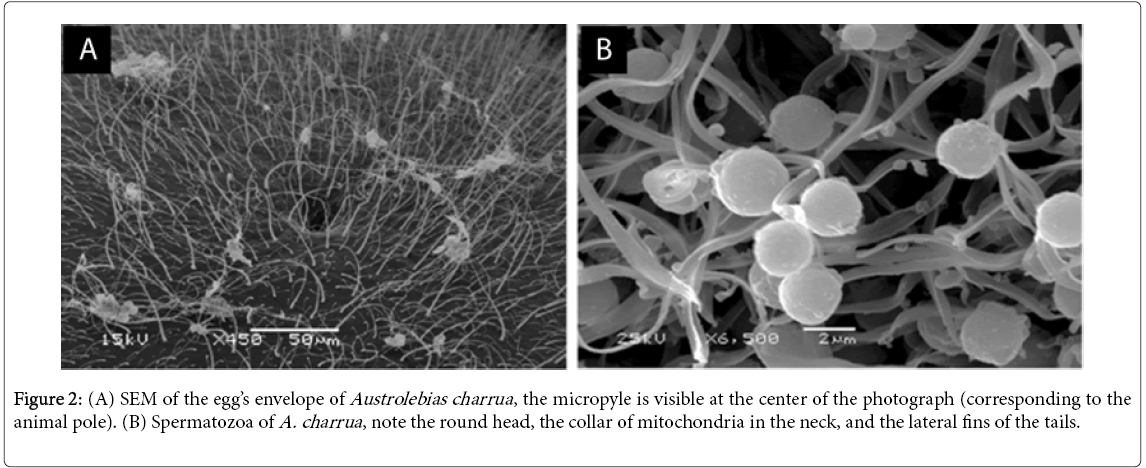
Figure 2: (A) SEM of the egg’s envelope of Austrolebias charrua , the micropyle is visible at the center of the photograph (corresponding to the animal pole). (B) Spermatozoa of A. charrua , note the round head, the collar of mitochondria in the neck, and the lateral fins of the tails.
As other teleosts with external fertilization, annual fishes have a typical uniflagellate anacrosomal aquasperm [69]. At scanning microscopy level, the sperm shows a spherical head, a midpiece containing a single row of round mitochondria, and one flagellum. The flagellar tail could have two, three, or four short lateral fins along almost the entire length (Figure 2B). Teleost fertilization exhibits distinct characteristics from those of other vertebrates and even other “fish” groups. Sperm interaction only occurs at the micropyle of the egg envelope. Since teleost sperm lacks acrosome, there is no acrosome reaction during fertilization. Once near the micropyle, fish spermatozoa show direct movement toward the micropyle opening [70]. Furthermore, this attraction is mediated by a glycoprotein (lectin based) found on the surface of the chorion and its removal result in a reduction of fertilization rate [62]. The sperm fuses to the oocyte cell membrane underlying the micropyle [71]. Therefore, in the cell membrane of both gametes lies the key to species-specificity. Consequently, the ultrastructure of egg envelope, micropyle, and sperm head features are considered in phylogenetic analyses and pre-zygotic isolation among closely related species. Studies of egg envelope ornamentation patterns, sperm morphology, sperm-micropyle attraction in Austrolebias are limited or absent. Available studies on related species, A. reicherti and A. charrua, support the idea that these taxa have emerged from recent cladogenetic events [72,73]; these species could then be a good species- pair model to study these traits.
Analyses of Zona Pellucida (ZP) gene sequences of teleost fishes have demonstrated the presence of two classes of genes that encode ZP proteins and are further distinguished by their expression in the liver, ovary, or both depending on the species. The ancestral condition for vertebrates is to have ovary expression of ZP genes [74,75]. In annual fishes, ZP genes have been identified in A. charrua, (achzpL and achzpH,) both expressed in the liver. The deduced amino acid sequences showed identity values between 65 to 80 % with ZPs from species belonging to a variety of fish groups (Fundulus heteroclitus, Oryzias latipes, Sparus aurata, and Danio rerio) [61].
Embryo development
Annual fishes diverge from other teleosts in two aspects related to their unique developmental pattern [3]. Epiboly is temporally and spatially detached from organogenesis and the embryos undergo one or more reversible arrests (diapauses). These developmental arrests can occur at three different stages: Diapause I during epiboly, Diapause II halfway through somite stage, and Diapause III at pre-hatching stage [66,76-78]. These unique developmental adaptations are closely related to their life cycle and ecological requirements to survive in temporal subtropical and tropical environments.
As described above, egg laying and fertilization in Neotropical annual fishes occurs while the adults are within the substrate. Through meroblastic cleavage, that follow the general teleost pattern, a discoidal blastoderm forms on the animal pole and three other distinct populations of cells can be distinguished: 1) a yolk syncytial layer, where the cleavage nuclei share a common cytoplasm [79,80], 2) a population of deeper cells, and 3) the enveloping multinuclear layer, an outmost layer of big and flat cells that surround and protect the embryo [76].
Teleost gastrulation begins when deep cells move and meet forming the embryo and at the same time epiboly of the enveloping multinuclear layer and yolk syncytial layer occur. Conservation in synchronicity of epiboly and convergence (presence of germ ring) during gastrulation has been shown in comparative studies of early development in several teleost species [81]. The first unique trait of annual fish development occurs during mid-late blastula stage (the dispersed phase) (Figure 3A): deep cells disperse over the entire syncytial layer concomitantly with epiboly of the other two cell populations [66,76,77] The triangular or rhomboidal deep cells progressively migrate toward the vegetal pole. The number of cells at early epiboly, compared with the typical teleost is quite low in all species of annual fishes analyzed [77]. For example, A. viarius has about 100 migrating deep cells [66] whereas in Danio rerio there are about 4000 migrating cells in this early stage [1]. The reaggregation phase occurs after dispersion when deep cells converge over the surface forming an aggregate (Figure 3B). The early reaggregate is one-cell thick but progressively the cells become tightly packed forming a discoidal plate several cells thick [66,76,77,82]. This plate is evocative of the blastodisc of amniotes during gastrulation. Interesting is that, at this stage, the axis of the embryo appears at the middle of the reaggregated plate by mechanisms that remain to be determined. Is the pattern of gastrulation in annual fishes different from the general teleost pattern or is a derived one? A study of early gastrulation in typical teleosts (Salmo, Salvelinus) [83] described a series of changes in the adhesiveness and motility of deep cells. The study reported that, at late blastula stage, these cells separate from one another and randomly migrates. Thus, at the beginning of gastrulation, an incipient dispersed phase has been reported as a general teleost feature and we could consider it a precursor of the dispersed stage of annual fishes. Furthermore, these authors established two overall patterns for teleost development that may contribute to understand the dispersion–reaggregation phases in annual fishes: 1) embryogenesis solely involves the deep cells and 2) the first multicellular structure that forms during embryogenesis (the ‘nubbin’), considered to be equivalent to the prechordal plate, results from the localized ‘accumulation’ (aggregation) of previously ‘disengaged’ (dispersed) deep cells [83]. Similar observations were reported for medaka (Oryzias latipes) [84]. Based on these reports, the departure in the annual fish pattern with distinctly defined dispersed and reaggregation stages could be explained as extensions of the incipient processes previously described in typical teleosts [4].
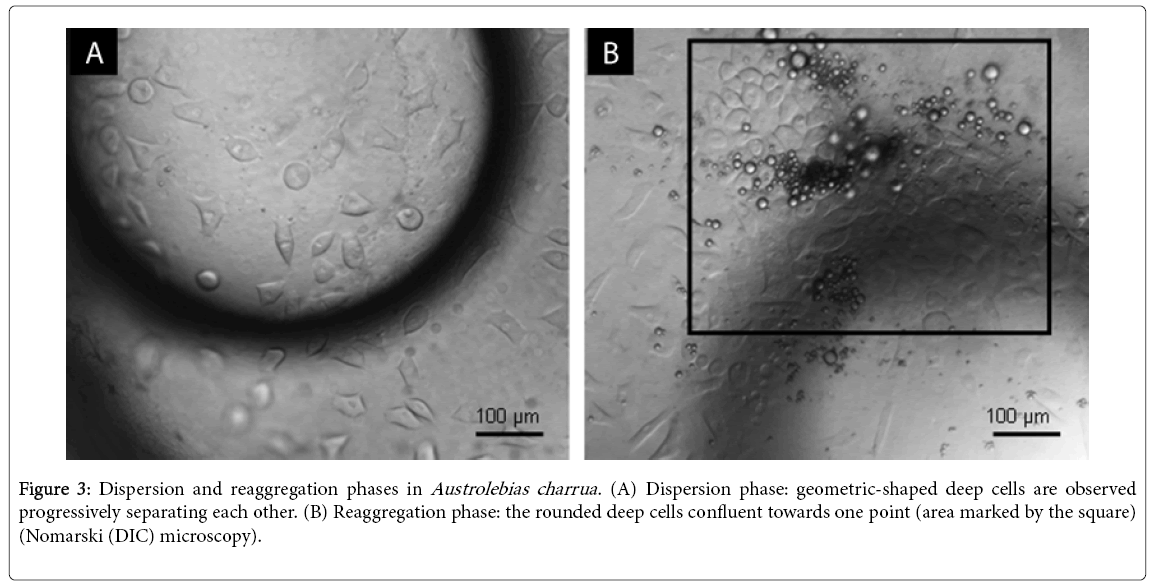
Figure 3: Dispersion and reaggregation phases in Austrolebias charrua . (A) Dispersion phase: geometric-shaped deep cells are observed progressively separating each other. (B) Reaggregation phase: the rounded deep cells confluent towards one point (area marked by the square) (Nomarski (DIC) microscopy).
Subsequent processes of axis formation, somitogenesis, and organogenesis in annual fishes development are similar to those of other teleosts. However, compared with typical teleosts the overall duration of embryo development in annual fishes has been documented to be longer, lasting between 40 to 320 days post-fertilization [78]. Considering that temperature influences the duration of development, temperature variation could explain time differences in developmental rate. In the African Nothobranchius guentheri, an increase of 5 °C resulted in a reduction of 10 days in development [85]. The duration of early embryonic development of two Neotropical annual fishes was compared with three phylogenetically close (Atherinomorpha series) non-annual teleosts: Cyprinodontiformes (Fundulus heteroclitus), Atheriniformes (Odontesthes bonariensis) and Beloniformes (Oryzias latipes) (Table 1) [86].
| viarius (25°C) | myersi (25°C) | F.heteroclitus 20°C | O.latipes (26°C) | O.bonariensis (19°C) | |
|---|---|---|---|---|---|
| Cleavage | 24 h | 7 h | 10 h | 5 h 15 min | 7,5 h |
| Blastula formation | 72 h | 20 h | 24 h | 8 h 15 min | 8,5 h |
| Epiboly | 168 h | 44-48 h | 40 h | 21 h | 38 h |
| Reaggregation | 288 h | 216 h | |||
| Embryonic axis | 312 h | 228 h | 37 h | 21 h | 29 h |
| Somitogenesis (0-10 pairs) | 384 h | 264 h | 56 h | 39 h | 53 h |
Table 1: Comparison of early development duration among annual fishes, Austrolebias viarius [67], Austrofundulus myersi [77] and non-annual fish Fundulus heteroclitus , Oryzias latipes [2], and Odontesthes bonariensis ; culture temperature for each species is noted.
Diapauses
Unique among vertebrates are the developmental arrests or diapauses of variable length that characterized annual fishes development (Figure 4). Furthermore, these could be facultative or obligatory depending on different genera. For example, in Austrolebias, Diapauses I and II are facultative whereas Diapause III is obligatory [78]. This fact has been described in A. nigripinnis [87], A. bellotti [78], A. viarius [66] and A. charrua [66] in embryos cultured under standardized laboratory conditions. In Austrofundulus, Diapause I is facultative but Diapause II and III are obligates [78,88]. Considering the possible combinations among the stages, diapauses can offer eight different developmental pathways to the timing of an embryo trajectory. From an ecological perspective, these possibilities allow the annual fishes to adjust to an unpredictable environment [78].

Figure 4: Diapauses (D) and their placement during the development of an annual fish from fertilization to hatching. The images correspond to embryos of Austrolebias charrua and represent the following events: dispersion (D I), somite embryo (D II), and pre-hatching embryo (D III).
Diapause I, which occurs during the dispersed stage of late blastula, has been interpreted as an early control mechanism imposed by the environment over the developmental pathway [77]. A study in the African Nothobranchius, reported that this stage was induced by chemical signals produced by adult fishes [89]. However, the polar hydrophilic substance responsible for this developmental arrest remains unknown [90].
Diapause II (that occurs at 35-40 somites stage) occurs in advanced embryos undergoing organogenesis [76,78]. Most of the available data about this stage are on the northern South American genus Austrofundulus. Previous studies [88,91,92] have shown that A. limnaeus shows: 1) depressed metabolism during diapause II, 2) most metabolic heat dissipation is due to anaerobic processes, 3) great tolerance to anoxia and the presence of a heat shock protein, and 4) cells are in a G0-like stage of the cell cycle. A study working with the same species explored the role of steroid hormones during Diapause II and showed that treatment of embryos with exogenous E2 induced the increase of the “escape embryos” (embryos that bypass diapauses II in this case) [93]. Furthermore, this study demonstrated that an age related decrease in maternal E2 is correlated with a decrease in the number of escape embryos. Therefore, steroid hormonal signaling seem to be involved in the regulation of diapauses II and progression of development in A. limnaeus.
Diapause III occurs at the pre-hatching stage when the embryo is almost ready to hatch. This stage precisely coincides with the variable duration of the dry season. The overall metabolism of diapause III embryos is lower when compared with those at the same stage that are induced to hatch [88,94]. The resistance to desiccation seems to be related to the ability of embryos to reduce evaporative water loss in A. limnaeus [95,96].
Some non-annual Cyprinodontiformes are able to delay hatching under particular environmental conditions. The non-annual killifish Fundulus heteroclitus usually has continuous development and it’s spawning and hatching coincides with high tide cycles in the coastal marshes that inhabits [97]. However, developing embryos exposed to air showed an accelerated developmental rate, whereas embryos show delay hatching when the eggs are completely submerged under water [98]. Furthermore, these authors found that levels of an aquaporin protein were reduced in air-exposed embryos and suggest that this class of proteins may play a role in water balance. Recently, a study reported that in embryos of F. heteroclitus exposed to air more than 800 genes were differentially expressed. Furthermore, these genes show transcriptional characteristics of ‘stress’ response proteins [99]. In this context, Diapause III could be considered as the addition of the extended delayed hatching as an attribute in annual fish development [78,97].
Conservation status
Most annual fish species, including Austrolebias, are threatened to extinction mainly due to habitat degradation and climate change. Ponds are being altered by different human activities [100]. As a response to this situation, for example, in Brazil the “National Action Plan for the Conservation of Threatened Extinction Killifishes” was recently created. The main objective of this effort is to establish mechanisms to protect and prevent the extinction of rivulids species and loss of their habitats in Brazil [101]. There is high conservation concern for this group of fishes since they are included under the “Critically Endangered” category of Rio Grande do Sul and Brazil [51,101]. In this context, knowledge about the biology and culture techniques of endangered species is essential to support conservation plans, population management, and restocking. Moreover, these studies also offer biological and methodological data to support the maintenance of brood stocks of this important biological model for laboratory studies [100]. In Uruguay in 2013, thirty-three species of Austrolebias were included in the SNAP (Sistema Nacional de Areas Protegidas) [102]
As summarize above, Austrolebias and other annual Neotropical killifishes have unique developmental adaptations that are closely related to the life cycle that make annual fishes a useful model for comparative, developmental, and evolutionary studies. However, in Austrolebias there are many questions that remain to be explored. It has been shown that this genus has specific courtship patterns and sexual selection is important during mating. Courtship has been described for a handful of species that limits our understanding of the role of specific behavioral units in this group. Furthermore, the role of sexual selection on the evolution of male characteristics and speciation has not been tested. Does sexual selection has a stronger evolutionary role in Austrolebias than in other freshwater fishes? Given their annual cycle and that populations inhabit geographical isolated ephemeral pond, one would anticipate a high role of sexual selection in morphological evolution and speciation. No study is available on the sperm-micropyle attraction as a prezygotic isolating mechanism in Austrolebias.
Field studies are needed to understand the natural history of most species; available ecological and population data as well as diet, growth and other phenomenological data is limited. This information would help to understand the dynamics of sympatric populations living in ephemeral ponds. A few studies have suggested the systematic value of the ornamentation of the chorion; however, the ecological role of this ornamentation has not been addressed.
Undoubtedly, the annual cycle and diapausing egg in this group is one of the most fascinating aspects of their developmental biology as one of the most extreme pathways during early development of a vertebrate. Annual fish diapauses are a kind of dormancy stages and an evolutionary innovation that functions as a survival strategy to inhabit harsh environments. These special stages show reduction or suspension of metabolic activities until environmental conditions become favorable. Considering that facultative and obligatory diapauses are of variable duration, sibling eggs can produce embryos that develop and hatch at different times, which in itself could provide an evolutionary advantage for the survival of the species. In this way asynchrony is increased and with it the ability of the organism to face a very unpredictable environment. The recent studies on non-annual killifish suggested that aquaporin proteins, as well as hundreds of other stress-like proteins, may play a role in water conservation during desiccation periods. No studies of gene expression have been done in annual killifishes to understand the molecular basis of annualism. The amazing biology of annual fish exemplifies one of the most extreme events of alternate developmental pathways during early development of a vertebrate.
Currently, studies on the evolutionary biology, ethology, reproductive strategies, regulation of developmental pathways, and senescence on several species of Austrolebias are in progress by different interdisciplinary and international research groups. These approaches should contribute to understand the complexity of annual fishes at the developmental, functional, evolutionary levels.
The authors acknowledge financial support from Facultad de Ciencias, PEDECIBA (Programa de Desarrollo de Ciencias Básicas), CSIC (Comisión Sectorial de Investigación, Universidad de la República), ANII (Agencia Nacional de Investigación e Innovación), and NSF-DEB 1144692.