Journal of Horticulture
Open Access
ISSN: 2376-0354
ISSN: 2376-0354
Research Article - (2014) Volume 1, Issue 2
Background: There are no data regarding the effects of conventional therapies or of dietary antioxidants in hypercholesterolemic or diabetic patients, on their lipid and oxidative stress-related daily rhythms.
Objective: To investigate the effects of Pomegranate Extract (POMx), simvastatin or metformin in hypercholesterolemic or diabetic patients on daily rhythms of serum lipids and oxidative stress, and their effects on macrophage atherogenicity.
Methods: Three healthy subjects, three hypercholesterolemic or two diabetic patients, participated in the current pilot study. Blood samples were collected around the clock from the same patient (every 4 h during a 24 h cycle), before and after two weeks treatment with POMx (1 pill/day), with simvastatin (20 mg/day), or with metformin (850 mgx2/day).
Results: In healthy subjects, but not in hypercholesterolemic patients, HDL-cholesterol levels showed daily changes with a maximal level in the afternoon. In both groups, serum Paraoxonase 1 (PON1) arylesterase activity increased during the day between 08:00 to noon, followed by a decrement between 16:00 to 04:00. In patients treated with POMx or simvastatin, serum oxidative stress decreased at each time point vs. results obtained before therapy. In contrast to the reduction in PON1 activity from noon to evening that was observed in serum before therapy, after simvastatin therapy, there was no such a reduction. In diabetics, serum LDL-cholesterol levels showed daily changes, which were completely abolished after POMx or after metformin treatment. Diabetic serum samples before therapy increased macrophage cholesterol accumulation and oxidative stress, and this atherogenic pattern was beneficially changed after metformin therapy. Serum cortisol levels showed similar daily patterns in healthy subjects and in diabetic patients (maximum level at 08:00) whereas in hypercholesterolemic patients the highest cortisol levels were noted at 04:00. The various treatments resulted in a daily rhythm similar to that of the healthy subjects.
Conclusion: In the present study we have shown, for the first time, that nutritional antioxidant such as Pomegranate Extract (POMx), as well as simvastatin or metformin therapies in hypercholesterolemic or in diabetic patients, respectively, significantly affect their daily rhythms of serum lipids and of serum oxidative stress, as well as their serum-mediated effects on macrophage atherogenicity. These results could contribute to our understanding of how to use the appropriate therapy along the appropriate time of the day for hypercholesterolemic or diabetic patients' therapy.
Keywords: Daily rhythms; Lipids; Oxidative stress; POMx; Simvastatin; Metformin
VLDL: Very Low Density Lipoprotein; LDL: Low Density Lipoprotein; HDL: High Density Lipoprotein; PON1: Paraoxonase 1; SH: Sulfhydryl Groups; POMx: Pomegranate Extract; ROS: Reactive Oxygen Species; AAPH: Azobis: Amidinopropane Hydrochloride; GSH: Reduced Glutathione; MDA: Malondialdehyde; DCFH-DA: Dichlorofluorescin Diacetate; PBS: Phosphate Buffer Saline; DMEM: Dulbecco's Modified Eagle's Medium; PJ: Pomegranate Juice; DM: Diabetes Mellitus; BMI: Body Mass Index; TBARS: Thiobarbituric Acid Reactive Substances; MFI: Mean Fluorescence Intensity; FACS: Fluorescence Assisted Cell Sorting; HBA1c: Hemoglobin A1c
Human circadian rhythms are molecular, cellular, biochemical, physical, and behavioral changes during a 24 h cycle [1-3]. These changes could be correlated with clock-controlled genes expression [4,5], which affect basic cellular processes [6].
In healthy males, daily rhythms were observed for serum cholesterol, triglycerides and glucose, but not for free fatty acids or phospholipids [7]. Clock genes (CLOCK or Nocturnin) regulate dietary lipid absorption in intestinal enterocytes [8,9], thus affecting serum levels of postprandial lipids and lipoproteins [10-12]. Circadian rhythms were shown to be connected to redox homeostasis and to oxidative stress [13-15]. The secretion rhythms of cortisol and melatonin have been shown to be under direct regulation of the circadian clock [16], and melatonin was shown to affect day and night reduced glutathione (GSH) and aldehydes (MDA) levels in healthy adults [17]. Disruption of daily rhythms may lead to the development of disorders such as cardiovascular diseases, diabetes, and cancer [13,18,19]. Indeed, large-scale genetic studies in rodents and in humans have shown the involvement of disrupted clock gene expression rhythms in the pathogenesis of type 2 diabetes [20]. In diabetic patients, circadian changes in hepatic glucose production were directly correlated with circadian changes in blood free fatty acids and cortisol levels [21]. A common variant in the gene that encodes the melatonin receptor (MTNR1B) was associated with impaired glucose homeostasis, reduced insulin secretion and increased risk for developing type 2 diabetes [20].
However, there are no data regarding the effects of conventional therapies in hypercholesterolemic or in diabetic patients, or on effects of dietary antioxidants on lipids and oxidative stress-related daily rhythms. Thus, the aims of the present pilot study were:
(a) To analyze daily changes in serum lipids and oxidative stress in hypercholesterolemic patients vs. healthy subjects.
(b) To analyze the effects of POMx consumption, or of simvastatin therapy in hypercholesterolemic patients on daily rhythms of serum lipids, cortisol and oxidative stress.
(c) To analyze the effects of POMx consumption or of metformin therapy in type 2 diabetic patients on daily rhythms of serum glucose, cortisol, lipids and oxidative stress.
(d) To analyzed the effect of serum harvested before or after therapy on macrophage atherogenicity.
Understanding these effects may improve patients' treatment.
Materials
Phenyl acetate, and 2',7'-Dichlorofluorescin Diacetate (DCFH-DA) were purchased from Sigma-Aldrich (St. Louis, MO, USA). PBS, DMEM and RPMI 1640 medium, FCS, penicillin, streptomycin, nystatin, L-glutamine, and sodium pyruvate were purchased from Biological Industries (Beth Haemek, Israel). POMx was purchased from Pom Wonderful LLC (Los Angeles, CA 90064, USA). POMx is stored in vegan capsules each containing 1000 mg natural pomegranate polyphenol extract, which constitutes 650 mg of gallic acid equivalents, which is equal in polyphenol content to one glass (8 ounces) Pomegranate Juice (PJ).
Subjects
The study was approved by the local ethics committee of Rambam Health Care Campus, Haifa, Israel (approval number 0196/13).
Eight subjects (non-smokers) were included in the study: three healthy subjects, three patients with familial hypercholesterolemia, and two patients with type 2 Diabetes Mellitus (DM). All subjects stayed in the internal medicine department of the hospital during 24 hours of blood sampling. They received their meals in the department in the same time (as described in Table 1). They went to sleep at 22:00 and they maintained in darkness during their normal sleep time. They were woken up for blood collection. Blood samples were collected around the clock from the same patient at 4:00, 8:00, 12:00, 16:00, 20:00 and at 24:00. Blood sample from the hypercholesterolemic patients were collected during 24 hours: before treatment, two weeks after consumption of POMx (1 g/day), and two weeks after statin (simvastatin, 20 mg/day) therapy (there was one week of "washout period" after POMx consumption and before statin therapy). Two new onset type 2 DM patients were also recruited. Repeated blood tests during 24 hours were performed before treatment, two weeks after consumption of POMx (1 g/day), and two weeks after metformin (850 mgx2/day) therapy (with a one week "washout period" after POMx consumption).
| Healthy subjects / Hypercholesterolemics | Kcal | Carbohydrates (gr) | Fats (gr) | |
|---|---|---|---|---|
| Breakfast | 2 pieces of white bread (30 gr) | 150 | 29.58 | 1.20 |
| 1 medium boiled egg | 90 | 0.65 | 6.11 | |
| 5% white cheese (50 gr) | 50 | 2.15 | 2.50 | |
| 1 cucumber (96 gr) | 14 | 3.48 | 0.10 | |
| 1 tomato (114 gr) | 21 | 4.47 | 0.22 | |
| Jam (20 gr) | 52 | 13.18 | 0.03 | |
| Tea with 1 teaspoon of sugar | 20 | 5.00 | 0.00 | |
| Summary | 397 | 58.51 | 10.16 | |
| Percentage per meal | 58.90% | 23.00% | ||
| Snack meal | Apple | 52 | 11.4 | 0.2 |
| Percentage per meal | 87.60% | 3.46% | ||
| Lunch | Chicken (125 gr) | 206 | 0 | 4.46 |
| 1 potato/1/2 cup of rice or grains | 71 | 16.5 | 0.08 | |
| Salad | 35 | 7.95 | 0.32 | |
| 2 pieces of white bread (30 gr) | 150 | 29.58 | 1.20 | |
| 2 spoons of vegetarian oil for the whole meal | 177 | 0 | 20 | |
| Summary | 639 | 54.03 | 26.06 | |
| Percentage per meal | 33.80% | 36.70% | ||
| Snack meal | Banana | 89 | 20.2 | 0.3 |
| Percentage per meal | 90.00% | 3.03% | ||
| Dinner | Soup with 1/2 cup of noodles | 79 | 14.55 | 0.45 |
| 5% white cheese (50 gr) | 50 | 2.15 | 2.50 | |
| 1 piece of potato quiche | 130 | 6.17 | 9.95 | |
| Summary | 259 | 22.87 | 12.9 | |
| Percentage per meal | 35.31% | 44.80% |
Table 1: Diet protocol for healthy subjects, or for hypercholesterolemic patients, or for diabetic patients.
The healthy group included three disease-free subjects as follows:
1. 28-year-old healthy male, over weight (wt 91 kg, ht 1.76 m, BMI 29.4 kg/m2, total cholesterol 187 mg/dL, LDL cholesterol 112 mg/dL, HDL cholesterol 48 mg/dL, triglycerides 134 mg/dL).
2. 29-year-old healthy male with normal weight (wt 71 kg, ht 1.74 m, BMI 23.5 kg/m2, total cholesterol 160 mg/dL, LDL cholesterol 96 mg/dL, HDL cholesterol 41 mg/dL, triglycerides 114 mg/dL).
3. 40-year-old healthy male with normal weight (wt 77 kg, ht 1.76 m, BMI 24.9 kg/m2, total cholesterol 179 mg/dL, LDL cholesterol 111 mg/dL, HDL cholesterol 36 mg/dL, triglycerides 160 mg/dL).
The hypercholesterolemia group included three patients as follows:
1. 50-year-old male patient with non-treated hypercholesterolemia and overweight (wt 92 kg, ht 1.81 m, BMI 28.1 kg/m2). Baseline blood test: glucose 107 mg/dL, creatinine 0.83 mg/dL, total cholesterol 238 mg/dL, LDL cholesterol 161 mg/dL, HDL cholesterol 38 mg/dL, triglycerides 196 mg/dL.
2. 32-year-old male patient with non-treated hypercholesterolemia and obesity (wt 103 kg, ht 1.75 m, BMI 33.6 kg/m2). Baseline blood test: glucose 102 mg/dL, creatinine 0.82 mg/dL, total cholesterol 206 mg/dL, LDL cholesterol 150 mg/dL, HDL cholesterol 34 mg/dL, triglycerides 109 mg/dL.
3. 51-year-old male patient with non-treated hypercholesterolemia and obesity (wt 89 kg, ht 1.72 m, BMI 30.1 kg/m2). Baseline blood test: glucose 116 mg/dL, creatinine 0.83 mg/dL, total cholesterol 253 mg/dL, LDL cholesterol 186 mg/dL, HDL cholesterol 38 mg/dL, triglycerides 146 mg/dL.
The diabetic patients group included two subjects as follows:
1. 35-year-old male patient with new onset type 2 DM and morbid obesity (wt 120 kg, ht 1.75 m, BMI 39.2 kg/m2). Baseline blood test: glucose 345 mg/dL, HBA1C 10.9%, creatinine 0.83 mg/dL, total cholesterol 194 mg/dL, LDL cholesterol 114 mg/dL, HDL cholesterol 32 mg/dL, triglycerides 240 mg/dL.
2. 36-year-old male patient with new onset type 2 DM and obesity (wt 100 kg, ht 1.80 m, BMI 30.9 kg/m2). Baseline blood test: glucose 432 mg/dL, HBA1C 12.8%, creatinine 0.91 mg/dL, total cholesterol 178 mg/dL, LDL cholesterol 119 mg/dL, HDL cholesterol 39 mg/dL, triglycerides 100 mg/dL.
Serum analyses
Serum lipids
Serum lipids [total cholesterol, HDL cholesterol and triglyceride) concentrations were measured in the Clinical Laboratory of Rambam Health Care Campus using automated enzymatic tests (Tayco Diagnostics- Agis Commercial Agencies, Israel).
Serum oxidative stress
Basal serum oxidation status
The levels of serum aldehydes were measured by the Thiobarbituric Acid Reactive Substances (TBARS) assay [22].
AAPH-induced serum lipid peroxidation
The diluted serum samples (x4 with PBS) were incubated with 100 mM of 2.2'-azobis,2-amidinopropane hydrochloride (AAPH, Wako, Japan) for 2 hours at 37°C [23]). The extent of lipid peroxidation was measured by the TBARS assay [22].
Serum SH groups
Serum protein bound SH groups and reduced glutathione concentrations were determined [24].
Serum PON1 arylesterase activity
Initial rates of hydrolysis were determined spectrophotometrically at 270nm for 3 minutes. The assay mixture included 1mM phenyl acetate in Tris buffer. The extinction coefficient is E=1310M-1 cm-1. One unit of arylesterase activity is equal to 1μmol of phenyl acetate hydrolyzed/min/ml [25].
Analyses in macrophages
J774 A.1 murine macrophage cell line
J774A.1 was purchased from the American Tissue Culture Collection (ATCC, Rockville, MD, USA). The cells were grown in DMEM+ 5% FCS.
Serum- induced macrophage cholesterol accumulation
J774A.1 macrophages were pre-incubated for 20 hours at 37°C in the presence of 20μl/ml of the serum samples (derived at each time point during day or night) from healthy subjects or from diabetic or hypercholesterolemic patients before or after therapy. Cellular lipids were extracted with hexane: isopropanol (3:2, v:v).The lipid phase was evaporated under nitrogen, and the amount of cellular total cholesterol was determined using a kit (CHOL, Roche Diagnostics GMbH, Mannheim, Germany).
After lipid extraction, NaOH 0.1N was added to the cells for protein determination by the Lowry method [26].
Serum-induced macrophage ROS formation
J774A.1 macrophages were pre-incubated for 20 hours at 37°C in the presence of 20 μl/ml of the serum samples (derived at each time point during day or night) from healthy subjects or from diabetic or hypercholesterolemic patients before or after therapy. Then the cells were washed with PBS and incubated with 10 μM DCFH-DA, in medium, for 30 min at 37°C [27]. Adherent cells were detached by gentle scraping, and washed (×2) with PBS. Measurement of cellular fluorescence by FACS was performed at 510–540 nm after excitation of the cells at 488 nm with an argon ion laser. Cellular fluorescence was quantitated as mean fluorescence intensity (MFI).
Statistics
Results are expressed as mean ± SEM, n=3 for the healthy subjects and hypercholesterolemic patients, n=2 for the diabetic patients. Each assay was performed in duplicate.
Hypercholesterolemic patients vs. healthy subjects
Daily changes in serum lipid levels
Serum total cholesterol and LDL-cholesterol concentrations (Figure 1A and 1B), were higher in the hypercholesterolemic patients vs. healthy subjects, whereas HDL- cholesterol levels (Figure 1C) were much lower at all time points throughout day and night. Serum triglyceride concentrations were higher in the patients vs. healthy subjects at all time points except for midnight (Figure 1D). From 04:00 to 16:00 there were no changes in the LDL-cholesterol levels in both groups. However, between 16:00 to midnight, LDL-cholesterol levels increased in the patients, but decreased in the healthy subjects (Figure 1B). HDL- cholesterol levels showed daily changes in healthy subjects, with a maximal increase at 16:00 followed by a decrement during the night (Figure 1C).
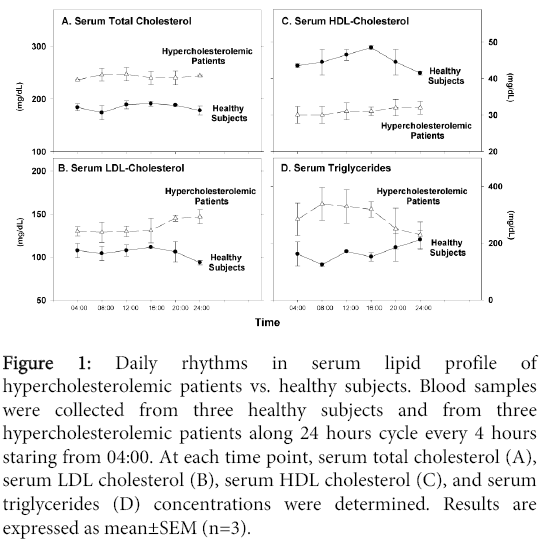
Figure 1: Daily rhythms in serum lipid profile of hypercholesterolemic patients vs. healthy subjects. Blood samples were collected from three healthy subjects and from three hypercholesterolemic patients along 24 hours cycle every 4 hours staring from 04:00. At each time point, serum total cholesterol (A), serum LDL cholesterol (B), serum HDL cholesterol (C), and serum triglycerides (D) concentrations were determined. Results are expressed as mean±SEM (n=3).
In contrast, in the hypercholesterolemic patients no such daily changes were noted (Figure 1C). In healthy subjects, the triglyceride levels increased in the afternoon, with maximal levels at midnight (Figure 1D). In contrast, in the patients there was an opposite trend with an increase in the morning remaining so till noon, followed by a decrease in the afternoon, and reaching lowest levels at midnight (Figure 1D).
Daily changes in serum oxidative stress
We next measured basal serum oxidative status, serum susceptibility to oxidation by the free radicals generator AAPH, serum SH groups levels and PON1 activity (an HDL-associated enzyme which breaks down specific oxidized lipids). As expected, there was increased oxidative stress in the patients vs. healthy subjects as noted by: increased amount of aldehydes in the basal state (Figure 2A), lower levels of SH groups (Figure 2C) and decreased PON1 arylesterase activity (Figure 2D). The extent of AAPH-induced lipid peroxidation however, was similar in both groups (Figure 2B). No daily changes could be noted in both groups for basal oxidative status (Figure 2A), or for AAPH-induced oxidation (Figure 2B). In contrast, daily changes were noted in both groups, in serum SH groups levels and in PON1 activity (Figure 2C and 2D). In healthy subjects, there was a small increase in serum SH groups' concentration at noon time, whereas in the patients, the levels of SH groups, which were low between 04:00 to noon time, increased from 16:00 to midnight (Figure 2C). In both groups, serum PON1 arylesterase activity increased during the day between 08:00 to noon, followed by a decrement between 16:00 to 04:00 (Figure 2D).
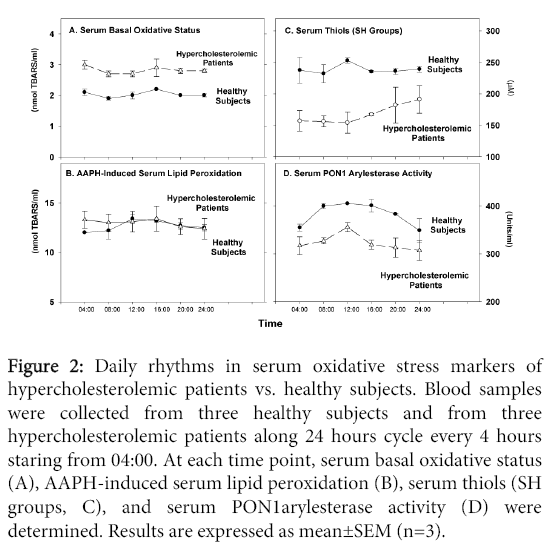
Figure 2: Daily rhythms in serum oxidative stress markers of hypercholesterolemic patients vs. healthy subjects. Blood samples were collected from three healthy subjects and from three hypercholesterolemic patients along 24 hours cycle every 4 hours staring from 04:00. At each time point, serum basal oxidative status (A), AAPH-induced serum lipid peroxidation (B), serum thiols (SH groups, C), and serum PON1arylesterase activity (D) were determined. Results are expressed as mean±SEM (n=3).
POMx consumption by hypercholesterolemic patients
Effects on daily changes of serum lipid levels
POMx consumption by hypercholesterolemic patients did not affect serum total cholesterol (Figure 3A), LDL-cholesterol (Figure 3B), or HDL-cholesterol levels (Figure 3C), as compared to the levels observed at each time point, in serum samples obtained before treatment (Figure 3A). In contrast, serum triglyceride concentrations were decreased, at each time point, after POMx treatment, as compared to triglycerides level observed in the patients' serum before treatment (Figure 3D). As shown in Figure 3, POMx consumption did not affect the pattern of daily changes in serum lipid levels (Figure 3).
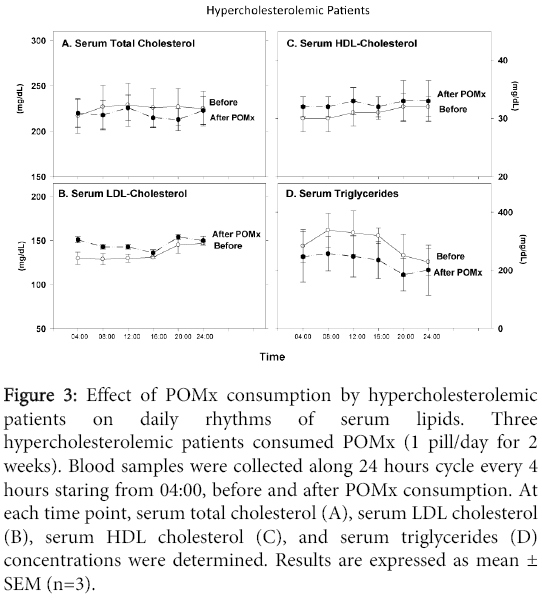
Figure 3: Effect of POMx consumption by hypercholesterolemic patients on daily rhythms of serum lipids. Three hypercholesterolemic patients consumed POMx (1 pill/day for 2 weeks). Blood samples were collected along 24 hours cycle every 4 hours staring from 04:00, before and after POMx consumption. At each time point, serum total cholesterol (A), serum LDL cholesterol (B), serum HDL cholesterol (C), and serum triglycerides (D) concentrations were determined. Results are expressed as mean ± SEM (n=3).
Effect on daily changes of serum oxidative stress
Determination of serum aldehydes levels at the basal state (Figure 4A) or after serum incubation with AAPH (Figure 4B) revealed that POMx consumption decreased serum oxidative stress at each time point, compared to the levels before treatment. Serum SH groups' concentrations (Figure 4C), or PON1 activity (Figure 4D) were not affected by POMx treatment. In contrast, before treatment, serum PON1 activity increased between 04:00 to noon, reaching a maximum at noon time, whereas after POMx treatment, there was a reduction in PON1 activity at these time points. Whereas PON1 activity before treatment decreased from noon time to midnight, after POMx consumption, PON1 activity increased at these time points (Figure 4D).
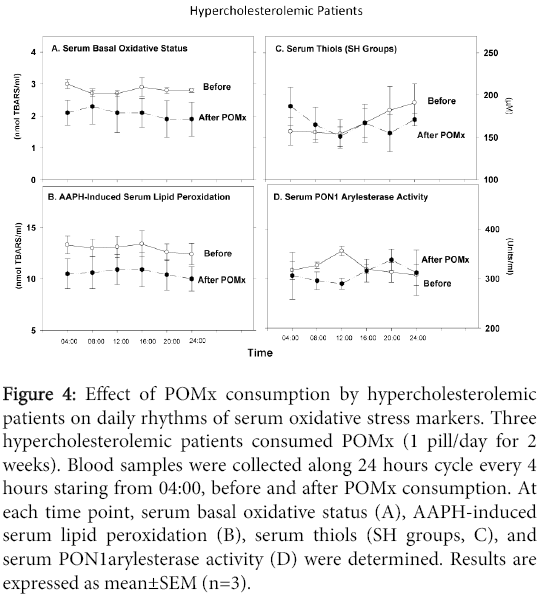
Figure 4: Effect of POMx consumption by hypercholesterolemic patients on daily rhythms of serum oxidative stress markers. Three hypercholesterolemic patients consumed POMx (1 pill/day for 2 weeks). Blood samples were collected along 24 hours cycle every 4 hours staring from 04:00, before and after POMx consumption. At each time point, serum basal oxidative status (A), AAPH-induced serum lipid peroxidation (B), serum thiols (SH groups, C), and serum PON1arylesterase activity (D) were determined. Results are expressed as mean±SEM (n=3).
Effects on serum-induced macrophage cholesterol accumulation and ROS formation
POMx treatment did not affect the patients' serum ability to induce macrophage cholesterol accumulation, vs. serum obtained before therapy. At all time points, POMx treatment decreased the serum ability to induce macrophage ROS formation vs. serum harvested before therapy (Figure 5).
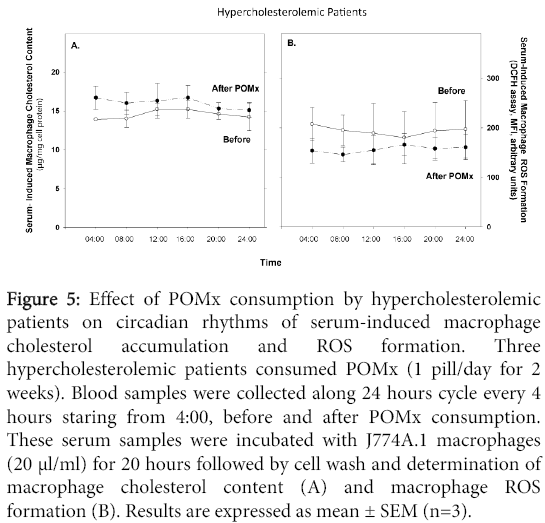
Figure 5: Effect of POMx consumption by hypercholesterolemic patients on circadian rhythms of serum-induced macrophage cholesterol accumulation and ROS formation. Three hypercholesterolemic patients consumed POMx (1 pill/day for 2 weeks). Blood samples were collected along 24 hours cycle every 4 hours staring from 4:00, before and after POMx consumption. These serum samples were incubated with J774A.1 macrophages (20 μl/ml) for 20 hours followed by cell wash and determination of macrophage cholesterol content (A) and macrophage ROS formation (B). Results are expressed as mean ± SEM (n=3).
Simvastatin therapy in hypercholesterolemic patients
Effect on daily changes in serum lipid levels
Simvastatin therapy to hypercholesterolemic patients, decreased their serum total cholesterol (Figure 6A), and LDL-cholesterol (Figure 6B) concentrations, at all time points studied, vs. values observed before therapy. Similar HDL-cholesterol levels were noted before and after therapy (Figure 6C). Serum triglyceride levels were lower after vs. before therapy (Figure 6D). Simvastatin therapy did not affect the daily changes pattern of total cholesterol, LDL-cholesterol or HDL-cholesterol, but it clearly affected the daily changes pattern in serum triglyceride levels. Whereas before therapy maximal triglyceride concentrations were noted at 08:00, followed by continuous reduction towards evening, after therapy an opposite pattern was noted with the lowest concentration at 08.00, followed by continuous increment reaching maximum levels in the afternoon (Figure 6D).
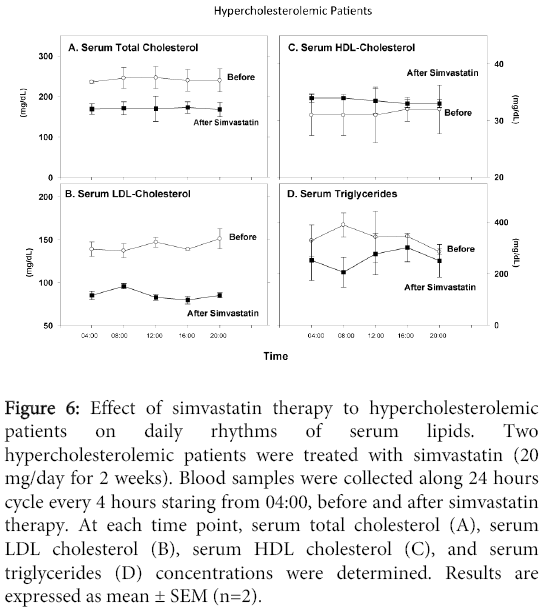
Figure 6: Effect of simvastatin therapy to hypercholesterolemic patients on daily rhythms of serum lipids. Two hypercholesterolemic patients were treated with simvastatin (20 mg/day for 2 weeks). Blood samples were collected along 24 hours cycle every 4 hours staring from 04:00, before and after simvastatin therapy. At each time point, serum total cholesterol (A), serum LDL cholesterol (B), serum HDL cholesterol (C), and serum triglycerides (D) concentrations were determined. Results are expressed as mean ± SEM (n=2).
Effects on daily changes in serum oxidative stress
Simvastatin therapy modestly decreased serum oxidative stress vs. before therapy, as shown by reduction in aldehydes levels in the basal state (Figure 7A), or after AAPH addition (Figure 7B). There was also a significant increase in serum SH groups' concentration after vs. before therapy at all time points (Figure 6C). Simvastatin therapy increased serum PON1 arylesterase activity at all time points except for noon time vs. before therapy (Figure 7D). Simvastatin therapy did not affect the daily changes pattern in basal serum oxidative status, in AAPH-induced oxidation or in SH groups levels (Figure 7A-7D). However, in contrast to the reduction observed in PON1 activity in serum (before therapy) from noon to evening, after simvastatin therapy there was no such reduction at these time points (Figure 7D).
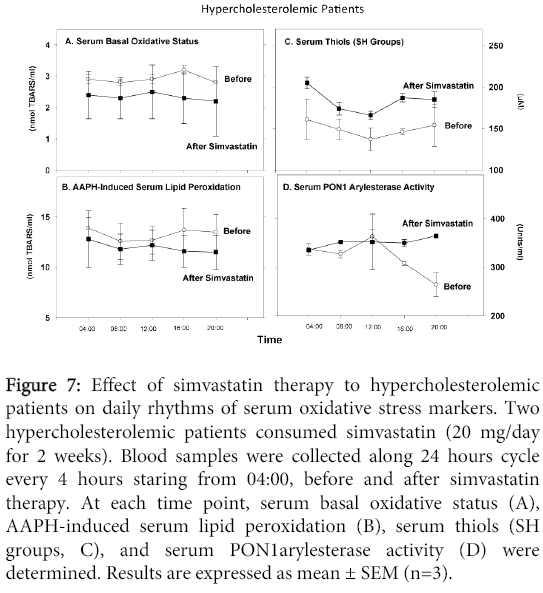
Figure 7: Effect of simvastatin therapy to hypercholesterolemic patients on daily rhythms of serum oxidative stress markers. Two hypercholesterolemic patients consumed simvastatin (20 mg/day for 2 weeks). Blood samples were collected along 24 hours cycle every 4 hours staring from 04:00, before and after simvastatin therapy. At each time point, serum basal oxidative status (A), AAPH-induced serum lipid peroxidation (B), serum thiols (SH groups, C), and serum PON1arylesterase activity (D) were determined. Results are expressed as mean ± SEM (n=3).
Effects of serum on macrophage cholesterol accumulation and Reactive Oxygen Species (ROS) formation
Serum samples obtained from the hypercholesterolemic patients before or after simvastatin therapy were incubated for 20 h with J774A.1 macrophages, followed by determination of cellular cholesterol content and ROS formation. Simvastatin therapy decreased serum-induced macrophage cholesterol content and the ability of the cells to produce ROS, at all time points, as compared to the values observed upon adding serum samples before therapy (Figure 8).
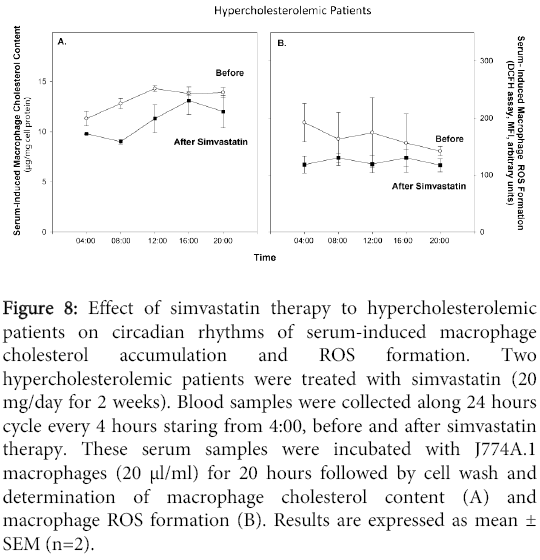
Figure 8: Effect of simvastatin therapy to hypercholesterolemic patients on circadian rhythms of serum-induced macrophage cholesterol accumulation and ROS formation. Two hypercholesterolemic patients were treated with simvastatin (20 mg/day for 2 weeks). Blood samples were collected along 24 hours cycle every 4 hours staring from 4:00, before and after simvastatin therapy. These serum samples were incubated with J774A.1 macrophages (20 μl/ml) for 20 hours followed by cell wash and determination of macrophage cholesterol content (A) and macrophage ROS formation (B). Results are expressed as mean ± SEM (n=2).
POMx or Metformin consumption by diabetic patients
Effect on daily changes in serum glucose and lipid levels
In diabetics, serum total cholesterol (Figure 9A), or LDL-cholesterol levels (Figure 9B) demonstrated similar patterns of daily changes, with an increase in the afternoon (maximum levels at 16:00), followed by a decrease during the evening. After POMx or metformin treatments, these daily patterns were completely modified (Figure 9A and 9B). The patients' HDL-cholesterol levels substantially increased during the morning till noon, reaching a maximum level at 16:00 (Figure 9C). POMx consumption modified this pattern, while metformin therapy had no significant effect (Figure 9C). The patients' serum triglyceride levels were reduced during the morning, followed by a remarkable increment during the evening, reaching a maximum level at midnight (Figure 9D). Both treatments (POMx or metformin) had no effect on this daily pattern (Figure 9D). The patients' serum glucose levels were decreased during the morning, reaching its lowest levels in the afternoon (16:00), followed by an increase, with a maximum levels at midnight (Figure 9E). In contrast, after POMx consumption there was a slightly increase in serum glucose levels in the afternoon, while after metformin therapy serum glucose levels were almost similar during day and night (Figure 9E).
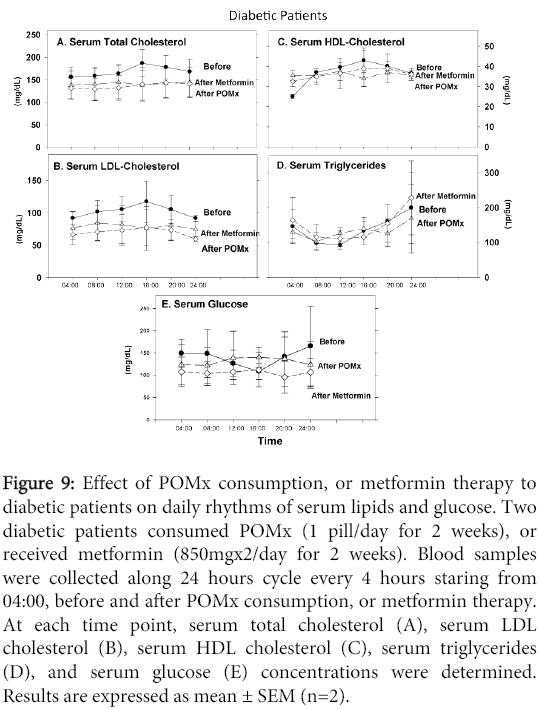
Figure 9: Effect of POMx consumption, or metformin therapy to diabetic patients on daily rhythms of serum lipids and glucose. Two diabetic patients consumed POMx (1 pill/day for 2 weeks), or received metformin (850mgx2/day for 2 weeks). Blood samples were collected along 24 hours cycle every 4 hours staring from 04:00, before and after POMx consumption, or metformin therapy. At each time point, serum total cholesterol (A), serum LDL cholesterol (B), serum HDL cholesterol (C), serum triglycerides (D), and serum glucose (E) concentrations were determined. Results are expressed as mean ± SEM (n=2).
Effect on daily changes in serum oxidative stress
The patients' serum basal level of aldehydes showed remarkable increment during the morning, reaching maximum at 16:00, followed by a reduction in the evening (Figure 10A). POMx consumption or metformin therapy substantially decreased the patients' basal serum oxidative status at all time points (Figure 10A). However, while POMx consumption modified this daily changes pattern, metformin therapy had no such an effect (Figure 10A). In contrast, we did not observe any specific daily changes pattern in AAPH-induced lipid peroxidation before or after POMx or metformin treatments (Figure 10B). Serum SH groups' levels however demonstrated a remarkable increment during the morning and noon, reaching a maximum level at the afternoon (increasing by 19%, Figure 10C), followed by a reduction in the evening and night. After POMx or metformin treatments, the maximum levels were noted at noon (increasing by 14%, Figure 10C). Serum PON1 arylesterase activity substantially increased during the day reaching s maximum in the evening (increasing by 30%, Figure 10D).
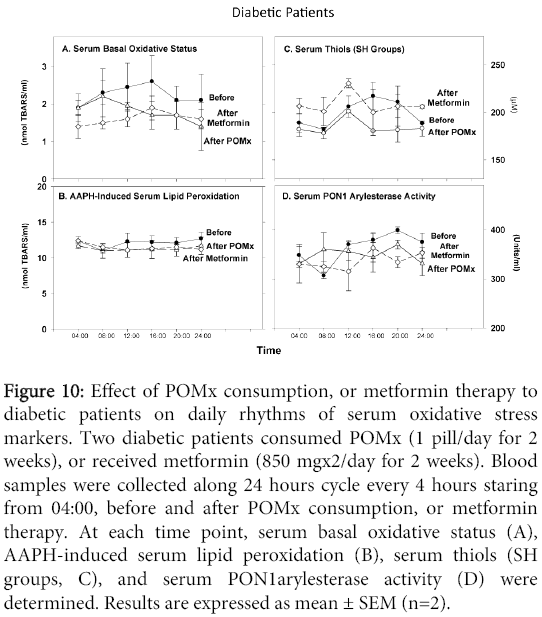
Figure 10: Effect of POMx consumption, or metformin therapy to diabetic patients on daily rhythms of serum oxidative stress markers. Two diabetic patients consumed POMx (1 pill/day for 2 weeks), or received metformin (850 mgx2/day for 2 weeks). Blood samples were collected along 24 hours cycle every 4 hours staring from 04:00, before and after POMx consumption, or metformin therapy. At each time point, serum basal oxidative status (A), AAPH-induced serum lipid peroxidation (B), serum thiols (SH groups, C), and serum PON1arylesterase activity (D) were determined. Results are expressed as mean ± SEM (n=2).
Effects of serum on macrophage cholesterol accumulation and ROS formation
Serum samples obtained from diabetic patients before or after POMx were incubated for 20 h with J774A.1 macrophages, followed by determination of cellular cholesterol content and ROS formation (Figure 11). Serum samples obtained before treatment affected macrophage cholesterol accumulation. There was an increase in cellular cholesterol content during the morning reaching a peak (17% increment) at 12:00, followed by a reduction in the afternoon (Figure 11A). Similar results were observed upon using serum samples after POMx consumption (Figure 11A). Macrophage cholesterol levels at all time points were substantially decreased following incubation with serum samples obtained after metformin therapy (Figure 11A). Baseline serum samples increased macrophage ROS levels by 43% at 8:00 compared to 4:00, followed by a reduction at noon time, and a constant level during the evening and night (Figure 11B). Serum samples after POMx consumption or metformin therapy showed almost similar pattern, but metformin therapy resulted in a remarkable reduction in the macrophage ROS levels (Figure 11B).
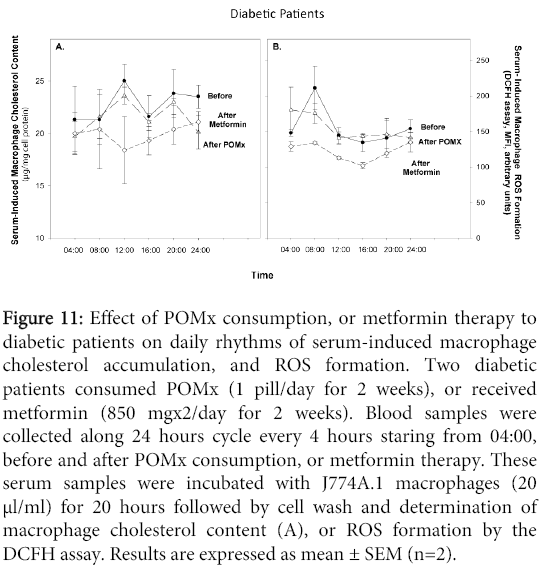
Figure 11: Effect of POMx consumption, or metformin therapy to diabetic patients on daily rhythms of serum-induced macrophage cholesterol accumulation, and ROS formation. Two diabetic patients consumed POMx (1 pill/day for 2 weeks), or received metformin (850 mgx2/day for 2 weeks). Blood samples were collected along 24 hours cycle every 4 hours staring from 04:00, before and after POMx consumption, or metformin therapy. These serum samples were incubated with J774A.1 macrophages (20 μl/ml) for 20 hours followed by cell wash and determination of macrophage cholesterol content (A), or ROS formation by the DCFH assay. Results are expressed as mean ± SEM (n=2).
Daily changes in serum cortisol levels in hypercholesterolemic or diabetic patients vs. healthy subjects: effect of POMx consumption or of simvastatin or metformin therapies
A similar pattern for serum cortisol levels was observed in healthy subjects and in diabetic patients, reaching a maximum level at 08:00, followed by a continuous decrement during the day, reaching the lowest level at midnight (Figure 12A and 12B). In contrast, in hypercholesterolemic patients, the maximal cortisol levels were observed at 04:00 (Figure 12C). POMx consumption or metformin therapy to diabetic patients did not affect the daily cortisol levels (Figure 12B). POMx consumption had also no effect on serum cortisol levels during 24 h in hypercholesterolemic patients (Figure 12C). In contrast, after simvastatin therapy, the maximal cortisol peaks were observed at 08:00 (Figure 12C).
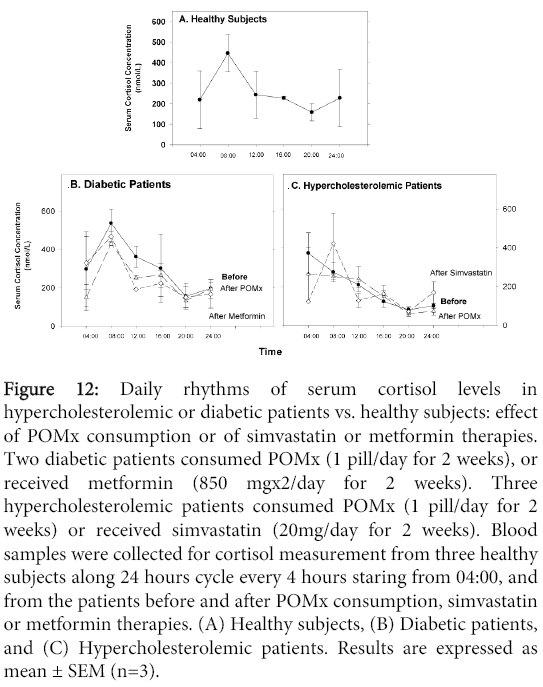
Figure 12: Daily rhythms of serum cortisol levels in hypercholesterolemic or diabetic patients vs. healthy subjects: effect of POMx consumption or of simvastatin or metformin therapies. Two diabetic patients consumed POMx (1 pill/day for 2 weeks), or received metformin (850 mgx2/day for 2 weeks). Three hypercholesterolemic patients consumed POMx (1 pill/day for 2 weeks) or received simvastatin (20mg/day for 2 weeks). Blood samples were collected for cortisol measurement from three healthy subjects along 24 hours cycle every 4 hours staring from 04:00, and from the patients before and after POMx consumption, simvastatin or metformin therapies. (A) Healthy subjects, (B) Diabetic patients, and (C) Hypercholesterolemic patients. Results are expressed as mean ± SEM (n=3).
In the present study we have demonstrated, for the first time, that POMx consumption or simvastatin therapy in hypercholesterolemic patients, and POMx consumption or metformin therapy in diabetic patients, affected their daily rhythms of serum lipids, glucose, cortisol and oxidative stress, as well as the serum-induced macrophage atherogenicity (cholesterol accumulation and ROS formation).
In healthy subjects, we observed daily rhythms patterns for serum HDL-cholesterol, LDL-cholesterol and triglycerides. It was previously shown that the major diurnal changes in normolipidemic males were observed in plasma triglyceride level and in the composition of plasma lipoproteins [28,29]. The daily changes in LDL/HDL/VLDL are related to interactions among these lipoproteins (enzymatic and non-enzymatic) with the loose of surface components from VLDL (to HDL) and with transfer of HDL- LDL associated lipids [29]. The feeding schedule is a powerful synchronizer of the daily lipidic profile [30]. Diurnal triglyceride or LDL profiels may be used to estimate the total daily load of potential atherogenic particles to which individuals are subjected during the day without the need for metabolic studies [31].
In hypercholesterolemic patients vs. healthy subjects, the HDL-cholesterol and triglyceride levels demonstrated an abnormal daily rhythm patterns. This could be related to differences in lipolytic activities in serum, and in dietary lipids absorption. It should be noted that the levels of triglycerides in the patients vs. healthy subjects were higher at all time points throughout the 24h cycle. Simvastatin therapy to hypercholesterolemic patients, which decreased, as expected, serum total cholesterol, and LDL-cholesterol levels, also decreased serum triglyceride levels and changed their daily rhythm profile to patterns similar to those observed in healthy subjects. Animal and human studies have shown that cholesterol biosynthesis exhibits diurnal periodicity with nocturnal increase in the level of cholesterol precursors. Administration of lovastatin in the evening reduced the nocturnal increase in mevalonate levels, and the administration of simvastatin completely abolished the night time rise. Inhibition of cholesterol biosynthesis by statins also modifies the normal diurnal rhythm of cholesterol biosynthesis in female FH patients [32], as observed in the current study.
POMx consumption by the hypercholesterolemic patients also resulted in a decrement in serum triglyceride levels at all time points, and the increment observed before therapy between 08:00 to 16:00 was abolished after POMx. These effects could be attributed, at least in part, to POMx-induced inhibition in triglyceride biosynthesis rate [33].
We observed increased oxidative stress at all time points in serum from hypercholesterolemic patients vs. healthy subjects, as determined by high levels of aldehydes, decreased levels of SH groups and decreased PON1 activity. The increment in the afternoon and evening, in serum SH groups' concentration in the patients vs. healthy subjects, could be a compensatory mechanism to the increased oxidative stress. POMx consumption decreased the patients' serum oxidative stress and beneficially affected the daily rhythms of SH groups and PON1. Similarly, pomegranate juice was shown to reduce oxidative stress in patients with carotid artery stenosis [34], in healthy subjects, or in apoE KO mice [35]. Simvastatin therapy also decreased the patients' serum oxidative stress (basal and AAPH-induced), and substantially increased serum SH groups' concentration, without causing changes in their daily rhythms' pattern. Similarly, simvastatin therapy moderately inhibited LDL oxidation [36]. In contrast to the results obtained before therapy showing a decrement in the patients' PON1 activity in the afternoon and evening, after simvastatin therapy, PON1 activity gradually increased during the day. This may be related to simvastatin-induced upregulation of PON1 [37,38], and/or to the reduction in oxidative stress and triglycerides, since both ROS and triglycerides inactivate PON1 [39,40].
At all time points analyzed, simvastatin therapy also decreased serum-induced macrophage cholesterol accumulation, and ROS formation. This could be the result of simvastatin therapy-induced reduction in serum cholesterol levels and in oxidative stress, and to the direct inhibitory effect of serum simvastatin on macrophage cholesterol biosynthesis rate [41], and ROS formation [42].
Diabetic patients were treated with either POMx or with metformin, a most potent drug against insulin resistance in type 2 diabetes [43]. POMx consumption or metformin therapy changed the daily rhythms pattern of serum glucose, total cholesterol or LDL-cholesterol, but had no effect on HDL-cholesterol or triglyceride levels. These effects could be related to POMx or metformin ability to inhibit cholesterol biosynthesis rate [41,44]. Different circadian variations in oxidative stress markers were also observed in the urine of healthy subjects vs. type 2 diabetic males [45], similar to the current results in serum. POMx consumption or metformin therapy to these patients decreased serum basal oxidative status, and modified the daily rhythms' pattern of serum SH groups and PON1. Similarly, it was shown in diabetic patients that pomegranate juice or pomegranate extract consumption, as well as metformin therapy, significantly decreased their serum oxidative stress [46-48]. Metformin therapy, but not POMx consumption however, decreased serum-induced macrophage cholesterol accumulation and ROS formation.
The limitations of the current pilot study are that it involves small number of subjects, as it is difficult to retrieve new, non-treated hypercholesterolemic patients or non-treated diabetic patients. The experimental procedures are quite complicated as the patients were to be hospitalized and their blood samples were collected (from the same patient) around the clock 6 times during a 24 h cycle, a procedure that was repeated 3 times.
Chronobiological and chronopharmacological studies are important experimental and clinical approaches to get a better insight into the regulation of the cardiovascular system, including their rhythmic organization [49]. Circadian rhythms could affect drug response, as it is possible that a given medication may be therapeutic and safe when administered at some time of the day, but subtherapeutic or poorly tolerated, at another time of the day [50]. We thus recommend that POMx intake for each group of patients should be about one hour before they reach their maximal peak in serum oxidative status.
In the present study we have shown, for the first time, that nutritional antioxidant like pomegranate extract (POMx), or simvastatin or metformin therapies in hypercholesterolemic or in diabetic patients, respectively, affect their daily rhythms of serum lipids and oxidative stress, as well as their serum-mediated effects on macrophage atherogenicity. These results could contribute to our understanding of how to use the appropriate therapy along the appropriate time of the day for hypercholesterolemic or diabetic patients' therapy.
We acknowledge the financial support from the Rappaport Family Research Institute in the Medical Sciences, and from the Clinical Research Institute at Rambam Health Care Campus (CRIR). We thank Professor Raymond Coleman for proof reading and for the editing of our manuscript.