Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Research Article - (2012) Volume 1, Issue 7
Bacteria have the capacity, as all living cells, to escape harm from a noxious agent by extruding the agent before it reaches its target and harms to the cell. This initial response is intrinsic and involves plasma membrane bound efflux pumps which have the capacity to recognise and extrude a large variety of structurally unrelated molecules. When the concentration of the agent is progressively increased, the number of efflux pumps is also progressively increased as a consequence of over-expression of genes that regulate and code for the synthesis of these pumps. Often, when the bacterium is transferred to drug free medium, the number of pump units returns to baseline levels. However, when the concentration of agent is maintained over a prolonged period of time, mutations in genes that code for essential proteins which are usual targets of antibiotics, begin to accumulate and the expression of genes that code for the efflux pumps decreases, oftentimes reaching wild type levels. When the patient is treated with an antibiotic over a prolonged period of time and initial therapy is ineffective, the bacterium most likely escapes via its own intrinsic efflux pump system and with time, it is expected that the number of efflux pump units progressively increase rendering the clinical isolate resistant to the given antibiotic. The amount of energy needed to maintain a high level of efflux pump activity is great and at the expense of other activities that are needed for survival and replication. When this point is reached, and following the second law of thermodynamics, the bacterium goes through changes to survive at low energy costs (for example: switches its mutator gene and a number of predicted mutations of essential proteins take place). These changes render the bacterium permanently resistant to given antibiotics, the need for efflux is no longer and the number of efflux pump units returns to baseline levels. This review will discuss the structure, genetic regulation, physiology of efflux pumps and the means by which a clinical isolate can be characterized for the components that contribute to its resistance during therapy, namely, evaluation of efflux pump activity versus mutations. The ability of a laboratory to perform these evaluations will go a long way toward the selection of effective antibiotic therapy on a real-basis.
<Keywords: Bacteria; Mycobacterium; Efflux pumps; Control; Genetic regulation; Biophysics; Physiology; Inhibitors of efflux pumps; Role of efflux pumps in infection; Role of efflux pumps in quorum sensing
Bacteria have multiple ways to survive the presence of noxious agents in their environment whether that environment is in the wild (toxic agents produced by competing bacteria and other microorganisms [1], toxic agents introduced by human populations [2]) or the human body (antibiotics [3], natural immune defences of the host [4], etc). Concerning the environment of the bacterially infected host, bacteria have the capacity to escape the activity of antibiotics if and when therapy is ineffective due to the presence of mutations that render the antibiotic intended target immune to the antibiotic [5], or to the secretion of enzymes by the given bacteria that destroy the activity of a given antibiotic (examples-lactamases [6]), or to poorly delivered therapy which favours the selection of spontaneous mutations rendering the antibiotic ineffective [7] and induces responses that are transient until the antibiotic is no longer present [8]. Among the transient responses are down-regulation of porins (channels through the bacterial envelope of Gram-negative bacteria which mediate the passage of hydrophilic agents into the cell [9]), induction of two-component regulon systems which render the bacterium relatively immune to many noxious agents [10], and up-regulation of efflux pumps [11]. It is the intent of this mini-review to primarily discuss the structure, function, control and genetic regulation of efflux pumps of pathogenic bacteria and the role that efflux pumps play in rendering the infective bacterium including mycobacteria, intrinsically resistant, transiently resistant and multidrug resistant (MDR) and their interaction with other mechanisms that supplement induced drug resistance such as the two-component regulon systems, quorum sensing and activation of a mutator system that as of this writing remains to be proven. In addition, assays, which can at this time be adopted for the evaluation and quantification of efflux pumps of a clinical isolate, will be presented and discussed.
Efflux pumps are proteins that are present in the cell envelope of all bacteria, are firmly attached to the bacterial plasma membrane and traverse the cell envelope such that a conduit from the plasma membrane to the surface of the cell makes it possible for the extrusion of noxious agents from the cytoplasm and periplasm. Because the efflux pumps of the Gram-negative E. coli have been very much studied and their structure, genetic regulation and control, physiological activity, energy sources for their function, interaction with other drug resistant mechanisms, means by which their function can be reduced or obviated and the means by which they can be evaluated when over-expressed in a given Gram-negative isolate are confirmed, we will restrict our discussion to the acrAB-TolC the main efflux pump of this bacterial pathogen.
E. coli has a wide variety of efflux pumps that may at one time or another extrude a given noxious agent [12]. The main efflux pump of E. coli is the acrAB-TolC pump, which belongs to the genetic family of transporters termed “resistant nodulation division” (RND). This pump has a trimer structure that consists of two fusion proteins coded by the acrA gene that anchor the pump to the plasma membrane of the cell and may mechanically assist the pump [13,14] in the translocation of a recognised noxious agent that is bound to the transporter component of the pump to the external environment. The second component of the efflux pump trimer is the transporter AcrB that is coded by the acrB gene and which has the capacity to recognise and bind noxious agent that are structurally unrelated [15-19]. The means by this recognition takes place is not yet understood. The third component is TolC, which is a barrel like structure that forms a contiguous channel with the AcrB transporter affording the conduit of the noxious agent to the surface of the cell [20-22]. Although the structures of the acrA fusion [17], the AcrB transporter [19] and TolC [23,24] are fully known, the actual position and structure within the cell envelope remains elusive. Nevertheless, a schematic figure of the acrAB-TolC efflux pump of E. coli is presented by Figure 1 modified from that of Pos [17]. Briefly, a noxious agent that has traversed the outer membrane to the periplasm or the cytoplasmic side of the plasma membrane, enters the transporter, is bound and extruded to the outside of the cell via the conduit TolC channel. The process is dependent on energy, which has been commonly accepted as coming from the proton motive force of the cell [8,25]. This concept has been challenged recently and modified within the context of what takes place for the maintenance of the proton motive force (PMF) when the pH of the environment is low or high and the source of protons that supply the energy for the pump [11,25,26].
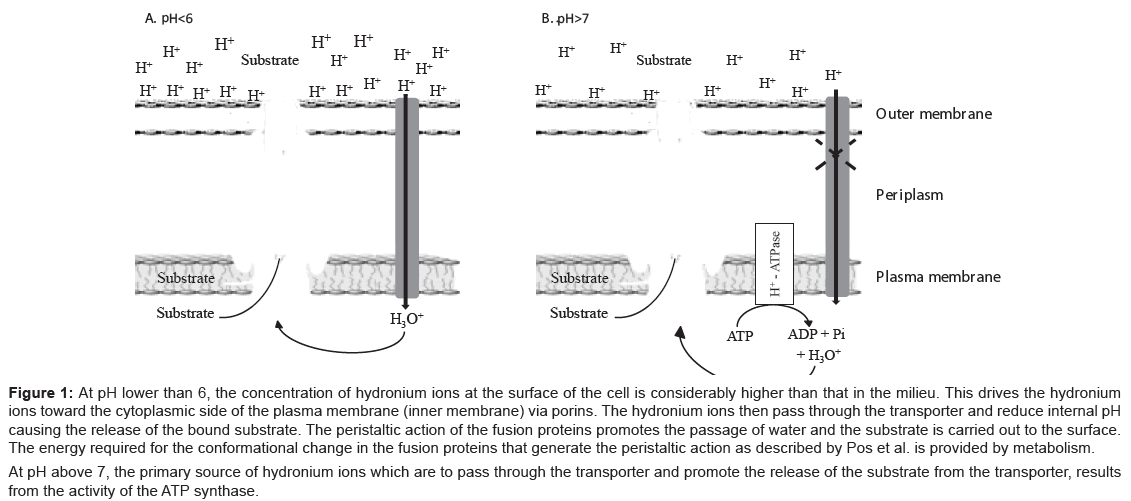
Figure 1: At pH lower than 6, the concentration of hydronium ions at the surface of the cell is considerably higher than that in the milieu. This drives the hydronium ions toward the cytoplasmic side of the plasma membrane (inner membrane) via porins. The hydronium ions then pass through the transporter and reduce internal pH causing the release of the bound substrate. The peristaltic action of the fusion proteins promotes the passage of water and the substrate is carried out to the surface. The energy required for the conformational change in the fusion proteins that generate the peristaltic action as described by Pos et al. is provided by metabolism.
At pH above 7, the primary source of hydronium ions which are to pass through the transporter and promote the release of the substrate from the transporter, results from the activity of the ATP synthase.
The PMF is derived from the differential concentration of protons at opposite sides of a unit membrane as defined by Mitchell’s chemiosmotic hypothesis [27]. The generation of protons results from metabolic activity of the cell and the transfer of these protons to the surface of the plasma membrane cause a relative difference between the concentrations of protons on the surface being much greater than that on the cytoplasmic (medial) side of the plasma membrane. This concentration difference results in an electrochemical gradient termed as the PMF. When certain activities of the cell, such as transport mediated by an RND efflux pump, take place, the movement of these protons to the periplasm makes it possible for the pump to extrude the substrate bound to the AcrB component of the transporter. However, this theory has been modified from results that examined the effect of pH on the source of energy required for the activity of the pump [28-30]. Briefly, whereas at pH 5.5 or below, metabolic energy is not required for the extrusion of an acrAB-TolC substrate such as ethidium bromide, at pH of 6.5 to 7.0, efflux is totally dependent upon a metabolic source of energy (glucose, glycolytic intermediates etc. [31]). Because the main contributor to the PMF is the activity of ATP synthase [32,33], and at low pH, it favours the synthesis of ATP whereas at high pH it favours the hydrolysis of ATP, at low pH protons are not generated by the synthase and the source of protons needed for the function of the pump must lie elsewhere. Mitchell’s theory, when applied to the PMF of bacteria, would result in the movement of protons into the vast ocean milieu that would soon eliminate the differential difference between the two concentrations of protons, and hence, the PMF would collapse [28] The modification of Mitchell’s chemiosmotic theory involves the now known fact that the protons generated by metabolic activity are distributed over the surface of the cell and bound to the lipopolysaccharide layer of the outer cell membrane and to basic amino acids of various proteins adsorbed to the surface of the cell [30]. These bound protons (hydronium ions) produce a pH at the surface of the cell that is two to three pH units lower than that of the bulk milieu [29,30]. Consequently, when the pH of the environment is low and there is a need for protons, the bound hydronium ions are mobilised to the periplasm through porin channels that normally permit the movement of water into the cell [34]. In the absence of metabolic energy, at low pH, the attraction of hydronium ions to the surface of the cell contributes to the PMF. At high pH, the attraction of hydronium ions will be considerably lower, hence the need for metabolic energy as a source of protons that may passively reach the surface of the cell via the efflux pump channel itself and contribute to the PMF [25,26]. Significant to this theory is the recent observation that at high pH the dissociation of a substrate from the AcrB transporter is very slow or not at all whereas at low pH the dissociation constant is very high [35]. Therefore, the movement of hydronium ions through the efflux pump reduces the pH and the AcrB bound substrate is released and carried to the surface by the movement of water [25,26] mechanically assisted by the peristaltic action of fusion proteins acrA [13,14]. The facts presented above also explain why the PMF of a bacterium is maintained in very acidic and very basic media [36-40]. Figure 1 depicts the events related to the anticipated sources of hydronium ions (protons) needed for the activity of the acrAB-TolC efflux pump at low and high pH.
The effect of pH on efflux has been studied and, as an example, Figure 2 demonstrates the effect of low pH (5.5) and the need for metabolic energy when the pH is high (pH 8.0) on the real time efflux of the universal substrate ethidium bromide (EB) by E. coli [41]. Similar studies had also been done for Salmonella enterica serovar Typhimurium [26,42], Enterobacter aerogenes [43], Enterococcus feacalis [44] and Straphylococcus aereus [45]. As noted by this figure, at low pH metabolic energy is not needed whereas at high pH metabolic energy (glucose) is an absolute requirement for efflux of EB. Other sources of metabolic energy can replace the need for glucose at high pH [42]. The method for real time as well as other methods for the evaluation of efflux pump activity will be fully described in later sections of this review.
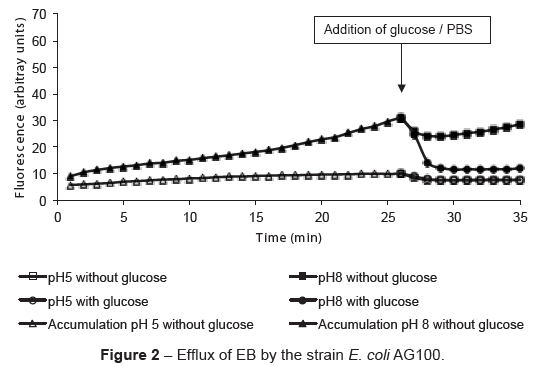
Figure 2: Efflux of EB by the strain E. coli AG100.
The genetic regulation of the acrAB efflux pump involves the activation of stress genes sox and rob sequentially followed by the activation of local regulators mar A and marB, acrA (codes for fusion protein A) and acrB (codes for transporter B). Prolonged exposure to increasing concentration of given antibiotics results in the increased resistance to the antibiotic as well as to other unrelated antibiotics (mdr). Exposure of E. coli to increasing concentrations of tetracycline increases the MIC of the agent [12,46], as well as increases the activity of genes that regulate and code for efflux pump components of the efflux pump acrAB of E. coli (Table 1 [12,46]). The transfer of the bacterium to drug free medium or medium containing an inhibitor of the pump results in the restoration of the MIC as well as the genes that regulate and code for the efflux pump components. However, if the efflux pump is over-expressed and then the organism is placed in medium containing the last concentration of the antibiotic used to induce efflux pump expression, and serially transferred to this constant concentration, with time, the MIC to tetracycline continues to increase whereas the activity of the gene that codes for the AcrB transporter returns to baseline levels (Table 1) [47]. The cause for the progressive increase of resistance to tetracycline as well as to the main antibiotic targets of beta lactams, 30S ribosomes and gyrase [47] is due to the accumulation of mutations with prolonged exposure to a constant concentration of a given antibiotic. This development of an MDR phenotype is predicated on the initial response to the antibiotic, which is the increased activity of genes that regulate and code for the acrAB-TolC efflux pump. In other words, the increased expression of the efflux pump system makes it possible for the generation and accumulation of mutations in the essential proteins of the organism. It also explains why a constant dose of an antibiotic for a prolonged period of time yields a clinical isolate for which the MIC of the given antibiotic may increase to 10, 50 or even 100 fold over that for the wild type strain.
| Genes | AG100TET [12,46] | AG100TET10 [47] |
|---|---|---|
| acrF | ↑↑ | ↑ |
| acrA | ↑↑↑ | n.a. |
| acrB | ↑↑↑ | ↑ |
| acrR | ↑↑ | n.a. |
| marR | ↑↑ | n.a. |
| marA | ↑↑↑ | ↑ |
| marB | ↑↑ | n.a. |
| tolC | ↑↑ | n.a. |
| micF | ↑↑↑ | n.a. |
| ompF | ↑ | n.a. |
| ompX | ↑↑↑ | ↑ |
| soxS | ↑ | ↑ |
| rob | ↑ | ↑ |
Table 1: Changes in the expression of genes that code for or regulate the expression of AcrAB efflux pump. Table represents the relative quantity of gene mRNA compared with the parental strain AG100 not adapted to tetracycline (TET). ↑↑↑-increase over 6 times; ↑↑-increase over 2 times; ↑-slight increase till 2 times; →-same level of expression; n.a.: data not available. AG100TET refers to the strains after increasing concentrations of TET [12,46]. AG100TET 10 refers to the strain AG100TET after serial passages at 10 ug/mL of TET [47].
Inducement of an efflux pump system of Mycobacterium tuberculosis by exposure to increasing concentrations of isoniazid (INH) has also been shown [48,49]. Moreover, as noted for E. coli prolonged exposure to a constant concentration of INH will at first be accompanied with increase in the expression of genes mmpL7, p55, efpA, mmr, Rv1258c and Rv2459 that code for the main efflux pumps of the organism followed by mutations in essential genes [49].
Similar results have been obtained with Gram-positive bacteria [45].
Communication between bacteria of the same strain or species and between species contributes to their survival [50-52]. The secretion of signals that invoke a specific response from the responder is termed Quorum sensing (QS). When it takes place between strains of the same species, communication is directed towards the reduction of population growth and hence, reducing the possibility of exceeding the nutritional support of the environment [53, 54]. Other signals involve the secretion of bioactive molecules that inhibit the replication of a competing population species [55] or even kill (biocidins) [56] or promote a swarming effect that recruits members of the same species to migrate to a specific location [57,58]. Other responses involving “community action” are also invoked and these result in secretion of materials that will protect the bacterium from external danger. These materials, termed biofilm, encase the bacteria at distances from each other and within the matrix of this biofilm are channels used for further communication [59-61]. Biofilms are produced in the wild to maintain the bacterial population in situ (for example: surface of rocks) [31] and are also produced at sites of the human colonised by infecting bacteria [62]. There is a relationship between efflux pumps (EP), QS and biofilm secretion, which has come to the forefront only recently [13]. In this case, the absence of the main efflux pump of a Gram-negative bacterium inhibits the secretion of biofilm suggesting that the efflux pump provides the means by which the synthesised biofilm is extruded much as is the case for the extrusion of toxic products produced by the metabolic processes of the bacterium [63]. These aspects suggest that efflux pumps perform other roles than simply extruding noxious agents that have penetrated the cell envelope. To this extent the question of whether efflux pumps have a role in the secretion of QS signals has been indirectly shown by the demonstration that agents which inhibit the PMF of a Gram-negative bacterium not only inhibit the activity of an efflux pump but also inhibit responses from a bacterium to which the QS signal is directed [52,64]. As shown by Figure 3, agents such as trifluoromethyl ketones (TKFs) that inhibit the PMF of bacteria [52], and their efflux pump sy stem also inhibit the response to a QS signal [52]. Other inhibitors of the PMF such as phenothiazines that also inhibit efflux pumps of bacteria have also been shown to inhibit the responses to QS signals.

Figure 3: The effect of a trifluoromethyl ketone (TF) on the response of CV026 to the QS signal acylated hydroxyl lactone (AHL). A: Control disk with TFK MM4 (20 μg) alone (no colour). B: Control disk impregnated with QS signal pure acylated hydroxyl lactone (AHL) (10 ng) alone (deep purple coloration indicates response by the responding bacterium that is obviated by the TFK MM4 compound) [52].
Other studies will more closely examine the relationship between the efflux pump and secretion of QS signals since the inhibition of the latter is considered to be of great significance for the prevention of biofilm secretion, as well as other secretions that promote tissue damage to the infected host.
The neutrophil does not readily kill phagocytosed Salmonella once it is entrapped by the lysosome [10]. The reason for this is due to the inducement of the pmrA/pmrB two component regulon by the acidic pH of the phagolysosome that results in rendering the organism practically resistant to almost everything [10]. The pmrA/pmrB regulon is activated by pH [10] and functions as follows: Activation of the pmrb gene produces a product pmrB, which undergoes self-phosphorylation. The phophorylated pmrB acts as a kinase and transfers the phosphate to pmrA which activates a series of nine genes that result in the synthesis of Lipid A that is introduced into the nascent polysaccharide (LPS) layer of the outer membrane of the organism [10]. Because this increased LPS renders the organism immune to the action of lysosomal hydrolases and prevents the penetration of antibiotics [10]; a systemic infection that results from the invasion of the organism into the peritoneal cavity is accompanied by high mortality. Clinical isolates of Salmonella from peritoneal infections or extracted from the phagolysosomes of the neutrophil have over-expressed efflux pumps. These overexpressed efflux pumps result from the activation of the gene pmrd by the pmrA protein and the product of pmrd, pmrD, directly activates ramA, the global regulator of the acrAB efflux pump of the organism [10]. Over-expression of the acrAB-TolC efflux pump contributes to the difficulties associated with the therapy of a Salmonella infection of the colon [26]. Although an inhibition of the efflux pump system of the strain whose pmrA/pmrB two-component regulon has been induced will not contribute much to therapy of a Salmonella peritoneal infection. The Salmonella strain that eventually colonises the colon is expected to have its efflux pump over-expressed as it passes through the stomach (pH as low as 3 or less) as a consequence of the activation of its induced pmrD product. Moreover, when the organism reaches the colon, it is already prepared to extrude the toxic bile salts present [23,41]. Consequently, inhibition of the acrAB-TolC efflux pump of colon-colonising Salmonella by non-toxic agents known to inhibit the pump may effectively treat a Salmonella infection of the colon when used in combination with antibiotics.
The response of Salmonella to chlorpromazine (CPZ) and thioridazine (TZ) has resulted in the understanding that the MIC determined 16 h later does not fully characterise the response of the organism. As an example, during the first 8 hours of exposure to various concentrations of CPZ, the organism does not grow, demonstrating susceptibility to the agent. However, following this period, the organism grows at the same rate as the drug free control culture [65]. The same result is obtained with TZ [42]. Obviously, the events after the initial period of 8 hours, an adaptive response must have taken place. As evident from the data presented by Table 2, the genes that regulate and code for efflux pumps of Salmonella are sequentially expressed during the first 8 hours of culture [42]. By the end of 16 hrs, the MIC of TZ is over 200 mg/L and the expression of efflux pumps has returned to almost normal baseline levels [42]. Nevertheless, the activity of the efflux pump after 24 h of exposure is not sufficient to permit the organism to survive a sub-inhibitory concentration. Survival appears to involve huge changes in the appearance of the cell envelope as well as the protein composition extracted from this envelope [65]. These studies show how well equipped Salmonella is for dealing with high concentrations of a noxious agent in its environment suggesting that in all probability, the dangers posed by food-borne infections of Salmonella will be with us for a long time.
| Genes | 0.5h | 1h | 4h | 8h | 16h |
|---|---|---|---|---|---|
| soxS | → | ↑↑ | ↑ | → | → |
| rob | ↑ | ↑ | ↑ | → | ↑ |
| ramA | ↑ | ↑ | ↑↑↑ | ↑↑ | ↑ |
| marA | → | → | ↑↑ | ↑ | → |
| acrB | → | ↑ | ↑ | ↑↑↑ | ↑↑ |
| pmrA | ↑ | ↑ | ↑ | ↑↑ | ↑ |
| pmrB | → | ↑ | ↑↑ | ↑ | ↑ |
Table 2: Activity of genes that regulate and code for the AcrAB efflux pump of Salmonella. During the first 8 h, the organism was not growing, but genes evolved in the expression of the AcrB transporter were sequentially activated: first soxS, whose expression was elevated after 1h of culture; followed by ramA, marA and pmrB (4 h culture). After 8 h of culture, ramA decreased its activity, marA returned to baseline activity, acrB was maximally increased in activity and pmrA was now active. By the end of the 16 h culture, only acrB activity remained elevated. Table represents the relative quantity of gene mRNA compared with the untreated Salmonella strain: ↑↑↑-increase over 15 times; ↑↑-increase from 5 to 15 times; ↑-increase till 5 times; →-same level of expression.
We have developed a number of methods that can easily be adapted to the daily routine of a general bacteriology laboratory. Among the simplest to use is the EB (ethidium bromide) agar method that requires no instrumentation and its preparation evaluation of results is performed with material that is universally present in these laboratories.
The fluorochrome EB is a universal efflux pump substrate [66] which when added to agar in increasing concentrations will identify the highest concentration that a clinical bacterial isolate can readily extrude. This concentration is readily noted by the presence of pink fluorescence associated with the colonies present on the surface of the cell [67-69]. The higher the concentration of EB that produces fluorescence of the colony as compared to that of a wild type reference control of the same species, the more active is the efflux pump system of the clinical isolate. The method is simple, reproducible and can readily be phased into a bacteriological laboratory. No special skills or laboratory equipment other than a short/long wave hand held UV light, is needed. For a more effective evaluation, a UV box will do the trick. The method and its variations have been fully described [67-69]. Figure 4 shows a schematic presentation of a common EB plate, which can be used for evaluation of as many as 8 to 12 strains at the same time. Characterisation of the clinical isolate in a quantitative manner is also possible and the reader is encouraged to adapt this part of the method [69]. For those situations where the use of EB is controlled or prohibited, a replacement method has been developed using acridine orange as the efflux pump substrate [70].
Figure 4: Schematic representation of the EB or AO plates. Cells are swabbed in the plate in lines that start nearly the center (without touching each other) to the outer part of the agar.
Assessment of efflux pump activity conducted by the automated EB method has been completely described [72]. This method affords a real-time estimation of accumulation of EB and its efflux. Briefly, bacterial strains are cultured in a suitable medium until they reach an optical density at 600nm of 0.6 and centrifuged. The pellets are washed twice with saline buffer, the OD at 600 adjusted again to 0.6 in glucosefree saline buffers at pH 7 (or other according to the objective of the experiment). Because metabolic energy is required for efflux of EB, the absence of glucose ensures sufficient accumulation of the agent. The final concentration of EB for all experiments is to be determined empirically for each bacterial species (note concentration of EB varies from 0.5 mg/L for enterococci and may be as high as 3 mg/L for Salmonella). At concentrations of EB that exceed the ability of the cell to extrude the agent, the level of intracellular agent rapidly increases and results in its intercalation between the nucleic bases of DNA. EB when bound to DNA is no longer available for extrusion [71]. The tubes are rapidly transferred to a Corbett 3000 thermo cycler (Qiagen, Doncaster, Australia) programmed with cycles of 1 min at a constant temperature of 37°C. Accumulation of EB in each tube is followed on a real-time basis by assessment of fluorescence emitted. Excitation and emission wavelengths are 535 nm and 585 nm, respectively. Whereas the medium containing as much as 4.0 mg/L of EB does not appreciably fluoresce, as the concentration of EB builds up in the periplasm of the Gram-negative bacterium, the instrument readily detects fluorescence.
For controls that demonstrate the accumulation of EB that results from a concentration of EB that exceeds the cell’s ability to extrude, varying concentrations of carbonyl cyanide m-chlorophenylhydrazone (CCCP) and Phe-Arg-naphthylamide (PAβN) can be used as described [71]. The methods for determination of efflux, physiological parameters that affect efflux and evaluation of compounds for activity against the efflux pump of mycobacteria have been fully described [72]. The data presented by Figure 2 present raw data directly from the Corbette 3000 instrument used.
The response of bacteria to a noxious agent such an antibiotic initially involves intrinsic efflux pumps, which can be over-expressed as the concentration of the agent increases. When the concentration of the agent is then maintained, efflux pumps continue to be over-expressed and mutations in essential genes take place, supposedly by activation of a master mutator gene. Soon thereafter the level of efflux pump activity returns to baseline level. The sequence of these processes has been confirmed for mycobacteria and other bacterial species. Moreover, as is the case for all matter in the universe, conservation of energy goes from a high to a lower level (second law of thermodynamics), and bacteria are no exception. Therefore, because the maintenance of high efflux activity extols a high-energy cost, mutations that render essential proteins immune from antibiotic actions provide a more energy efficient response that can be maintained indefinitely. However, if the environment is purged of the noxious agent, the bacteria that contain mutations in essential genes cannot compete and within a few serial passages in drug free medium with their wild type relatives, they cease to exist [41].
It is the consensus of the pharmacologically oriented microbiologists that with the use of non-toxic modulators of efflux pump activity in combination with antibiotics, to which the initial strain is resistant, will render the organism fully susceptible to the antibiotic. This consensus must be qualified to “provide of course that the initial resistance was due entirely to efflux pump activity”. However, if mutations have taken place, even if efflux is still evident, this form of combinational therapy will probably fail. Because therapy of some infections is prolonged to as long as one month (example Lyme disease) and even years (therapy of an MDR TB infection), in all probability, the clinical bacterial isolate will express some efflux and mutations. Consequently, the ability to determine the contributions made by efflux and mutations for the antibiotic resistance of the clinical isolate may prove useful for the selection of appropriate therapy. The methods presented in this review are directed at this potential need and should be considered for introduction into general bacteriology laboratories.
Martins A is supported by SFRH/BPD/81118/2011 from Fundação para a Ciência e a Tecnologia, Portugal.
GS was supported by TÁMOP-4.2.1/B-09/1/KONV-2010-0005-Creating the Center of Excellence at the University of Szeged supported by the European Union and co-financed by the European Regional Fund and TÁMOP- 4.2.2/B-10/1-2010-0012 project: "Broadening the knowledge base and supporting the long term professional sustainability of the Research University Center of Excellence at the University of Szeged by ensuring the rising generation of excellent scientists" European Union grant.
The authors acknowledge the support of the Szeged Cancer Foundation.