Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Research Article - (2015) Volume 4, Issue 1
Oxidative stress is a key component in the development of cardiovascular diseases and chronic kidney diseases. Statins have cardio-protective activity, and previous reports have indicated that they activate nuclear factor erythroid 2-related factor 2 (Nrf2), although their molecular mechanism is unknown. Nrf2 is an oxidative stress-responsive transcription factor with a crucial role in cellular defense against oxidative stress. We investigated the molecular mechanisms of Nrf2 activation by rosuvastatin. Nrf2 activity and Nrf2-mediated antioxidant gene expression were upregulated by rosuvastatin in human umbilical vein endothelial cells. Rosuvastatin increased Nrf2 protein levels by reducing Nrf2 degradation and upregulating the interaction between Nrf2 and p21Cip1, which was inhibited by p21Cip1- targeted siRNA. Rosuvastatin-mediated activation of endothelial Nrf2 provides a possible therapeutic alternative for cardiovascular diseases.
<Keywords: Endothelial cell; p21Cip1; Nrf2; Statin; Oxidative stress
Nrf2: Nuclear Factor Erythroid 2-related Factor 2; Keap1: Kelch-like ECH-Associated Protein 1; ARE: Antioxidant Responsive Element; SOD: Superoxide Dismutase; NQO1: NAD(P) H:quinone Oxidoreductase-1; HO-1: Heme Oxygenase-1; GCLM: Glutamate-Cysteine Ligase Modulatory Subunit; Statins: 3-hydroxy- 3-methyl-glutaryl-CoA reductase inhibitors; HUVECs: Human Umbilical Vein Endothelial Cells; RSV: Rosuvastatin
The nuclear factor erythroid 2-related factor 2 (Nrf2) is an important oxidative stress-responsive transcription factor, which has a vital role in combating oxidative damage [1]. Upon activation, it becomes free from cytoplasmic sequestering and negative regulation by Kelch-like ECH-associated protein 1 (Keap1), resulting in nuclear accumulation and transactivation of a vast array of cytoprotective genes through the cognate antioxidant responsive element (ARE) [2]. Nrf2 plays a critical role in the basal activity and coordinated induction of genes encoding numerous antioxidant and phase II detoxifying enzymes, including superoxide dismutase (SOD), NAD(P)H:quinone oxidoreductase-1 (NQO1), heme oxygenase-1 (HO-1), glutamatecysteine ligase modulatory (GCLM) subunit, and thioredoxin, among others [3,4]. Therefore, targeting the coordinated upregulation of genes coding for detoxifying proteins, antioxidants, and anti-inflammatory regulators may be a potential therapeutic strategy to protect against insults such as inflammation and oxidative stress [5]. Indeed, targeting antioxidant defenses through modulation of Nrf2 could represent a new therapeutic approach with potentially major advances over conventional therapies [6].
In many clinical trials, 3-hydroxy-3-methyl-glutaryl-CoA reductase inhibitors (statins) have shown clear benefits in cardiovascular disease beyond their lipid-lowering actions [7], as they also function as antioxidants [8]. Statins not only decrease cellular reactive oxygen species production but also enhance the antioxidant response by upregulating the expression of many antioxidants [9]. Makabe et al. reported that fluvastatin protects vascular smooth muscle cells from oxidative stress through the Nrf2-dependent antioxidant pathway [10], although the molecular mechanism is unknown. The aim of this study was to identify the molecular mechanisms of Nrf2 activation by rosuvastatin (RSV).
Cell culture and transfection
Primary human umbilical vein endothelial cells (HUVECs; Cell System, Kirkland, WA, USA) were cultured in endothelial cell basal medium-2 (Lonza, Walkersville, MD, USA) containing 5% (v/v) fetal bovine serum under humidified conditions (95% air, 5% CO2) at 37 ºC. Subconfluent cells between passages 7 and 10 were used in the experiments. These cells were divided into four groups: 5.5 mM d-Glucose (Glu) + 22 mM l-Glu (normal glucose), normal glucose + 0.1, 1.0, or 10 μM RSV (RSV), 5.5 mM d-Glu + 22 mM d-Glu (high glucose), and high glucose + RSV. RSV was provided by AstraZeneca (London, UK). The dosage of RSV for the cells was in accordance to previous studies [11,12]. The cells were incubated at 37 ºC for 6 to 24 h (6 h for RNA isolation and nuclear protein preparation and 24 h for total protein preparation). Cos-7 cells (Health Protection Agency Culture Collections, Salisbury, UK) were used for the luciferase assays. Cells were cultured to 70% to 80% confluence and were transfected with 50 nmol/L p21Cip1-siRNA (sc-29427; Santa Cruz Biotechnology, Dallas, TX, USA) or control-siRNA for 24 h by using Lipofectamine 2000 (Life Technology, Grand Island, NY, USA) according to the manufacturer’s protocol. The extent of knockdown was assessed by western blotting.
Nrf2 reporter assay
Since transfection efficiency was low in HUVECs, we used monkey african green kidney fibroblast-like (Cos-7) cells (Health Protection Agency Culture Collections, Salisbry, UK) for the luciferase assay. Cos-7 cells were cultured with Dulbecco’s Modified Eagle’s Medium (Sigma-Aldrich Japan, Tokyo, Japan) containing 5% fetal bovine serum. Subconfluent cells were co-transfected with a reporter construct containing a human ARE-driven luciferase construct and a Renilla luciferase reporter as an internal control. After transfection, the cells were pretreated with RSV for 24 h. The cells were harvested, washed, homogenized, and analyzed for luciferase activity. The dual luciferase reporter assay system (Promega, Madison, WI, USA) and a luminometer (MimiLumat LB9506, Berthold Technology, Bad Wildbad, and Germany) were used to measure luciferase activity, according to the manufacturer's protocols.
mRNA stability assay
The mRNA stability of NQO1 and GCLM was determined by actinomycin D chase experiments. HUVECs were cultured under high glucose conditions for 24 h. At time 0, medium was exchanged with medium containing 1.0 μg/mL actinomycin D plus RSV or vehicle. RNA samples were prepared from RSV-treated and untreated cells in triplicate at 0, 1, 3, and 6 h. Quantitative real-time polymerase chain reaction (qPCR) of NQO1 and GCLM was performed as described below.
Cycloheximide chase assay for Nrf2 degradation/protein half-life analysis
After 24-h starvation, HUVECs were treated with 10 μM RSV for 6 h in a high glucose medium, and then harvested at 0, 15, 30, or 60 min after the addition of cycloheximide (25 μg/mL). Lysates from cycloheximide-treated cells were analyzed by western blotting.
RNA isolation and qPCR
Total RNA was extracted with TRIzol (Life Technology). Reverse transcriptase reactions were performed by using the ReverTra Ace qPCR RT Kit (Toyobo Biosciences, Osaka, Japan) for first-strand cDNA synthesis. qPCR was performed by using ABI Prism 7500 sequence detection system (Applied Biosystems, Foster City, CA, USA). Primers and probes for TaqMan analysis were designed by Primer Express 1.5 (Applied Biosystems) by using information from the supplier based on the sequence information from GenBank or EST databases. The primers and probes used for NQO1, GCLM, and p21Cip1 are next: NQO1 (NM_008706) Forward, 5′- ttctctggccgattcagagt -3′; Reverse, 5′- tccagacgtttcttccatcc -3′; Probe, FAM -5′- tttacagcattggccacactccacc -3′- TAMRA. GCLM (NM_008129) Forward, 5′- caatgacccgaaagaactgc -3′; Reverse, 5′- attcccctgctcttcacgat -3′; Probe, FAM-5′- attcccctgctcttcacgat -3′-TAMRA. p21Cip1 (NM_007669) Forward, 5′- ttgcactctggtgtctgagc -3′; Reverse, 5′- tctgcgcttggagtgataga -3′; Probe, FAM -5′ - aaacggaggcagaccagcctgac -3′ - TAMRA. For each gene, 10 ng of cDNA was analyzed on an ABI PRISM 7500 by using the TaqMan Universal PCR Master Mix (Applied Biosystems). A standard curve was prepared from a positive control with known concentrations, and the copy numbers of 18S and the target genes were calculated. The relative ratios of target gene to 18S are shown as a bar graph. Fold change analysis was based on standardizing RNA levels by correcting for 18S levels in the sample.
Western blot analysis
Extraction of total cellular proteins and nuclear proteins was performed by using an extraction reagent (T-PER tissue protein extraction reagent, NE-PER nuclear and cytoplasmic extraction kit; Thermo Fisher Scientific, Rockford, IL, USA), according to the manufacturer’s instructions. For immunoprecipitation assays, cells were lysed in radioimmunoprecipitation assay buffer (10 mM sodium phosphate, 150 mM NaCl, 1% Triton X-100, 1% sodium deoxycholate, and 0.1% sodium dodecyl sulfate SDS) in the presence of 1 mM dithiothreitol, 1 mM phenylmethanesulfonylfluoride, and a protease inhibitor cocktail (Roche Diagnostics, Indianapolis, IN, USA). For the analysis of Nrf2 expression, the proteasome inhibitor MG132 (10 μM; Sigma-Aldrich Japan) was added to the extraction buffer to avoid Nrf2 protein degradation. Cultured cells were homogenized on ice in 1.0 mL of lysis buffer containing protease inhibitors and centrifuged at 8000 g for 10 min. The protein concentration in the supernatants was determined with a Bio-Rad protein assay kit (Hercules, CA, USA). SDSpolyacrylamide gel electrophoresis was performed using cell lysates (20 μg/lane). Anti-Nrf2 (Cell Signaling Technology, Beverly, MA, USA), anti-p21Cip1 (Cell Signaling Technology), anti-Lamin A/C (Santa Cruz Biotechnology), anti-GAPDH (Santa Cruz Biotechnology), and antiactin (Sigma-Aldrich Japan) antibodies were used as the primary antibodies. For immunoprecipitation assays, cell lysates were incubated with anti-p21Cip1 antibody and Protein A/G PLUS-Agarose (Santa Cruz Biotechnology) for overnight at 4 ºC. After washing the protein A/G agarose beads complex, proteins were extracted with SDS buffer and proceed to western-blot for Nrf2 or p21Cip1. Signals were detected by using the ECL or ECL plus system (Amersham Biosciences, Piscataway, NJ, USA). The relative optical band densities were quantified by ImageJ analysis software V1.48 [13].
Statistical analyses
Data were expressed as mean ± standard error of mean (SEM). All variables were evaluated by two-tailed unpaired Student’s t-test or oneway analysis of variance for comparison of multiple means. A p value < 0.05 was considered significant.
RSV activates Nrf2
To determine whether RSV influences the ARE-Nrf2 pathway, we used an ARE-luciferase reporter assay in Cos-7 cells. ARE-luciferase activity was significantly higher in high glucose treated cells than in normal glucose conditions without RSV (Figure 1A). RSV increased ARE-luciferase activity under normal and high glucose conditions (Figure 1A). We then examined whether RSV increases nuclear translocation of Nrf2 protein in HUVECs. As shown in Figure 1B, rosuvastatin produced a marked increase in Nrf2 translocation into the nucleus under normal and high glucose conditions.
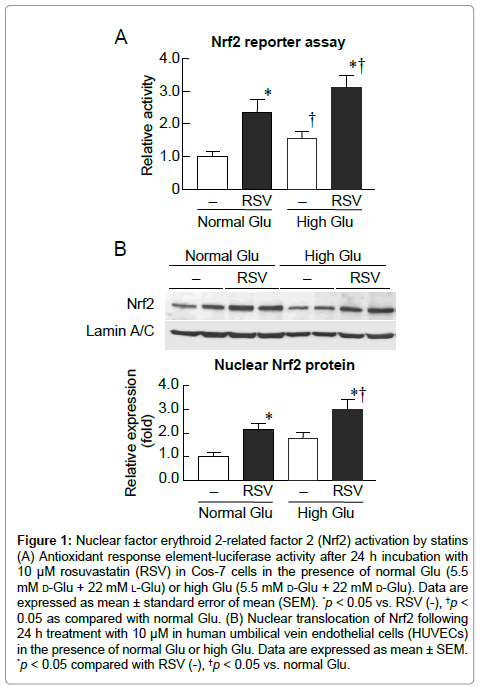
Figure 1: Nuclear factor erythroid 2-related factor 2 (Nrf2) activation by statins (A) Antioxidant response element-luciferase activity after 24 h incubation with 10 μM rosuvastatin (RSV) in Cos-7 cells in the presence of normal Glu (5.5 mM d-Glu + 22 mM l-Glu) or high Glu (5.5 mM d-Glu + 22 mM d-Glu). Data are expressed as mean ± standard error of mean (SEM). *p < 0.05 vs. RSV (-), †p< 0.05 as compared with normal Glu. (B) Nuclear translocation of Nrf2 following 24 h treatment with 10 μM in human umbilical vein endothelial cells (HUVECs) in the presence of normal Glu or high Glu. Data are expressed as mean ± SEM. *p < 0.05 compared with RSV (-), †p < 0.05 vs. normal Glu.
RSV increases Nrf2-targeted gene expression
Nrf2-targeted gene expression was evaluated by qPCR. RSV produced a dose-dependent increase in NQO1 and GCLM mRNA expression under normal and high glucose conditions (Figure 2A,2B). To examine the cause of this Nrf2-dependent induction, we assessed mRNA stability by actinomycin D chase experiments. RSV did not affect NQO1 and GCLM mRNA stability (Figure 2C, 2D).
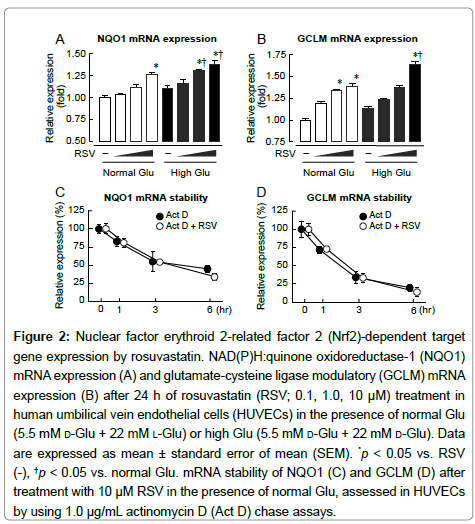
Figure 2: Nuclear factor erythroid 2-related factor 2 (Nrf2)-dependent target gene expression by rosuvastatin. NAD(P)H:quinone oxidoreductase-1 (NQO1) mRNA expression (A) and glutamate-cysteine ligase odulatory (GCLM) mRNA expression (B) after 24 h of rosuvastatin (RSV; 0.1, 1.0, 10 μM) treatment in human umbilical vein endothelial cells (HUVECs) in the presence of normal Glu (5.5 mM d-Glu + 22 mM l-Glu) or high Glu (5.5 mM d-Glu + 22 mM d-Glu). Data are expressed as mean ± standard error of mean (SEM). *p < 0.05 vs. RSV (-), †p < 0.05 vs. normal Glu. mRNA stability of NQO1 (C) and GCLM (D) after treatment with 10 μM RSV in the presence of normal Glu, assessed in HUVECs by using 1.0 μg/mL actinomycin D (Act D) chase assays.
RSV increases p21Cip1-Nrf2 protein complex formation
It has been reported that p21Cip1 activates Nrf2 by stabilizing Nrf2 protein [14]. To elucidate the mechanisms of Nrf2 activation by RSV, we examined p21Cip1 expression. As shown in Figure 3A, RSV treatment produced a dose-dependent increase in p21Cip1 mRNA under normal and high glucose conditions. p21Cip1 protein expression also increased in the presence of RSV (Figure 3B). The interaction between endogenous Nrf2 and p21Cip1 was assessed in RSV-treated HUVECs. As shown in Figure 3C, p21Cip1 protein formed a complex with Nrf2. Furthermore, binding of endogenous p21Cip1 to Nrf2 was enhanced by RSV under normal and high glucose conditions.
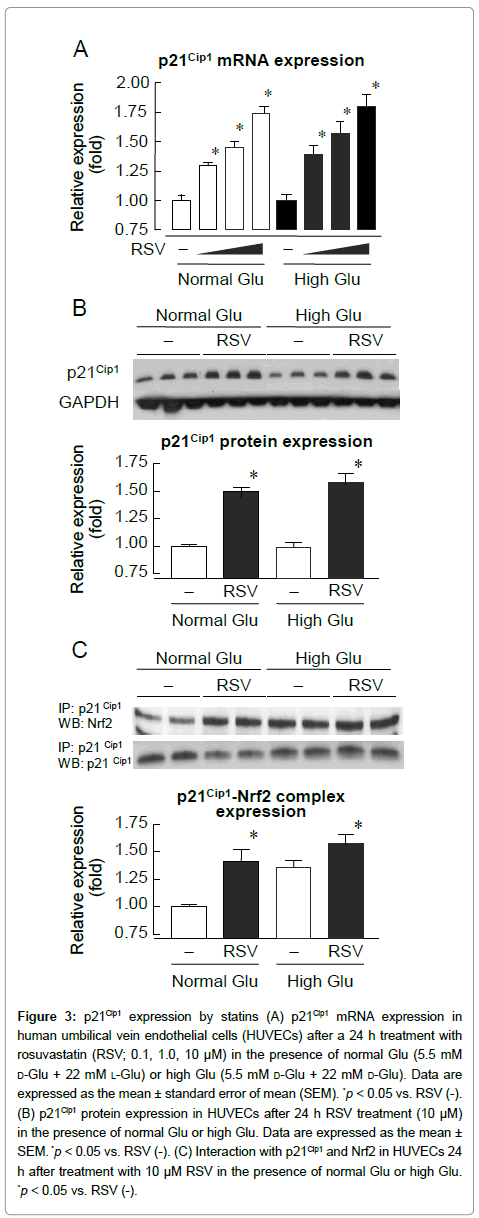
Figure 3: p21Cip1 expression by statins (A) p21Cip1 mRNA expression in human umbilical vein endothelial cells (HUVECs) after a 24 h treatment with rosuvastatin (RSV; 0.1, 1.0, 10 μM) in the presence of normal Glu (5.5 mM d-Glu + 22 mM l-Glu) or high Glu (5.5 mM d-Glu + 22 mM d-Glu). Data are expressed as the mean ± standard error of mean (SEM). *p < 0.05 vs. RSV (-). (B) p21Cip1 protein expression in HUVECs after 24 h RSV treatment (10 μM) in the presence of normal Glu or high Glu. Data are expressed as the mean± SEM. *p < 0.05 vs. RSV (-). (C) Interaction with p21Cip1 and Nrf2 in HUVECs 24 h after treatment with 10 μM RSV in the presence of normal Glu or high Glu. *p < 0.05 vs. RSV (-).
RSV increases Nrf2 protein stability
To examine the role of p21Cip1 in Nrf2 activation, we examined Nrf2 protein stability in p21Cip1 siRNA knockdown HUVECs. p21Cip1 protein expression was significantly reduced by transient transfection with p21Cip1siRNA versus control siRNA (Figure 4A). Nrf2 protein expression was increased by RSV in control cells but was not altered in p21Cip1 knockdown cells (Figure 4B). To assess the Nrf2 protein stability, a cycloheximide chase-based assay was conducted in p21Cip1 siRNA-transfected cells. RSV prolonged the Nrf2 half-life in control siRNA-transfected cells but did not affect the half-life of Nrf2 in p21Cip1 knockdown cells (Figure 4C).
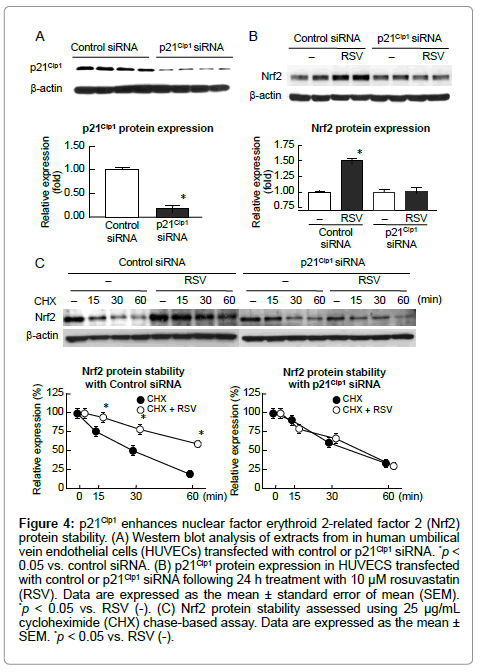
Figure 4: p21Cip1 enhances nuclear factor erythroid 2-related factor 2 (Nrf2) protein stability. (A) Western blot analysis of extracts from in human umbilical vein endothelial cells (HUVECs) transfected with control or p21Cip1 siRNA. *p< 0.05 vs. control siRNA. (B) p21Cip1 protein expression in HUVECS transfected with control or p21Cip1 siRNA following 24 h treatment with 10 μM rosuvastatin (RSV). Data are expressed as the mean ± standard error of mean (SEM). *p < 0.05 vs. RSV (-). (C) Nrf2 protein stability assessed using 25 μg/mL cycloheximide (CHX) chase-based assay. Data are expressed as the mean± SEM. *p < 0.05 vs. RSV (-).
The aim of this study was to identify the molecular mechanisms of Nrf2 activation by RSV. We demonstrated that RSV activates the transcription factor Nrf2 in endothelial cells, thereby enhancing Nrf2- dependent anti-oxidant genes. To our knowledge, these findings are the first reported evidence that RSV exerts its antioxidant effect through the Nrf2/ARE pathway in endothelial cells.
We have shown that p21Cip1 upregulation by RSV increased Nrf2 protein levels in HUVECs. Under non-stressed conditions, Nrf2 is negatively regulated by Keap1. Nrf2 is polyubiquitinated by the Keap1- Cul3 E3 ligase, and degraded by the 26S proteasome. Some electrophiles and oxidants oxidize the cysteine thiols of Keap1 and activate Nrf2 [15]. Recent reports have indicated that p21Cip1 directly upregulates Nrf2 protein levels [14]. These interactions have been mapped to the DLG and ETGE motifs in Nrf2 and the KRR motif in p21Cip1, which directly activates the Nrf2 pathway by competing with Keap1 for Nrf2 binding, thereby inhibiting Keap1-dependent ubiquitination of Nrf2 [14]. p21Cip1regulates various cellular processes such as cell-cycle arrest, DNA replication and repair, cell differentiation, senescence, and apoptosis. We showed that RSV upregulates p21Cip1 expression in HUVECs, prolonging Nrf2 protein stability and activating the Nrf2/ ARE pathway.
We confirmed that RSV activates the Nrf2/ARE pathway and enhances NQO1 and GCLM expression in endothelial cells. Some reports have suggested NQO1 activation is beneficial for the treatment of the metabolic syndrome by ameliorating obesity, preventing arterial smooth muscle cell proliferation, and mitigating spontaneous hypertension in animal models [16-18]. Other Nrf2-dependent antioxidants include heme oxygenase 1 (HO-1) and superoxide dismutase (SOD) [19]. Pharmacological inhibition of HO activity or deletion of the HO-1 gene worsens renal injury induced by toxic substances [20,21], ischemia reperfusion and diabetes [22]. SOD is the major antioxidant enzyme that removes superoxide, converting superoxide into hydrogen peroxide and molecular oxygen [23,24]. Thus, RSV may protect against cardiovascular diseases, including kidney diseases, through the upregulation of NQO1, HO-1, and SOD expression via Nrf2 activation.
RSV is a potent inhibitor of cholesterol biosynthesis and is used as a cholesterol-lowering drug [25]. Previous studies show that statins, including RSV, possess powerful pleiotropic effects that are independent of their effects on lipids and lipoproteins [26,27]. Recent reports have indicated that other statins also upregulate p21Cip1 expression [28,29], which mediates Nrf2 activation. Thus, we assume that other statins may also activate the Nrf2/ARE pathway, contributing to their pleiotropic effects, including decreasing oxidative stress and inflammation.
In conclusion, we showed that the anti-oxidant effects of RSV include the amplification of antioxidant potential through Nrf2 activation. Numerous reports have previously described the vascular protective mechanisms of statins; however, this is the first report to suggest that statins activate Nrf2 via p21Cip1 upregulation. Oxidative stress is the basis of morbidity in many cardiovascular diseases, including chronic kidney disease, and statin treatment may be a useful therapeutic strategy to enhance anti-oxidative capacity.
We would like to thank Ms. Etsuko Yorimasa, Ms. Makoto Moriya, Ms. Miyuki Yokohata, Ms. Keiko Satoh, and Ms. Satomi Hanada (Kawasaki Medical School) for animal care and technical assistance. This work was supported by Grants-in- Aid for Scientific Research from the Japan Society for the Promotion of Science (No. 23591209 to N.K., No. 24390218 to N.K.), and a Research Project Grant from Kawasaki Medical School (No. 25Ki-70 to C.I.).