Journal of Glycobiology
Open Access
ISSN: 2168-958X
ISSN: 2168-958X
Review Article - (2012) Volume 1, Issue 1
Keywords: Candida albicans; Immune sensing; Cell wall; Mannans; Glucans; Dimorphism
Candida albicans is a dimorphic fungus that normally colonizes mucosal tissues, forming part of the microflora. Its population is normally controlled by the mucosal microenvironment and the bacterial flora; although changes in any of these factors may lead to fungal overgrowth and tissue damage. The host immune system is capable to control such infection, and in immunocompetent patients the disease is restricted to superficial tissues. However, when the host experiences a temporal or permanent immunosuppression the fungus has the potential to cause systemic infections that are associated to high mortality rates [1].
Microbial pathogens contain pathogen-associated molecular patterns (PAMPs) that allow the innate immune system, via pattern recognition receptors, to recognize them as non-self molecules, triggering a cascade of molecular events that will end with a protective response against the invader. For the case of C. albicans, and other fungal pathogens, most of the PAMPs are found at the cell wall (Figure 1), as this protective shield is composed of molecules that are not synthesized by host cells [2]. However, intracellular components such as DNA can be recognized by the innate immune system [3].
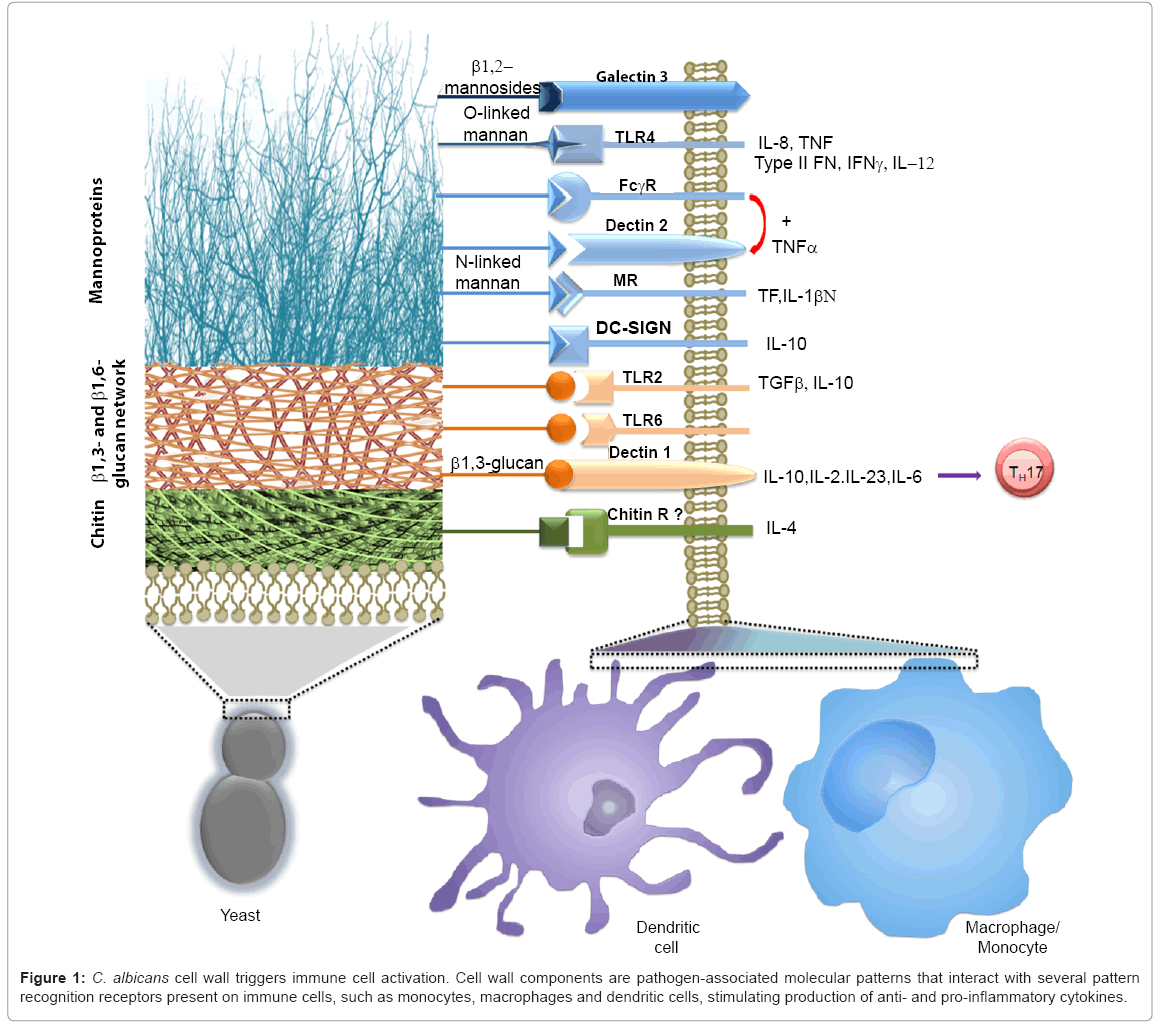
Figure 1: C. albicans cell wall triggers immune cell activation. Cell wall components are pathogen-associated molecular patterns that interact with several pattern recognition receptors present on immune cells, such as monocytes, macrophages and dendritic cells, stimulating production of anti- and pro-inflammatory cytokines.
The C. albicans cell wall is organized in two layers composed of different polysaccharides, and this organization is relevant for the fungus immune sensing (see below). Chitin and b-glucans are the main components of the skeletal inner layer of the cell wall, closer to the plasma membrane. These polysaccharides are not exposed at the cell surface, with exception of the chitin present at the primary septum and the budding scars [4]. Chitin represents about 1-2% of the yeast cell wall dry weight [5], and from this, about 5% is in the deacetylated form, i.e., as chitosan [4]. In hyphae chitin is more abundant than in yeast cells, contributing with 4-6% of wall dry weight [6,7]. The b1,6-glucan represents about 20% of cell wall dry weight, and is covalently linked to chitin and b1,3-glucan, working as a molecular bridge between these two cell wall components [5]. Cell walls from hyphae contain three times less of this polysaccharide than yeast walls [8,9]. The b1,6-glucan is exposed enough at the yeast cell wall surface to be recognized by antibodies, but not in hypha walls, where FITC-conjugated anti-b1,6- glucan antibodies are barely bond to this polymer [9]. The b1,3-glucan is one of the most abundant cell wall components, contributing with 40% of the yeast cell wall dry weight, but the content of this polymer increases twice in hypha cell wall [5,8]. This glucose polymer can directly interact with cell wall proteins, b1,6-glucan and chitin, thus it was proposed that works as a molecular scaffold to bring together most of the cell wall components [5]. As part of the inner cell wall layer, b1,3-glucan is not normally exposed at the cell surface, being buried underneath the thick outer layer composed of mannoproteins, and being only exposed at the budding scars [10-12]. The mannoproteins represent 35-40 % of the cell wall dry weight, and mannans about 40% of total cell wall polysaccharide content [5,13]. Glycolipids are a minor cell wall component of both yeast and hyphae cell wall (1-7% cell wall dry weight) [13].
Immune sensing of cell wall polysaccharides
Chitin and Chitosan: Chitin is a polymer of b1,4-GlcNAc that is essential for cell viability [14], and thus far attempts to generate mutant cells lacking this cell wall component have been unsuccessful. Its synthesis has been thoroughly reviewed before [15] and is performed by four chitin synthases: Chs1, Chs2, Chs3, and Chs8 [6,7,14,16,17]. Chs1 is in charge of the primary septum elaboration and contributes to the synthesis of chitin found at the lateral wall [14]. This is the only chitin synthase essential for the C. albicans viability, stressing the importance of the primary septum elaboration for cell physiology, division, and integrity [14]. Chs3 and Chs8 synthesize short- and long-chitin microfibrils, respectively [18], with Chs3 synthesizing about 85% of the chitin present in the cell wall of both yeasts and hyphae [16,19]. Chs2 is a hypha-specific enzyme responsible of the elaboration of about 40% of chitin found in this morphology [6]. The biosynthesis of chitosan in C. albicans has not been yet characterized, but the genome contains the ortholog of Saccharomyces cerevisiae CDA2, a chitin deacetylase involved in chitosan production [20].
The interaction between chitin and immune cells is thus far not well understood, and is currently an active research area. We have recently demonstrated that purified chitin from either C. albicans or S. cerevisiae cell wall is not able to stimulate production of pro- and anti-inflammatory cytokines by human peripheral blood monocytes (PBMCs) [4,21], nor activation of the complement alternative pathway [22]. Moreover, chitin can block the proper recognition of C. albicans by human PBMCs in a dectin-1-dependent mechanism, but this is independent on Mincle, TLR2 or TLR4 [4]. However, other groups have shown that chitin stimulates pro- and anti-inflammatory cytokine production in different cell types [23-26], but those effects are dependent on particle size and phagocytosis. Furthermore, the particle size seems to be relevant for eosinophilia stimulation [27], and for the kind of receptors involved in chitin sensing: small chitin particles are phagocytosed and recognized via TLR2, dectin-1, and mannose receptor, while non-phagocytosed chitin stimulates cytokine production via TLR2 and dectin-1 [25]. Despite these advances in the area, there are not clear evidences about the receptor(s) involved in chitin sensing. Thus far there are some candidates that are currently under study: the C-type lectin NKR-P1 receptor recognizes chitooligomers and this ligand-receptor interaction triggers activation of natural killer cells [28]; RegIIIg (HIP/PAP) is also a C-type lectin expressed in the neutrophil-like Paneth cells of the small intestine [29], FIBCD1- a calcium-dependent acetyl group-binding receptor that is also expressed in the gastrointestinal tract [30]; TLR2, Dectin-1 and mannose receptor [4,24-26,31,32].
The relevance of chitosan during C. albicans immune sensing has not been yet analyzed; however, it has been recently shown that this deacetylated β1,4-glucosamine polymer strongly activates the inflammasome in macrophages, stimulating production of IL-1β [33]. As with chitin sensing, the observed effect with chitosan was dependent on particle size and phagocytosis [33].
β1,6-Glucan
This is a short and amorphous polysaccharide, with most of the monosaccharides forming a linear structure and about 7% of them contain branching points [34]. The encoded products by KRE1, KRE9, KRE6 and SKN1 are involved in the biosynthesis of this polysaccharide, but the latter is specifically upregulated during hypha transition [35-37], which suggest that the biosynthetic machinery involved in elaboration of β1,6-glucan may be different in yeast cells and hyphae. Some of these enzymes are essential for C. albicans viability [36,37], underlining the importance of this polysaccharide for cell wall organization. Thus far the nature of the immune receptors involved in recognition of this cell wall component are unknown, but there are strong evidences indicating the immune system recognizes it: β1,6-glucan potentiates the immunoglobulin E-mediated histamine release from blood leucocytes [38], causes abnormal phenotypes during differentiation of human PBMCs to dendritic cells, and the latter have modification in the normal functions [39]. Furthermore, data indicate that this polymer can be involved in the stimulation of chemokines by human PBMCs [9], mediates phagocytosis, and production of reactive oxygen species in human neutrophils [40]. Finally, β1,6-glucan also participates in the complement activation, in a properdin-dependent way, being C3b/C3 deposited more readily on β1,6-glucan than on β-1,3-glucan [22,40].
β1,3-Glucan: The synthesis of β1,3-glucan has been extensively reviewed before [41]. The β1,3-glucan synthase is a multiprotein complex composed of Fks1 and Gsl1 [42], and this enzyme activity is regulated by Rho1, an essential small GTPase of Rho family that inhibits the biosynthetic activity via binding to Fks1 [43]. The linkage between β1,3- and β1,6-glucans is likely to be performed by Phr1 and Phr2, GPI-anchored cell wall glycosidases [44].
This polysaccharide is recognized by dectin-1, a C-type lectin expressed in myeloid cells [45,46], and TLR2 [47]. It was originally thought that recognition of β1,3-glucan was a minor pathway contributing to cytokine production by PBMCs [47] and had no role in the host response mechanisms against C. albicans [48]; however, this information was further revisited and nowadays it is well known that recognition of this cell wall polymer triggers a strong activation of immune cells and is essential for the anti-C. albicans immune control in both mice and humans [49]. Interestingly, it has been found that some mice strains show differences in the ability to interact with β1,3- glucan, and expression of two isoforms of dectin-1 are responsible of this [50]. Dectin-1 has an extracellular C-type lectin like domain and stalk region, followed by a single transmembrane pass and a tyrosinebased activation-like domain at the cytoplasm. The dectin-1B isoform lacks the stalk region and this compromises the ability of the receptor to bind β1,3-glucan at 4°C, but unexpectedly, triggers the production of more TNFa than dectin-1 [50]. These effects might be result of different abilities to interact with other surface receptors, such as TLR2 and CD63 [50]. The binding of dectin-1 with TLR2 is important to promote production of TNFa and IL-12 [51, 52], but interaction dectin-1-Syk kinase, and further activation of CARD9, are required for a TLR2-independent pathway that stimulates production of IL-2, IL- 10, IL-23 and IL-6 [53, 54]. Furthermore, dectin-1 is also involved in the phagocytosis and killing of C. albicans by polymorphonuclear cells [55], and stimulates production of IL-1β via a phagocytic process and activation of NLRP3 inflammasome [56].
Initially the role of β1,3-glucan during immune sensing of C. albicans was considered as a minor pathway, because mannans were demonstrated to contribute to most of the cytokine production stimulated by this organism. In addition, mannans are forming the outer cell wall layer, covering the inner components, including β1,3- glucan. However, it was demonstrated that exposure of β1,3-glucan at the surface by heat inactivation [11], antifungal drugs [12,57] or mutations that affect the mannan layer [58-60] stimulates a strong cytokine production and phagocytosis. The β1,3-glucan is normally exposed at the budding scars, and has been demonstrated that this is enough to trigger C. albicans phagocytosis and production of reactive oxygen species through dectin-1 activation [10]. However, the participation of dectin-1 seems to be restricted to yeast cells, as hyphae do not divide through a budding process, and thus do not have β1,3- glucan exposed on the cell surface, and do not trigger activation of immune cells via dectin-1 [10]. This elegant mechanism of immune evasion that C. albicans possesses can be dismantled by treatment with caspofungin: in vitro and in vivo observations indicate that subinhibitory concentrations of caspofungin drive unmasking of β1,3- glucan in hypha, being this cell wall polymer exposed enough to be recognized by dectin-1 [57].
It has been recently shown that the immune response triggered by dectin-1 requires a clustering mechanism of the receptor, forming a phagocytic synapse that allows the immune cells to discriminate between direct microbial contact and soluble ligands [61].
Mannoproteins: The cell wall mannoproteins are synthesized in the endoplasmic reticulum and then transported to the cell wall by the secretory pathway. Once the proteins are translocated to the endoplasmic reticulum lumen, they may be post-translationally modified by addition of oligosaccharides rich in mannose residues (mannans) that can be covalently attached to Ser/Thr residues via an ester bond (O-linked mannans), or to Asn residues through an amide link (N-linked mannans). The synthesis of these molecules has been extensively reviewed elsewhere [2], beginning in the endoplasmic reticulum and finalising in the Golgi complex. Addition of N-linked mannans is performed by the oligosaccharyl transferase complex that transfers the preformed N-linked glycan core from a dolichol-based donor to Asn residues within the sequon Asn-X-Ser/Thr, where X can be any amino acid except Pro [2]; whereas the first mannose residue of the O-linked mannans is attached to proteins by members of the PMT gene family [62]. Then, mannans undergo an elongation process in the Golgi complex, where they get the final structure: the O-linked mannans may be composed of up to five a1,2-mannose residues [63], while N-linked core mannan is modified with the outer chain that can contain up to 200 mannose units [64]. This N-linked mannan outer chain has a backbone of a1,6-mannoses that is modified with branches of a1,2-mannose residues that resemble the O-linked mannans [64]. This branches can be terminated with either a1,2-, a1,3- or b1,2- mannose residues. Furthermore, both N- and O-linked mannan are modified with mannose residues attached to the oligosaccharide through a phosphodiester bond, named phosphomannan [64,65]. Phosphomannan may work as a molecular scaffold to add up to 14 b1,2-mannoses to the N-linked mannans [66], by action of Bmt2, Bmt3, and Bmt4 [67].
Recognition of O-linked mannans is a relatively minor pathway for cytokine stimulation in macrophages and PBMCs, and depends on interaction of this wall component with TLR4 [47]. This has been confirmed in a TLR4-deficient mouse model of disseminated candidiasis, where mice lacking this receptor are equally susceptible to the systemic disease as the wild type control animals [68].
The immune recognition of N-linked mannans is more complex than the one against O-linked mannans, as the former are composed of mannose residues linked with different glycosidic bonds. The mannose receptor is a C-type lectin that recognizes a-linked mannoses within branched oligosaccharides, such as the N-linked mannans [69]. This may explain why this receptor does not recognize O-linked mannans, even though they resemble a branch of the N-linked mannan outer chain. This receptor, present on the surface of PBMCs, macrophage and dendritic cells, is indeed involved in cytokine stimulation and phagocytosis through N-linked mannan recognition [47]. Once this role was characterized, it was proposed as the main pathway to trigger cytokine production upon C. albicans sensing, as mutants lacking the N-linked outer chain only stimulated 20% of cytokine produced by wild type control cells [47]. The C-type lectin DC-SIGN, expressed only at the surface of dendritic cells, is also involved in the recognition of N-linked mannans via the a1,2-branches, is equally good to bind C. albicans cells as mannose receptor, but might be higher expressed on dendritic cells than mannose receptor; thus, having a predominant role in C. albicans recognition by this type of immune cells [70]. Furthermore, the b1,2-mannose moieties can be recognized by galectin 3, and this contributes to cytokine stimulation in macrophage and discrimination between C. albicans and S. cerevisiae [71]. However, this might not be the case for human PBMCs, where yeast cells lacking b1,2-mannose residues are as good as wild type cells to induce cytokine production [58]. Phosphomannan also plays a role during C. albicans recognition by macrophages, being required for proper phagocytosis [72], and the effect of cationic antimicrobial peptides generated by immune cells [73]. Mincle is another C-type lectin expressed on the surface of macrophages that is involved in recognition of C. albicans mannans. This receptor is dispensable for C. albicans phagocytosis, but is required for a strong stimulation of pro-inflammatory cytokines [74]. Although the exact nature of its ligand is still unknown, its importance for establishment of anti-Candida defences is highlighted by the increased susceptibility of mice lacking mincle to systemic candidiasis [74]. Dectin-2, a C-type lectin expressed on the surface of macrophages and dendritic cells, also recognizes N-linked mannans, showing a preference to interact with high-mannose oligosaccharides, especially with the N-linked mannan core [75]. This receptor lacks an intracellular signaling motif, therefore is unable to transduce intracellular signals by itself. However, coupling with FcRγ is enough to overcome this problem and to induce cytokine production via Syk kinase and CARD9 [76-78]. The receptor is dispensable for the establishment of an immune response against C. albicans [77], but mice lacking dectin-2 are more susceptible to systemic candidiasis [78]. Recognition of both C. albicans hyphae and yeast cells through dectin-2 induce Th17 cell differentiation [78], and along with dectin-1 stimulate the production of IL-17 and a Th1 response [77]. Interestingly, dectin-2 has the ability to discriminate between C. albicans yeast cells and hyphae, showing a preference to bind the latter [76,79], but yeast cells stimulated the production of IL-1β and IL-23 via dectin-2, while hypha-stimulated cytokine production was partially dependent on this receptor [78].
Phospholipomannan: The glycolipid is composed of a phytoceramide associating a phytosphingosine and C(24) hydroxyl fatty acids, which works as a molecular scaffold for a linear oligosaccharide that contains up to 19 b1,2-mannose residues [80]. This glycolipid is considered as C. albicans virulence trait, as demonstrated by its ability to trigger macrophages apoptosis upon C. albicans phagocytosis [81]. Despite the importance of this glycolipid, little is known about its biosynthesis. It has been shown that the BMT gene family encodes for b-mannosyl transferases involved in the b-mannosylation of N-linked mannans, but not of phospholipomannan [82]. C. albicans phospholipomannan stimulates TNFa production by PBMCs and macrophages [83,84], and is sensed by TLR2, although loss of TLR4 and TLR6 significantly decreased the levels of TNFa stimulated by the glycolipid [84].
Interplay among pattern recognition receptors: The study of the interaction C. albicans-immune cells has been approached using mutants lacking specific cell wall components, blocking agents for specific ligands, or cell lineages lacking pattern recognition receptors. However, the cell-cell interaction process relies on a series of contacts among ligands and receptors, all working in a coordinate manner, and with the timing to allow a biological process. Thus, it is naive to conceive that the interaction C. albicans-immune cells may depend only on one ligand-receptor interaction. In addition, such kind of approach, although useful to establish the basic components during C. albicans immune sensing, pays little attention to the potential crosstalk between receptors, the recognition of one cell wall components by two different receptors at the same time, as in the case of N-linked mannans, or the co-stimulation triggered by the recognition of two different wall components at the same time.
As mentioned above, it is well known that dectin-1 works in collaboration with TLR2 for stimulation of some cytokines by b1,3- glucan (Figure 2), but it also synergizes with TLR4, TLR5 and TLR9, via Syk kinase, for cytokine production in both PBMCs and macrophages [85-87]. Furthermore, dectin-1 and DC-SIGN work in collaboration during activation of arachidonic acid cascade by zymosan [88], and can also collaborate with SIGNR1, a mannan receptor found on murine macrophages surface, during C. albicans recognition, enhancing cellular oxidative burst [89,90], and with galectin-3 for cytokine production in macrophages [91].
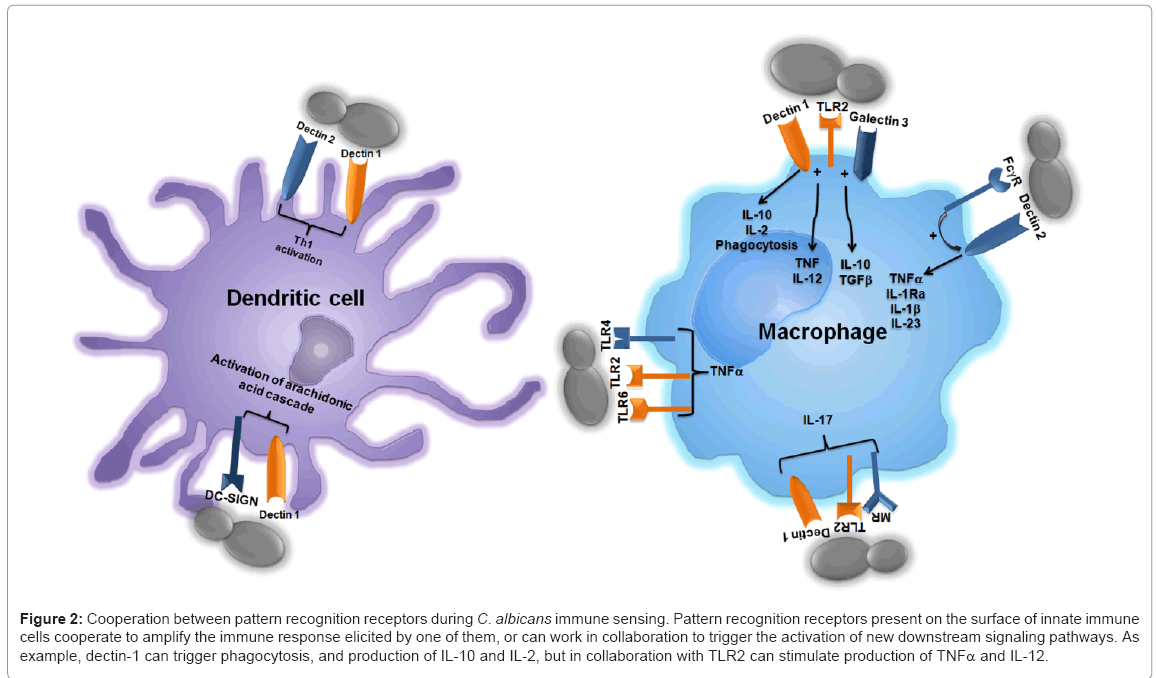
Figure 2: Cooperation between pattern recognition receptors during C. albicans immune sensing. Pattern recognition receptors present on the surface of innate immune cells cooperate to amplify the immune response elicited by one of them, or can work in collaboration to trigger the activation of new downstream signaling pathways. As example, dectin-1 can trigger phagocytosis, and production of IL-10 and IL-2, but in collaboration with TLR2 can stimulate production of TNFα and IL-12.
Mannose receptor, along with dectin-1 and TLR2 are the main receptors involved in the stimulation of IL-17 production by human PBMCs [92]. It has been recently shown that dectin-1 mediates the metalloprotease-dependent hydrolysis of mannose receptor, generating a soluble protein with the ability to still recognize N-linked mannans and cover fungal particles [93]. The biological implications of this processing are not clear yet, but are likely to modulate the mechanisms behind fungal sensing.
Future directions
The last fifteen years have experienced a significant advance in the knowledge of the molecular mechanisms behind the C. albicans immune sensing. We have now valuable information about the main fungal components and the receptors involved in the immune recognition of this pathogen, and their relative contribution for the general outcome upon sensing. Despite this knowledge, we are not close yet to have the complete information set, as there are still receptors that need to be found, such as the one responsible for chitin or b1,6-glucan recognition. As described above, there are data indicating that dectin-1 plays a major role interacting with TLR’s, but little information is known about the potential cooperation between different C-type lectins, and TLR’s with C-type lectins different to dectin-1. Furthermore, the models that can be drawn so far, may only apply to recognition of yeast cells, as hyphae are differentially sensed by immune cells. Indeed, it has been reported that yeast cells but not C. albicans hyphae can trigger cytokine stimulation by human PMBCs and macrophages, and this might be related to the inability of the latter morphology to stimulate cytokines via TLR4 [94]. However, other authors have reported that hyphae, but not yeast cells, are able to stimulate production of IL-1b via the Nlrp3 inflammasome in primary mice macrophages [95,96], a protective Th17 response [96], and an innate response in oral and vaginal epithelial cells [97,98]. Finally, once the models would be established, it remains to be determined whether they can be applied to other Candida species that are emerging as causative agents of systemic candidiasis.
H.M.M.M. wishes to thank CONACyT, PROMEP (UGTO-PTC-261) and Universidad de Guanajuato (grant 025/11) for the financial support; L.A.P.G. and D.F.D.J. are supported by a scholarship from CONACyT.