Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Research Article - (2013) Volume 2, Issue 4
The pigment content (Chlorophyll-a, Chlorophyll-b and Total Chlorophyll) of dominant macrophytes was investigated in this present study along with subsequent analysis of relevant physico-chemical variables. In this study, the relationship between the pigment content of dominant species and the ambient environmental parameters was established. In general, Chlorophyll-a, Chlorophyll-b and Total Chlorophyll contents varied from 1.5 ± 0.1 µg/ml to 3.7 ± 0.5 µg/ml, 0.3 ± 0.2 µg/ml to 1.7 ± 0.5 µg/ml and 2.1 ± 0.9 µg/ml to 4.1 ± 0.5 µg/ml respectively with highest values during summer coinciding with increasing water temperatures. Chlorophyll-a and Chlorophyll-b and Total Chlorophyll levels were positively correlated (P<0.05) with water temperature and negatively correlated with nutrient salts such as nitrate, ammonia and total phosphorus. Maximum pigment content occurred in submerged macrophytes, whereas minimum levels were obtained in rooted- floating macrophytes. The analysis of Chlorophyll A, B and Total in the macrophytes indicated the presence of species specificity as well as of seasonal dynamics of these pigments. Statistical analysis computed among environmental parameters and pigment content suggests the potential role played by the abiotic parameters on the pigment content of macrophytes.
<Keywords: Pigment content; Seasonal dynamics; Submerged macrophytes
Aquatic macrophytes which are considered to be the photosynthetic organisms of freshwater habitats are central to the function of fresh water ecosystems [1]. Macrophytes are a potential source of food and fodder for humans and bovine population, besides serving as a base of aquatic food chain [2-6]. In order to evaluate the food potential of macrophytes and estimation of their nutritional value, the knowledge of their chemical composition is of paramount importance [5,7,8]. It is well known that under the influence of abiotic factors such as light, temperatute, sediment composition and water chemistry, macrophytes can modify their metabolic responses (photosynthesis and growth rates) and levels of proximate constituents [9-12] thus necessitating the determination of biochemical ingredients of the tissues produced under each ecological condition. Light and temperature act together in affecting growth, morphology, photosynthesis, chlorophyll composition and reproduction of macrophytes [13-18]. The biological functions such as photosynthesis, physiology, etc. in macrophytes are dependent on chlorophyll a (chl a), a key photosynthetic pigment. Consequently, changes in chl a can indicate plant growth [19]. or disturbances from stressors [20]. It is believed that the distinct variation in pigment content as well as specific features of the pigment complex ensure adaptation of macrophytes to light conditions in water stratum [17,18,21].
The present study aims to analyze the seasonal variations and comparative analysis in the pigment content of aquatic macrophytes characterized by different degree of submergence (emergent, rooted floating, free floating, and submerged leaves) with respect to relevant hydrological parameters (like surface water temperature, transparency, nutrients (ammonia, nitrate and phosphate).
Study site
Geographical the lake is situated at an altitude of 1580 m (a.m.s.l) lying between 34°16´24.67? - 34° 20´26.26? N latitudes and 74° 33´41.42? - 74o 44´02.90?E longitudes of Kashmir Valley. The oxbow type lake is mono- basined, elliptical in shape and is of fluviatile in origin, formed by the meandering of River Jhelum. Various rivers and streams which contribute their might draining to the Wular lake are River Jhelum, Erin Nallah, Madhumati Nallah, Ferozepur Nallah, Sukhnag Nallah, Ningal Nallah, Ajus Nallah, etc. and the only exit (outlet) of the lake is River Jhelum. The macrophyte dominant lake has well developed littoral zone. Extended littorals in general, are dominated by (i) reeds like Phragmites australis, Typha angustata and Sparganium ramosum, (ii) rooted floating-leaf types like Nymphoides peltatum, Trapa natans and Nymphaea mexicana, and (iii) freefloating types dominated by Salvinia natans, Azolla sp. and Lemnids, all growing together intermixed and forming a complex physiognomy. The slightly deeper zone has abundant growth of submergeds like Ceratophyllum demersum, Potamogeton crispus and Potamogeton lucens forming meadows at various places. The littorals towards the north-eastern side are densely populated with Salix plantation which is eating the vitals of the lake.
Sampling and analysis of pigment content
Seasonal sampling of aquatic plants was carried out between March, 2011 to November, 2011 spelling three seasons. The aquatic plants were collected according to their availability from nine sampling locations within the Wular lake, in order to determine the chlorophyll content of these macrophytes (Figure 1). These aquatic plants were harvested in a lush - green condition using quadrants of 0.25 m2 in an area [22,23] from different areas within each stand to form three representative samples for each species. The harvested materials were placed in polyethylene bags and transported to the laboratory in cool boxes (temperature 4°C) where they were incised to include only leaves. These were then washed thoroughly and moisture was drained before being analyzed for chlorophyll content.
Chlorophyll content (Chl a, Chl b and Total Chlorophyll) of macrophytes was determined Spectrophoto-metrically, in acetone extracts prepared from fresh samples using standard methods [24,25]. Acetone extract of the macrophytes was prepared by macerating 1 g fresh weight of plant leaves with 80% aqueous acetone using a mortar and pestle. The decanted suspension was centrifuged for 3 minutes at 1320 r.p.m. After centrifugation, the upper green clear solution was decanted from the colorless residue and then made up to 10ml with 80% acetone in 10 ml test tubes. The solution was then subjected to centrifugation at 10,000 r.p.m for 10 minutes. The absorbance of the solution was determined using a spectrophotometer (Systronics-116) at wavelengths of 665 and 649 nm respectively. The results were expressed as μg/ml.
Chlorophyll ‘a’ (μg/ml)=11.63 (A665)-2.39 (A649)
Chlorophyll ‘b’ (μg/ml)=20.11 (A649)-5.18 (A665)
Total Chlorophyll (μg/ml)=6.45 (A665) +17.72 (A649)
Analysis of hydrological parameters
Surface water samples were collected from all the selected stations in clean TARSON bottles for analysis of hydrological parameters like surface water temperature, transparency, nitrate, ammonia and phosphate on seasonal basis. The temperature of each water sample were measured at the field by means of a mercury thermometer, while as transparency in water was measured with Secchi disc. Other water quality parameters such as nitrate, ammonia and phosphate, which require photometric measurements, were done according to the standard methodology outlined in APHA [26] by using model spectrophotometer (Systronics-116) on the same day in Aquatic Ecology Laboratory, Department of Environmental Science, University of Kashmir-190006, India.
Statistical analysis
All the data were expressed in terms of mean ± S.D (standard deviation) and range. In addition, Pearson correlation coefficient (r) was also computed between the pigment content of the macrophytes and environmental parameters. Descriptive statistics were calculated using StatSoft Statistica 7.0 statistics software.
Seasonal variation in hydrological parameters
Maximum temperature was recorded in summer (25.3 ± 7.9°C) and minimum in autumn (14.4 ± 7.3°C). Ammonia and Total Phosphorus exhibit same seasonal patterns with autumn depicting higher values (123 ± 46.7 and 194 ± 61.1) and summer depicting lower values (69 ± 32.7 and 124 ± 53.9), respectively. The seasonal mean values of nitrate in the present investigation varied between 299.8 ± 179.7 μg/l and 482.6 ± 214.8 μg/l in summer and spring respectively. In contrast maximum transparency values (0.9 ± 0.3 m) were obtained in autumn against a minimum (0.5 ± 0.1 m) being recorded in spring (Table 1).
| Parameter (units) | Spring±S.D | Summer±S.D | Autumn±S.D | Mean±S.D |
|---|---|---|---|---|
| Water temperature (°C) | 15.3±3.7 | 25.3±7.9 | 14.4±7.3 | 18.3±4.9 |
| Nitrate(µg/l) | 482.6±214.8 | 299.8±179.7 | 431.85±167 | 404.8±77 |
| Ammonia(µg/l) | 99±31.3 | 69±32.7 | 123±46.7 | 97±22.1 |
| Total phosphorus(µg/l) | 173.7±13.4 | 124±53.9 | 194±61.1 | 163.9±29.4 |
| Transparency (m) | 0.5±0.1 | 0.7±0.2 | 0.9±0.3 | 0.7±0.2 |
Table 1: Seasonal variations (Mean±S.D) of some physico- chemical parameters in surface waters from Wular lake
Seasonal variation in pigment content
Perusal of data on the pigment content of macrophytes revealed significant difference for Chlorophyll-a, Chlorophyll-b and Total Chlorophyll in the different groups of macrophytes collected from Wular lake. The levels of Chlorophyll-a, the primary photosynthetic pigment in macrophytes in the present study varied from 1.5 ± 0.1 μg/ml to 3.7 ± 0.5 μg/ml. The values were highest in Ceratophyllum demersum (3.7 ± 0.5 μg/ml) and lowest in Azolla sp. (1.5 ± 0.1μg/ml). The Chlorophyll-b content recorded its highest value in Ceratophyllum demersum where the data observed (1.7 ± 0.5 μg/ml) was succeeded by Potamogeton crispus with a value of (1.6 ± 0.5 μg/ml). The lowest Chlorophyll-b values were obtained in Azolla sp. having a value of (0.3 ± 0.2 μg/ml).The Total Chlorophyll content also recorded its highest value in Ceratophyllum demersum (4.1 ± 0.5 μg/ml) followed by Lemna minor with a value of 4.0 ± 0.7 μg/ml, decreasing to a lowest of 2.1 ± 0.9 μg/ml in Azolla sp. (Table 2).
| Plant | Species | Chl.a (Mean ± S.D) | Chl.b (Mean±S.D) | Total Chl. (Mean±S.D) |
|---|---|---|---|---|
| Emergent | Phragmites australis | 2.2 ± 0.3 | 0.7 ± 0.3 | 2.4 ± 0.8 |
| Polygonum hydropiper | 2.0 ± 0.6 | 0.4 ± 0.2 | 2.2 ± 0.6 | |
| Rootedfloating | Nymphaea mexicana | 2.8 ± 0.4 | 1.2 ± 0.5 | 3.7 ± 0.8 |
| Nymphoides peltatum | 2.4 ± 0.5 | 0.6 ± 0.4 | 2.6 ± 0.5 | |
| Submerged | Potamogeton crispus | 3.1 ± 0.6 | 1.6 ± 0.5 | 3.5 ± 0.6 |
| Ceratophyllum demersum | 3.7 ± 0.5 | 1.7 ± 0.5 | 4.1 ± 0.5 | |
| Free floating | Lemna minor | 2.8 ± 0.4 | 1.4 ± 0.6 | 4.0 ± 0.7 |
| Azolla sp. | 1.5 ± 0.1 | 0.3 ± 0.2 | 2.1 ± 0.9 |
Table 2: Mean values of pigment content found in macrophytes harvested from Wular lake Note: Chl.a = Chlorophyll a, Chl.b = Chlorophyll b, Total Chl. = Total Chlorophyll and S.D = standard deviation of the mean
The macrophytes collected during different seasons exhibited distinct seasonal profile in the pigment content. Significant changes were seen in all the macrophytes except that of Ceratophyllum demersum and Potamogeton crispus where slight fluctuations were noticed. In the present work, maximum pigment content was observed during summer season, the period of active growth characterized by optimum nutrient load and temperature. The lowest levels of Chlorophyll-a and Chlorophyll-b were obtained during the period of less vigorous growth in autumn, and they increased considerably during the period of sprouting and early growth stages of macrophytes in spring and were highest in summer. However, relative content of Total Chlorophyll in the macrophytes under study was notably lower in spring season than in autumn (Figures 1-4).
Figure 1: Map showing study area and sampling stations.
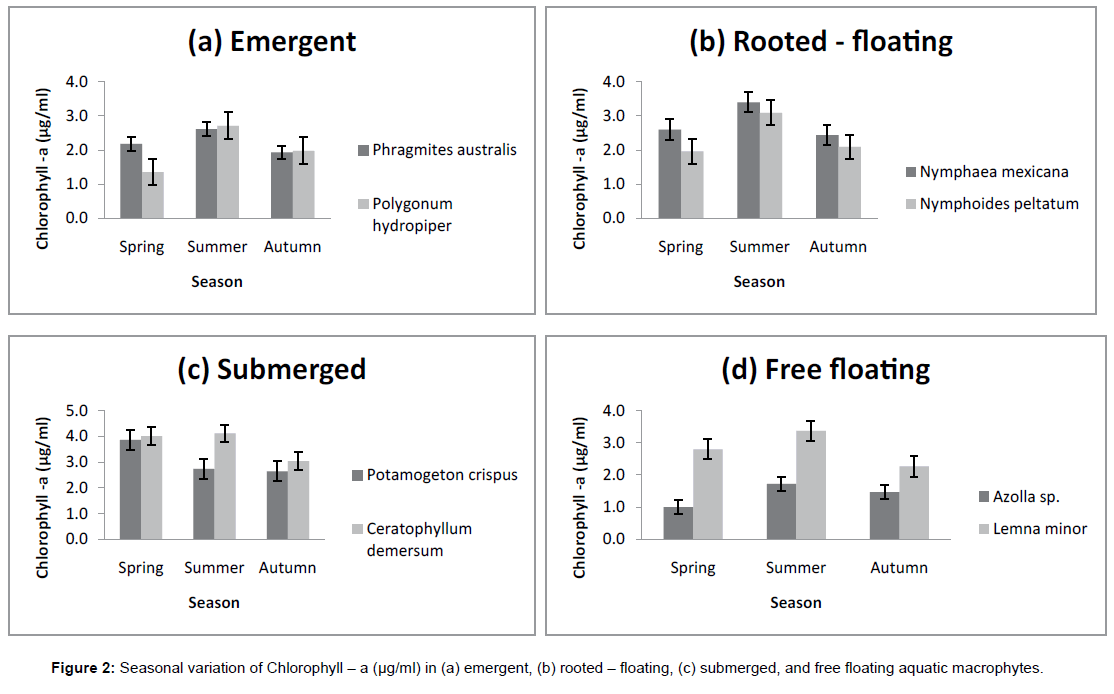
Figure 2: Seasonal variation of Chlorophyll – a (μg/ml) in (a) emergent, (b) rooted – floating, (c) submerged, and free floating aquatic macrophytes.
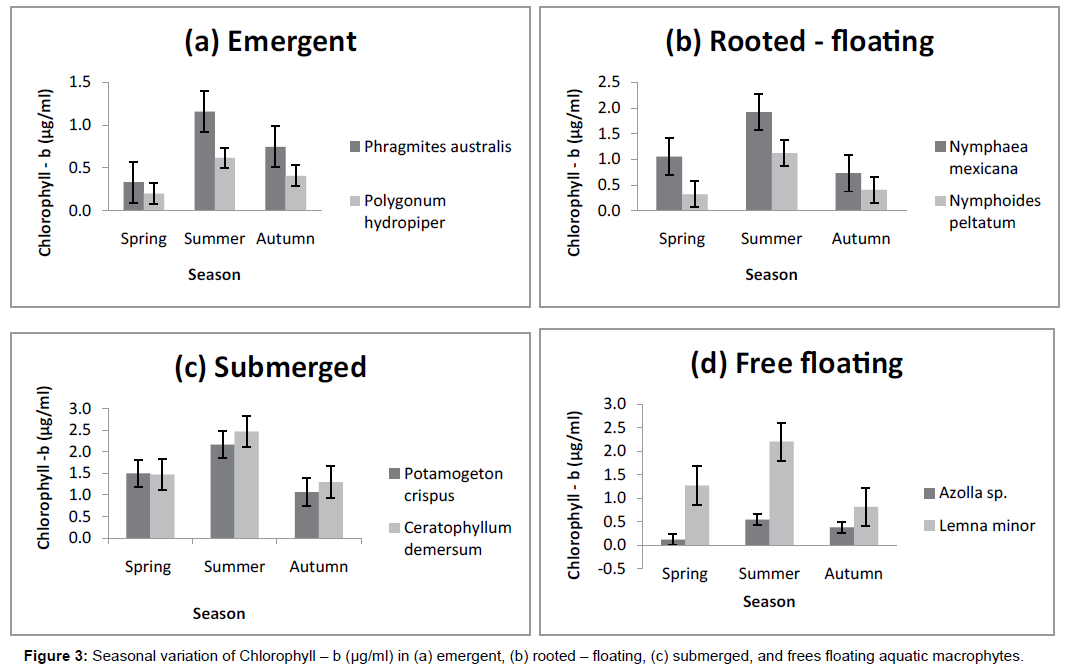
Figure 3: Seasonal variation of macrophytes – b (μg/ml) in (a) emergent, (b) rooted – floating, (c) submerged, and frees floating aquatic macrophytes.
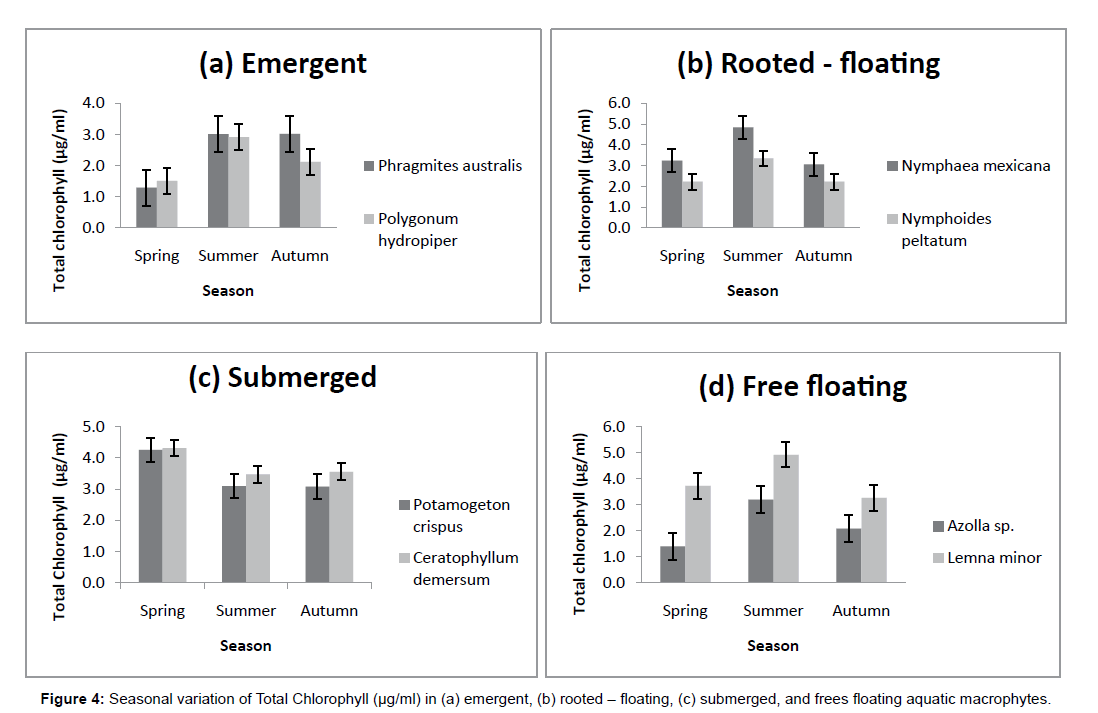
Figure 4: Seasonal variation of Total Chlorophyll (μg/ml) in (a) emergent, (b) rooted – floating, (c) submerged, and frees floating aquatic floating.
In general submerged macrophytes represented in the present study by Ceratophyllum demersum and Potamogeton crispus were characterised by high pigment content.While as lower pigment levels were obtained in emergents and rooted- floatings. Chlorophyll-a and Chlorophyll -b and Total Chlorophyll showed highly significant positive correlation (r=0.96, 1.00 and 0.95, P<0.05) with water temperature and negatively correlated with nutrient salts such as nitrate, ammonia and total phosphorus. However, transparency showed a non-significant positive correlation (r=0.23, P=0.05) with Total Chlorophyll and and negative correlation with Chlorophyll-a and Chlorophyll-b (Table 3).
| Water Temperature | Transparency | Nitrate | Ammonia | Total Phosphorus | |
|---|---|---|---|---|---|
| Water Temperature | 1.00 | ||||
| Transparency | -0.07 | 1.00 | |||
| Nitrate | -0.94 | -0.27 | 1.00 | ||
| Ammonia | -0.93 | 0.44 | 0.74 | 1.00 | |
| Total Phosphorus | -0.98 | 0.28 | 0.85 | 0.98 | 1.00 |
| Total Chlorophyll | 0.95* | 0.23 | -1.00* | -0.77* | -0.87* |
| Chlorophyll a | 0.96* | -0.35 | -0.81* | -1.00* | -1.00* |
| Chlorophyll b | 1.00* | -0.06 | -0.95* | -0.92* | -0.97* |
Table 3: Correlation coefficients between environmental variables and pigment content of macrophytes of Wular lake *Correlation at 0.05
The analysis of our data revealed some distinct trends in the pigment content in the fresh water macrophytes sampled during different seasons. In the present work, maximum pigment content of the macrophytes was observed during summer season; the period characterized by optimum nutrient load and water temperature. Chlorophyll-a and Chlorophyll-b contents were significantly lower in autumn, Total chlorophyll was the only exception. The trend may be attributed to the positive role of water temperature in promoting greater chlorophyll concentration and photosynthesis [13-16]. The summer fall in the amounts of nutrients (nitrate, ammonia and total phosphorus) is attributed to the greater photosynthetic activity of macrophytes especially submergeds leaving very small quantities of such nutrients in water [27]. This is confirmed from the significant positive correlation between Chlorophyll-a, Chlorophyll -b and Total Chlorophyll and water temperature and negative correlation with nutrients (nitrate, ammonia and total phosphorus).The lower quantity of chlorophyll pigment in macrophytes in other seasons reflects their adaptation to decreased water temperature [28]. Our results are in accordance with the published data [29,30]. Thus the active period of pigment synthesis in macrophytes coincides with the increase in water temperature and decrease in nutrient load.
The macrophytes as a whole were characterized by low chlorophyll pigment content per fresh weight basis in comparison to the terrestrial plants of middle latitudes [31]. The comparative analysis of data (Table 2) showed appreciable variations in the pigment content among different groups of macrophytes with different degree of leaf submergence (emergent, floating, free floating and submerged). In emergent and rooted- floating macrophytes, the low chlorophyll pigment content might be attributed to their existence at high irradiance [17]. In rooted- floating macrophytes the leaves are perpendicular to incident radiation. As a consequence of which they contain lower pigment content compared to submerged aquatic plants. In submerged macrophytes, the leaves are almost vertically oriented [17]. As a result, they do not shade each other resulting in higher pigment content in them. In the submerged leaves, in contrast to the emergent and floating ones, an increase in the photosynthetic rate of a single chloroplast as well as their existence in water under low light conditions results in an increase in the pigment content in their chloroplast. Our results further gain support from the findings of [17,32] who also reported higher pigment content in submerged aquatic plants. The free floating macrophytes represented in our study by Azolla sp. and Lemna minor contain higher pigment levels in contrast to emergent and rootedfloating macrophytes is due to their presence of homogenous type of mesophyll structure [14,17] suggested that within individual plants, total chlorophyll can vary significantly in response to light, with greatest chlorophyll concentration at the basal (darkest) portion of the plant. The normal life activity is ensured by low pigment content in plants adapted to high radiation. One of the advantages in having the low pigment content is that it prevents cell damage caused by photooxidation [17]. This regularity was observed for desert and highmountain plants existing under conditions of high light intensity [31].
It is evident from the study that different groups of macrophytes showed distinct profile in their seasonal pigment content. The results obtained in our study establish that physico-chemical parameters have profound influence on the pigment content of aquatic plants. Significant correlation values computed among the environmental parameters and pigment content suggest that the abiotic parameters seem to have potential role in photosynthetic pathways of macrophytes.
The authors are highly grateful to Head, Department of Environmental Science, University of Kashmir, Hazratbal, Srinagar-190006 for providing fund and facilities for smoothly conducting of research work.