Journal of Glycobiology
Open Access
ISSN: 2168-958X
ISSN: 2168-958X
Case Report - (2014) Volume 3, Issue 1
Keywords: HIV-1; V3 loop; Coreceptor tropism; N-glycosylation; Viral entry efficiency
The infection of cells by human immunodeficiency virus type-1 (HIV-1) requires the interaction of the external glycoprotein 120 (gp120) to adherence factors and specific glycosylated receptor molecules. Viral adherence factors are cell membrane associated heparan sulfate proteoglycans (HSPG) represented by the transmembrane syndecans [1-3], followed by binding to CD4 [4] and at least one of the coreceptors CCR5 or CXCR4 [5] as specific receptors.
The gp120 envelope is highly Nglycosylated [6] and contains five variable loop regions, designated V1-V5 loop [7] with the hyper variable V3 loop as the part that binds to the coreceptors. The V3 loop is surrounded by five N-glycans (g13-17) and especially N-glycan g15, within the V3 loop, plays an important role in viral escape from neutralizing antibodies and greatly influences the efficiency of viral infection [8].
In general, the CD4 molecule is the primary receptor for HIV-1 and a binding target for gp120. One of the first functional V3 loop studies pointed out that the V3 loop is the major determinant that controls coreceptor selection. In this study, a single amino acid mutation within the gp120 V3 loop, S306R, completely converted the tropism of the virus from non-syncytia inducing (NSI) to the syncytia inducing (SI) phenotype [9]. These data strongly argued for a second HIV-1 receptor and with the identification of the two chemokine receptors CCR5 and CXCR4 as second receptors [10], NSI and SI viruses are now named R5- and/or X4-tropism, respectively. Thus, just the introduction of arginine R306 into the V3 loop was responsible for the shift to X4-tropism. With the progression in recombinant HIV-1 technology virus mutants could be generated much easier and viruses with mutations affecting the V3 loop N-glycosylation site NNT demonstrated that the g15 Nglycan operates as a protective shield against neutralizing antibodies [11,12]. It also became apparent that there are two best-fit combinations for coreceptor usage [8,13,14]. The viruses with a V3 loop sequence characterized by a low overall positive V3 loop charge (<5) and the presence of the g15 Nglycan are optimal adapted to CCR5. They also showed a performance advantage over other R5-tropic viruses that are lacking the V3 loop g15 N-glycan. As shown in Figure 1 the R5-tropic virus NL-V3A that lacks N-glycan g15 (NL-V3A, g15) was efficiently neutralized by soluble gp120 from the same virus that only differs in Nglycosylation (V3A, +g15). Here it was shown that the V3A Nglycosylated V3 loop is a strong competitor for CCR5 binding. In CCR5-specific infection, the g15 N-glycan effect is not influenced by hexadimethrine bromide (polybrene) in contrast to CXCR4-specific infections. The “best-fit” for CXCR4-binding is characterized by a high overall positively charged V3 loop (>5) and the lack of N-glycan g15 [14]. We have constructed two X4-tropic viruses lacking four of the N-glycosylation sites in the V3 region (g13, g14, g15, g17) which only differ in the KKK>RRR sequence at V3 position 304-306. Based on the RRR and KKK sequences, the KKK virus was 10-times less infectious than the RRR mutant. As a consequence, the KKK virus was efficiently neutralized by the soluble gp120-RRR when the cells had been pretreated with polybrene (Figure 1). On the other hand, when the cell membrane-associated carbohydrate structures were not covered by polybrene, infection of the KKK virus was enhanced. The same principal effect was seen with the X4-tropic NL-NPI—gp120-NNI pair. Here, the enhancing effect was significantly more distinct, compared to the RRR-KKK pair, and was also completely negated when the cell membrane associated heparan sulfate structures were cleaved off by heparinase I, while leaving the specific viral receptors, CD4 and chemokine receptors CXCR4 and CCR5 untouched (Figure 1).
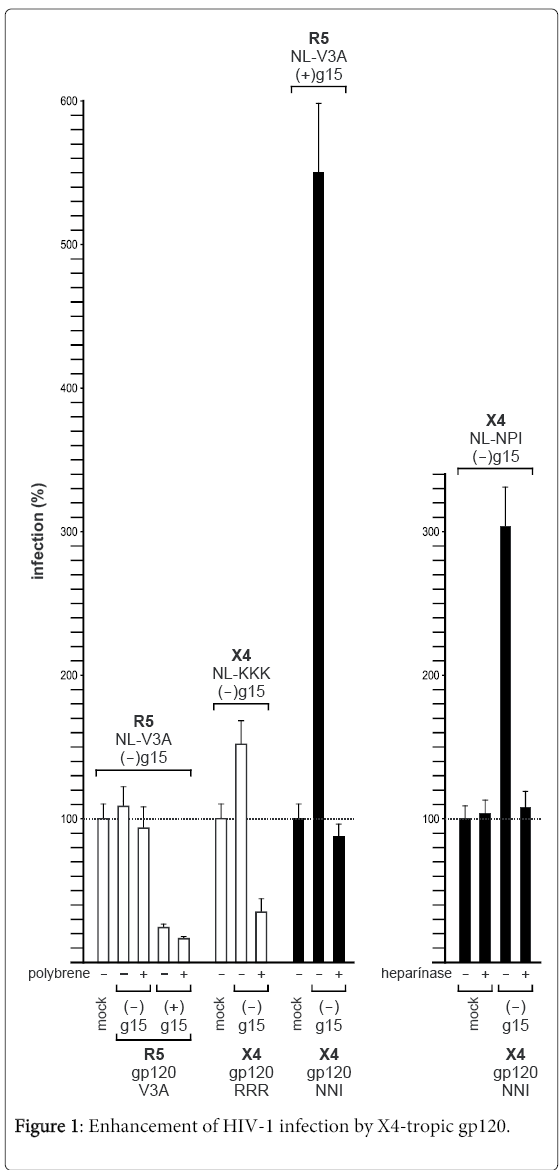
Figure 1: Enhancement of HIV-1 infection by X4-tropic gp120.
Principally, using soluble gp120 as a competitor is a new method to study viral entry without the rivalry of a second virus [15]. With this tool it is also easy to analyze viral entry in the presence of an envelope that uses the “wrong” coreceptor. In this regard (Figure 1), X4-gp120-NNI (-g15, X4 best-fit) was added to an R5-tropic infection experiment including NL-V3A (+g15, R5 best-fit). As a control, NL-V3A was tested on a variety of CXCR4-expressing reporter cell lines without any signs for CXCR4-specific infection. Therefore, NL-V3A was characterized as a highly CCR5-monotropic isolate without any X4 background activity. Interestingly, the X4-gp120-NNI enhanced the R5-NL-V3A virus by about 550%. The X4-gp120-NNI enhancing effect was completely negated when the cells were pretreated with polybrene.
The treatment of cells with polybrene and heparinase I indicates a role of heparan sulfates as a factor used by X4-gp120 to enhance infection of other X4- but more importantly other R5-tropic viruses. To our knowledge this is a new observation that might be explained, for example, by the special carbohydrate-binding properties of arginine amino acids in general. Particularly, arginine R298 is highly conserved in CCR5-adapted V3 loops and was shown to interact with carbohydrates [3]. It is therefore conceivable that X4-gp120 will mask carbohydrates and as a consequence, V3 loop based arginine amino acids of the R5viruses will be free now to bind preferentially to CCR5. The binding to and therefore masking of carbohydrate structures by X4-gp120 could also be a principle mechanism that reduces the surface net charge of the cellular membrane finally leading to virus enhancement. Using the new virus-envelope paired competitive assay we have identified this new activity of gp120, the cross-enhancing effect of X4-gp120 on R5-viruses and have found a new aspect of how the virus quasispecies can manipulate its own entry process by making target cells more permissive for its viral peers. V3 loop virus mutants were constructed as described earlier [8] and soluble gp120 was produced in HeLa-P4 cells [15,16]. For cell infection we used TZM-bl cells which stably express the HIV-1 receptors CD4, CCR5, CXCR4 [15]. The cell line enables simple and quantitative analysis of a so-called HIV-1 single round infection using the reporter protein β-gal whose expression is controlled by a HIV-1 TAT promoter. The β-gal reporter is activated by TAT protein as soon as HIV-1 has entered the cell. Expression of β-gal therefore allows the identification of single infected cells (foci) by blue-staining with 5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside (X-gal). For each virus we used a virus amount producing 100 infected cells (foci forming units) which represents the 100% infection rate. An infection rate of 0% represents 100% neutralization and rates above 100% represent enhancement of infection.
The virus amount producing 100 ffu was given to the cell layer of 10.000 TZM-bl cells in a single cavity of a 96-well microplate together with cell culture supernatants containing soluble gp120 (final conc. 7,5 ng/ml). Soluble gp120 was applied as cell culture supernatant from cells (HeLa-P4) transfected with gp160 expression vectors leading to the surface expression of the gp41/gp120 complex and finally to the release of gp120 into the culture medium. The amount of soluble gp120 in cell culture supernatants was monitored by an in-house immune-dot assay as described earlier [15]. Polybrene was used at a final concentration of 2 µg/ml in DMEM cell culture medium including 5% fetal calf serum. Cells were incubated with polybrene (Sigma-Aldrich) for 1 h and washed with phosphate buffer saline prior to infection. Heparinase I (Sigma-Aldrich) was applied to 10.000 cells at 1 unit in 50 µl phosphate buffed saline and 50 mM MgCl2.
Viruses:
NL V3A (+) g15; a mutant of the NL4-3 laboratory strain containing a R5-tropic V3 loop sequence and the NNT glycosylation site: 296CTRPNNNTIQSVRIGPGQTFYATGQIVGDIRQAHC.
NL V3A (-) g15, carrying the NNT>QNT mutation. NL-KKK, a mutant of the NL4-3 laboratory strain containing an X4-tropic V3 loop sequence: 296CTRPNNNTKKKITLGPGRVLYTTGEIIGDIRKHAC.
NL-NPI a mutants of the NL4-3 laboratory strain containing an X4-tropic V3 loop sequence lacking the NNT glycosylation site: 296CTRPGNPIRKRVGIGPGQAFHATGNIIGDIRRAQC.
Soluble gp120:
2568-958x-3-110gp-120-RRR, a V3 loop mutant from the NL-KKK virus 304KKK306>304RRR306.
2568-958x-3-110gp-120-NNI, a V3 loop mutant from the NL-NPI virus with a P312N mutation.
2568-958x-3-110Mock: Cell culture supernatant of HeLa-P4 cultures without soluble gp120.
2568-958x-3-110X4: CXCR4-monotropism
2568-958x-3-110R5: CCR5-monotropism
2568-958x-3-110(-), (+) g15: Absence or presence of N-glycan g15 within the HIV-1 gp120 V3 loop.