Cell & Developmental Biology
Open Access
ISSN: 2168-9296
ISSN: 2168-9296
Review Article - (2014) Volume 3, Issue 3
During organogenesis, the liver develops from the foregut endoderm and grows into the adjacent septum transversum resulting in the formation of the liver bud. Growth factors released from the septum transversum and the cardiac mesenchyme induce endodermal differentiation and proliferation, thus, forming the primordial liver and extrahepatic biliary structures. Endodermal precursor cells within the liver bud comprise bi-potential liver progenitor cells called hepatoblasts, which differentiates into hepatocytes and cholangiocytes. While the postnatal liver has remarkable capacity to regenerate via the proliferation of mature hepatocytes, this compensatory mechanism may be overwhelmed during states of chronic injury. Under these conditions, resident stem/progenitor cells proliferate to replace lost liver parenchyma. Significant progress has been made recently in elucidating the role of various signaling pathways and progenitor cells in liver development and regeneration. In this review, we summarize our recent understanding of progenitor cells in liver development, regeneration and repair.
<As the largest organ in the body, the liver is involved in numerous critical roles during homeostasis. The liver is organized into small hexagonal anatomic subunits, termed lobules, each consisting of a draining central vein and surrounded at their six corners by triads of hepatic portal veins, arteries, and bile ducts. Hepatocytes, which comprise over 85% of the total liver mass, reside alongside sinusoids, which carry admixed portal and arterial blood from the triads toward the draining central veins which themselves ultimately drain into hepatic veins. Hepatocytes are vital in terms of nutrient metabolism and storage, synthesis of secreted proteins, blood detoxification, and bile production amongst many key functions. Cholangiocytes or biliary epithelial cells, line the ducts that carry bile from the liver. Cholangiocytes are also involved in the absorption and secretion of water, lipids and electrolytes, thus maintaining ionic homeostasis of the bile. In addition, cholangiocytes play an important role in the local immune response by interacting with immune cells via secreted cytokines that in turn modulates the physiological functions of cholangiocytes as well as surrounding liver cells and invading inflammatory cells [1,2]. Hepatic stellate cells play a major role in liver regeneration following injury via secretion of growth factors such as Fibroblast Growth Factor (FGF), Hepatocyte Growth Factor (HGF), WNT (Wingless/Integrated), VEGF (Vascular Endothelial Growth Factor) and Insulin-like Growth Factor (IGF) [3,4]. Kupffer cells, the resident macrophages within the liver, clear aged red blood cells and pathogens present in the blood through phagocytosis. Sinusoidal endothelial cells line the capillaries in between the stacked hepatocytes within the lobule and play an important role in liver regeneration via production of growth factors such as HGF and EGF (Epidermal Growth Factor) [5].
During early stages of hepatogenesis, a portion of the foregut endoderm invades the adjacent cardiac mesenchyme and septum transversum [6,7]. Both mesoderm-derived structures induce endodermal cells to proliferate, branch, and form the glandular epithelium of the liver via a combination of complex signaling pathways. Fibroblast Growth Factor (FGF), Bone Morphogenetic Protein (BMP) and Transforming Growth Factor-Beta (TGFβ) signaling pathways regulate liver bud induction during embryogenesis (Figure 1).
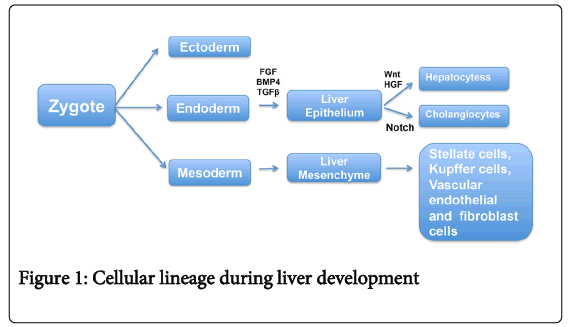
Figure 1: Cellular lineage during liver development
During this stage, hepatic endodermal cells undergo a morphologic transformation to epithelial cells, which attach to a basement membrane [7-14]. Cells within the liver bud express several hepatic genes including Albumin, Alpha-fetoprotein, and Transthyretin, along with several transcription factors, all of which are indicators of an early hepatic cell fate [7].
A subset of endodermal cells within the liver bud form a population of bi-potential progenitor cells, known as hepatoblasts, which express hepatocyte markers such as Albumin along with the biliary epithelial marker Cytokeratin-19 (Ck19) [7,8]. Additionally, putative stem/progenitor cell markers, such as Prominin-1 (also known as CD133) and CD49f, are expressed by murine hepatoblasts as early as embryonic day 12.5. FGF Receptor-2 (FGFR2)-mediated activation of AKT/β-catenin signaling via CREB binding protein (CBP) plays a critical role in the proliferation and survival of hepatoblasts during development [13,15].
During later stages of development, hepatoblasts differentiate into CK19-expressing biliary epithelial cells or HNF (Hepatocyte Nuclear Factor)-4α and Albumin-expressing hepatocytes. Gradients of growth factor signaling involving FGF, HGF (Hepatocyte Growth Factor) and TGFβ, BMP4, WNT (Wingless/Integrated) and Notch signaling pathways determine the fate of these cells toward either hepatocytes or biliary epithelial cells and are later organized into unique structures with distinct functions [10-12,16-21]. Portal fibroblasts regulate the differentiation of hepatoblasts towards a cholangiocyte phenotype and promote intrahepatic bile duct formation via Notch signaling [18-23]. Hepatoblasts at a greater distance from portal fibroblasts are thought to differentiate into hepatocytes and arrange into sheets of cells lined by sinusoidal epithelial cells and bile canaliculi. Several transcription factors such as GATA4, Hhex, HNFβ, HNF4α, HNF6, Tbox transcription factor 3, and Prox1 are known to regulate the hepatic specification of endodermal cells and later their differentiation to cholangiocytes or hepatocytes [14,24-30].
The liver has a remarkable regenerative capacity compared to other organs. Mature hepatocytes have the ability to replicate and repopulate the liver during acute injury. Mesenchymal cells are known to have a critical role in liver regeneration. Stellate cells, fibroblasts, endothelial and kupffer cells are the main types of mesenchymal cells in the liver as well as the sources of growth factors during liver regeneration [3-5]. Following acute injury, hepatocytes release mitogenic signals to mesenchymal cells and induce their activation and proliferation. Activated mesenchymal cells in turn release growth factors and cytokines such as HGF, EGF, FGF, TNFα and IL6, all of which facilitate hepatocyte proliferation [31,32]. Under normal conditions, hepatocyte proliferation ceases one week after injury and liver mass is restored within two to three weeks. Studies suggest that TGFβ signaling may play an important role in this process [32-34] (Figure 2). However, the definitive molecular mechanism(s) that regulate the termination of hepatocyte proliferation during normal liver regeneration is unclear.
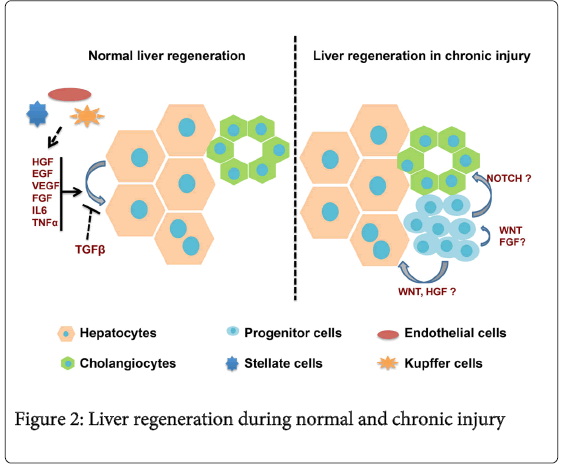
Figure 2: Liver regeneration during normal and chronic injury
During chronic injury, hepatocytes undergo ‘cellular senescence’, losing their proliferative capacity through the expression of cell cycle arrest proteins [35,36]. Studies demonstrate that Hepatic Progenitor Cells (HPCs), or oval cells, which are quiescent in the healthy liver, may be activated when the regenerative capacity of mature hepatocytes is severely impaired [37-39]. These activated postnatal HPCs express a variety of stem/progenitor markers such as EpCAM, OV6, Prominin1, CD49f, and A6 [15,40-43].
It is theorized that a stem cell niche populated by cholangiocytes, small hepatocytes, portal fibroblasts, and HPCs exists within the Canals of Hering, adjacent to the periportal regions of the liver [44]. When the liver is damaged, HPCs within this niche may become activated and differentiate into either cholangiocytes or hepatocytes (Figure 2). Intrinsic signals from the hepatic microenvironment regulate the expansion and differentiation of liver progenitor cells. FGF-mediated AKT/β-catenin signaling, for instance, modulates the presence of cells expressing the putative progenitor cell marker A6 in an experimental model of acute liver injury [45].
Whether this represents reprogramming of hepatocytes to express a progenitor marker or maturation and differentiation of progenitors toward a hepatocyte cell fate is unclear. There are lineage-tracing studies that provide evidences for hepatocyte differentiation of HPCs during injury. Recent study by utilizing a transgenic mouse model expressing inducible Cre recombinase under the control of Osteopontin gene, a known progenitor cell marker (46); demonstrated that HPCs contribute to hepatocyte mass in a choline deficient ethionine diet induced liver injury model (47). Another recent cell lineage study demonstrated that SRY-related HMG Box Transcription Factor-9 (SOX9) expressing cholangiocytes/progenitor cells are the source of hepatocytes in different experimental models of liver injury [48]. Upon transient amplification, these HPCs infiltrate along the liver plate toward the central vein and differentiate into hepatocytes to restore liver function and cell mass [47,48]. Despite all of these studies, the origin of progenitor cells and their migration during liver regeneration remains unclear [3-5]. Fibroblast progenitor cells are known to contribute to liver regeneration via activation of Hedghog signaling [49]. Macrophage-derived WNT is one of several factors, which influence the differentiation of progenitor cells towards hepatocyte lineage during liver regeneration [50]. Activated Notch signaling has been demonstrated during liver regeneration [51]. Given the known role of Notch signaling in biliary differentiation during liver development [18], it is plausible that this pathway also regulates progenitor cell differentiation into cholangiocytes during liver regeneration. Hence, cell-to-cell interactions and the liver microenvironment may influence the progenitor cell fate and thereby the extent of tissue regeneration during chronic liver injury.
Cellular homeostasis is a critical factor that determines proper organ development and regeneration. This is mainly achieved through a tight regulation of progenitor/stem cell proliferation and differentiation via a complex network of signaling pathways. Deregulation of these factors could lead to impaired development and tissue regeneration. Hence, a better understanding of the mechanisms by which liver progenitor cells interact with other liver cell types as well as the regulatory signaling pathways that get activated in response to injury is critical before implementing any cell based therapies for liver diseases.
MVN is supported by California Institute for Regenerative Medicine (CIRM) postdoctoral training grant.