Cell & Developmental Biology
Open Access
ISSN: 2168-9296
ISSN: 2168-9296
Research Article - (2013) Volume 2, Issue 4
A possible role of sonic hedgehog (Shh) in recruitment of C cell precursors to the Ultimobranchial Body (UB) and embryonic thyroid was investigated in Shh-/- mice. Nkx2.1 and Foxa2 co-expression distinguished UB originating in the fourth pharyngeal pouch from other derivatives of pharyngeal endoderm. In mutants UB formed a single structure that failed to bud and instead of fusing with the midline thyroid primordium adhered to the thymic rudiments. Mature C cells appeared in the UB remnant and ectopically in the thymic parenchyma, foregut endoderm and trachea. Thyroid did not contain C cells except minute numbers close to the tracheal interface. Tracing progeny in Shh-CRE/Rosa26R mice showed the vast majority of both UB and thyroid progenitors derived from Shh negative endoderm, but Shh expressing cells appeared in both thyroid primordia before fusion of the two. The findings indicate that Shh determines the endoderm territory for C cell differentiation and guides the migration of C cell precursors into the thyroid, presumably by regulating the separation of glandular domains in the pharyngeal pouch endoderm. A cell-autonomous role of Shh in thyroid morphogenesis is suggested.
<The thyroid contains two endocrine cell types, the follicular epithelial cells and the parafollicular C cells, which have different embryonic origins. Thyroid follicular progenitors are specified in the anterior foregut and form a midline primordium that buds from the pharyngeal endoderm and descends to the final position of the gland close to the larynx and proximal trachea. During migration the growing thyroid rudiment will fuse with the paired Ultimobranchial Bodies (UB) that arise from lateral endoderm forming the most posterior or distal pharyngeal pouch. The UB carry C cell precursors to the thyroid.
Based on the classical chick-quail chimeras experiments on chickquail chimeras by Le Dourain and co-workers [1,2] it is generally believed that C cells stem from cranial Neural Crest (NC). In fact, NC cells (NCC) constitute the main cell component of the avian ultimobranchial glands [3], which unlike the mammalian homologue exist as distinct organs that do not merge with away from the thyroid proper in birds, amphibians and reptiles. Mature C cells express neuropeptides and other neuronal markers [4]. Hence, medullary thyroid cancer originating from transformed C cells is classified as a neuroendocrine type of tumor [5]. A neuroectodermal origin of thyroid C cells was, however, recently challenged by data obtained from genetic fate mapping studies in mice [6]. This showed that neither the UB nor mature C cells expressed the R26 reporter gene that is known to be indelibly co-activated in all NCC progeny once by recombination with Wnt1 is expressed in the dorsal neural tube destined to a NC fate-Cre, which indelibly marks the progeny of NCC [7]. The embryonic origin of mammalian C cells thus remains to be established.
Genetic deletion of transcription factors involved in the development of the UB leads to aberrant localization or even loss of C cells [8-11]. However, the actual mechanisms and molecules involved regulating the entry of C-cell precursors to the UB and the forward dissemination of maturing C cells in the thyroid are yet unknown. Several lines of circumstantial evidence suggest that Shh, a prototypic morphogen with strong influence on both NCC and pharyngeal development, is an attractive candidate. Firstly, Shh promotes NCC migration and survival [12,13], reflected by the pronounced pattern disturbances appearing in Shh deficient craniofacial tissues to which NCC contribute significantly [14]. Secondly, Shh null mice exhibit multiple congenital malformations of many foregut derivatives comprising pharyngeal arch anomalies, esophageal atresia and lung dysplasia [15]. In the pharyngeal apparatus Shh signaling regulates Tbx1 transcriptional activity that is required for normal arch development [16]. We previously showed that the embryonic thyroid is hypoplastic and aberrantly located in the neck in both Shh -/- and Tbx1 -/- late gestation stage embryos [17,18], suggesting that a Shh/Tbx1- dependent morphogenetic signal also regulates thyroid organogenesis. Regarding the fate of the C cells it is interesting to note, however, that the pharyngeal phenotype of the Shh and Tbx1 homozygous mutants differ in several aspects. Specifically, loss of Tbx1 affects more pronouncedly posterior arch development leading to aplasia of the thymus, parathyroid glands and UB [9]. Accordingly, the thyroid of Tbx1 -/- embryos is lacking C cells [18]. In contrast, Shh deficiency impacts more on the formation of the anterior pharyngeal arches with no or little influence on the fourth pharyngeal pouch [19]. As the UB buds from the fourth pouch endoderm, the Shh mutant mouse may be instrumental deciphering the fate of the ultimobranchial epithelium in a situation where the contribution of NCC to pharyngeal development is abrogated. This in turn may provide clues to the identification of the C-cell source.
In this study, we investigated whether Shh regulates the formation of UB and thyroid homing of C-cell precursors in mice. We hypothesized that if the ancestral cells were NC derived mature C cells would not show up in the thyroid rudiment of Shh deficient embryos because they either do not survive or migrate to the correct location. Alternatively, if the C-cell progenitors are specified within the pouch endoderm normally lacking Shh expression [19] C cells would be expected to differentiate also in the absence of Shh signaling, similar to the follicular cell phenotype [17], and be present within the thyroid or UB parenchyma.
Animals
Breeding Shh +/- mice generated Shh homozygous mutant embryos. Shh-CRE mice with a CreGFP fusion under control of the Shh promoter [20] were crossed to ROSA26-Reporter (R26R) mice with a beta-galactosidase (β-gal) coding region (lacZ) controlled by the Cre-recombinase reporter [21] to generate Shh-CRE-/+; R26R-/+ embryos. The age of the embryos was estimated to 0.5 dpc the morning a vaginal plug was detected. The local ethic committee at the University of Gothenburg approved all animal handling and experiments.
Immunoreagents
The following antibodies were used for immunohistochemical staining: Rat mAb against E-cadherin (ECCD-2), rabbit pAb against Titf1/Nkx2.1 (Biopat, Milan, Italy), rabbit pAb against calcitonin (DAKO, Glostrup, Denmark), rabbit pAb against Foxa2 (Seven Hills Bioreagents, Cincinnati, OH), guinea pig pAb against Foxa1 (Seven Hills), rabbit pAb against β-gal (Molecular Probes Inc, Eugene, OR), and biotin-conjugated anti-rat and rhodamine red™-X-conjugated anti-rabbit IgGs (Jackson ImmunoResearch, West Grove, PA). Streptavidin-FITC was purchased from DAKO.
Immunohistochemistry
Embryos were fixated in 4% paraformaldehyde and incubated in 30% sucrose, both overnight at 4°C, followed by embedding in Tissue Tek (Sakura, Zoeterwoude, The Netherlands) and freezing at -80°C. To facilitate tissue infiltration of fixative embryos older that E12.5 were decapitated and removed from the caudal part of the trunk. Ten μm thick transverse or sagittal sections were cut on a Microm HM 500M cryostat microtome, collected on polylysine glass slides (Menzel-Gläser, Braunschweig, Germany), and air-dried. Sectioned embryonic tissues were permeabilized by incubation with 0.1% Triton X-100 for 20 min, blocked in PBS with 2% normal donkey serum (Jackson) for 1 hour at room temperature, and incubated overnight at 4°C with primary antibodies diluted in blocking buffer. Secondary antibodies diluted in blocking buffer were added for 1 hour followed by streptavidin-FITC for 30 minutes at room temperature. Each incubation step was followed by rinsing in 0.1% Triton X-100 for 3-5 minutes. Samples were viewed and photographed in a Nikon Microphot FXA epifluorescence microscope equipped with a QLC100 confocal laser scanning module (VisiTech International, Sunderland, UK) or, for analysis of GFP or β-gal immunoreactivity, a Bio Radiance 2000 Laser Scanning Microscope. Images were processed using the Image Pro Plus software (Media Cybernetics, Silver Spring, MD) and Adobe Photoshop Elements 3.0.
Impaired fusion of thyroid primordia and ectopic localization of C cells in Shh -/- mouse embryos
The morphogenetic traits of normal thyroid development in the mouse have been pinpointed in detail before [22]. To set the scene of the present study only some details of importance will be given here. After budding and descent the median thyroid primordium extends bilaterally in front of the prospective trachea and approaches the paired UB at E12.5 (Figure 1A) after which the fusion process starts. Lobe formation is completed two days later and endocrine progenitors start to differentiate into hormone-producing cells between E14.5-15.5 (Figure 1B). Thus, from E15 onwards thyroid C cells identified by their calcitonin content appear in increasing number scattered throughout the lobe parenchyma (Figure 1C).
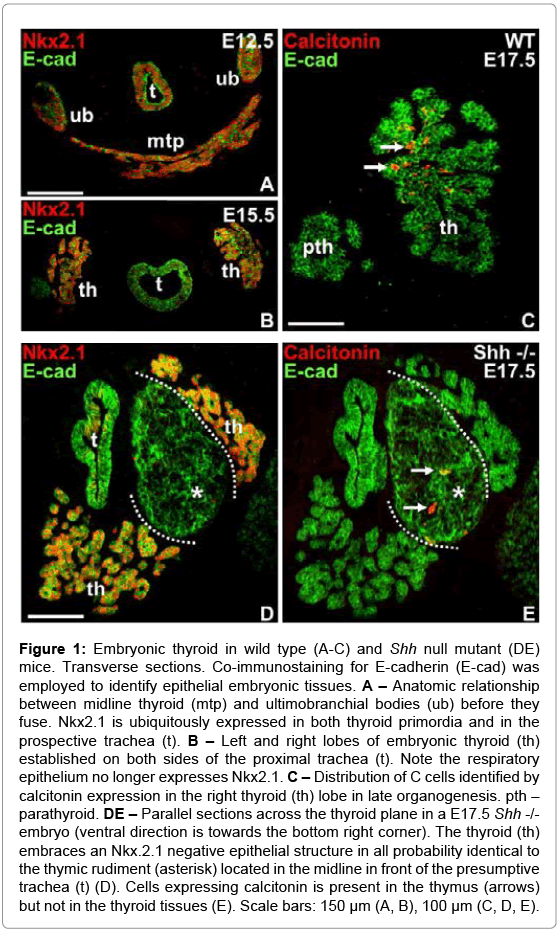
Figure 1: Embryonic thyroid in wild type (A-C) and Shh null mutant (DE) mice. Transverse sections. Co-immunostaining for E-cadherin (E-cad) was employed to identify epithelial embryonic tissues. A – Anatomic relationship between midline thyroid (mtp) and ultimobranchial bodies (ub) before they fuse. Nkx2.1 is ubiquitously expressed in both thyroid primordia and in the prospective trachea (t). B – Left and right lobes of embryonic thyroid (th) established on both sides of the proximal trachea (t). Note the respiratory epithelium no longer expresses Nkx2.1. C – Distribution of C cells identified by calcitonin expression in the right thyroid (th) lobe in late organogenesis. pth – parathyroid. DE – Parallel sections across the thyroid plane in a E17.5 Shh -/- embryo (ventral direction is towards the bottom right corner). The thyroid (th) embraces an Nkx.2.1 negative epithelial structure in all probability identical to the thymic rudiment (asterisk) located in the midline in front of the presumptive trachea (t) (D). Cells expressing calcitonin is present in the thymus (arrows) but not in the thyroid tissues (E). Scale bars: 150 μm (A, B), 100 μm (C, D, E).
The Shh knockout mouse is previously known to exhibit a thyroid phenotype consisting of a hypoplastic gland and ectopic thyroid tissue located in the tracheal wall [17]. The paired UB is reported to initially form normally from the fourth pharyngeal pouch [19], but there is no information on the developmental fate of the UB thereafter. We therefore screened E15-E19 Shh-/- embryos using E-cadherin as a marker of foregut endoderm to detect all pharyngeal derivatives and calcitonin to localize mature C cells. Because of the severely altered anatomy appearing progressively during mid-to-late development mutant embryos were serially sectioned from the level of the cyclopic eye to the distal lung buds to cover the entire pharyngeal region.
A consistent finding in Shh deficient late (E17.5) embryos was the presence of a large epithelial body located ventrally in close proximity to the endodermal tube that formed the tracheal rudiment (Figure 1D). This structure was partly embraced by but yet separated from thyroid tissue identified by Nkx2.1 expression and the presence of premature follicles (Figure 1D). Intriguingly, calcitonin-positive cells were encountered in the interior of the central tissue mass but not in the thyroid tissue proper (Figure 1E). No other solid E-cadherin+ structure was observed in the head-and-neck region of mutants, suggesting this might be a UB homologue that did not fuse with the midline thyroid and in which C cells reside because of a fusion defect. However, its further enlargement and the texture and organization of E-cadherin in its interior indicate this structure is identical to the thymic rudiment that in contrast to parathyroid is reported to develop essentially normally in the absence of Shh signaling [15]. In fact, serial sectioning revealed that thymus is the largest epithelial organ in the cervicothoracic region of E19 Shh null mutants, and that it is still attached in its central part to a hollow structure presumably constituting the pharyngeal epithelium (Figure 2C). Interestingly, single or small groups of cells expressing calcitonin were observed in different parts of the thymus (Figure 2A-D). In addition, the endoderm enclosing the pharyngeal cavity contained numerous C cells that apparently were part of the epithelial lining (Figure 2C and 2E). Inferior to the thymus-pharynx connection the endoderm of the proximal trachea displayed aberrant outgrowths of which many but not all cells were immunoreactive to calcitonin (Figure 2D). This location likely corresponds to where thyroid follicles ectopically bud from the tracheal wall in Shh mutants, as previously reported [17]. Notably, the thyroid proper present close to this tracheal segment did not show C cells in the parenchyma apart from a small number in the marginal zone towards the ectopic thyroid tissue that emerged from the trachea (Figure 2F).
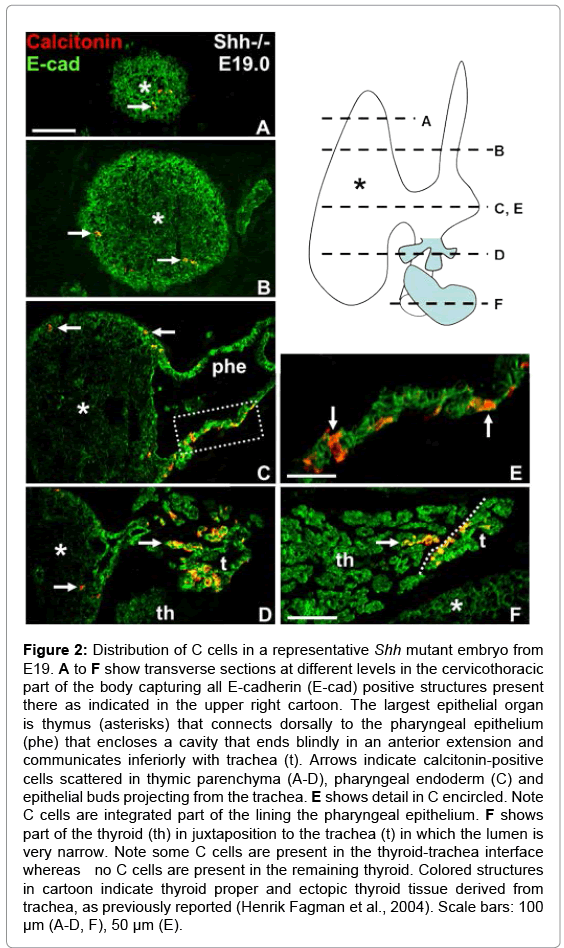
Figure 2: Distribution of C cells in a representative Shh mutant embryo from E19. A to F show transverse sections at different levels in the cervicothoracic part of the body capturing all E-cadherin (E-cad) positive structures present there as indicated in the upper right cartoon. The largest epithelial organ is thymus (asterisks) that connects dorsally to the pharyngeal epithelium (phe) that encloses a cavity that ends blindly in an anterior extension and communicates inferiorly with trachea (t). Arrows indicate calcitonin-positive cells scattered in thymic parenchyma (A-D), pharyngeal endoderm (C) and epithelial buds projecting from the trachea. E shows detail in C encircled. Note C cells are integrated part of the lining the pharyngeal epithelium. F shows part of the thyroid (th) in juxtaposition to the trachea (t) in which the lumen is very narrow. Note some C cells are present in the thyroid-trachea interface whereas no C cells are present in the remaining thyroid. Colored structures in cartoon indicate thyroid proper and ectopic thyroid tissue derived from trachea, as previously reported (Henrik Fagman et al., 2004). Scale bars: 100 μm (A-D, F), 50 μm (E).
Ultimobranchial bodies fail to bud from Shh deficient pharyngeal endoderm
C cell precursors normally enter the thyroid by fusion of the midline thyroid primordium and the UB as evidenced by retention of differentiated C cells in the UB remnant in Nkx2.1 haploinsufficient mice [8]. The ectopic distribution of C cells in thymus and trachea in Shh-/- embryos suggested C cell precursors aberrantly migrated there, presumably from the UB, or de novo developed in these locations. Notably, no solid or cystic structures reminiscent of UB could be found in late mutant embryos, indicating they were either not properly developed from the pharyngeal endoderm or complete regressed. We therefore investigated UB formation in mutant mice.
As originally shown [23], the UB normally express Nkx2.1 but this cannot be used as marker to distinguish UB-derived cells from thyroid follicular cells and their progenitors (Figure 1A and 1B). In fact, unlike for other budding organs originating in the pharyngeal endoderm there are yet no known transcription factors that specifically govern cell-autonomously the formation of the UB from the inferior-most pharyngeal pouch. In search for a suitable marker we focused on Foxa2 that is ubiquitously expressed in anterior endoderm [24] and recently found to be sustainably down-regulated in midline thyroid progenitors already when the thyroid bud is formed [25]. The expression pattern of Foxa2 was compared with that of Foxa1, both of which are known to exert either distinct or redundant functions in various endoderm-derived organs [26]. To our surprise this showed that the fourth pouch endoderm was entirely devoid of Foxa1 and Foxa2 immunoreactivity contrasting to the strong expression of both markers in adjacent pharyngeal endoderm (Figure 3A and 3B). However, as the UB emerged and delaminated from the endoderm lining the UB epithelium gradually acquired Foxa2 expression (Figure 3C) so that the majority of cells were Foxa2+ at E12.5 (Figure 3D). Importantly, at this stage the UB co-expressed Foxa2 and Nkx2.1 whereas the approaching midline thyroid expressed only Nkx2.1 (Figure 3E), indicating it is feasible to combine both markers to unerringly identify UB in early organogenesis. It is noteworthy that Foxa1 was not expressed in UB cells until E12.5 (Figure 3F) suggesting that Foxa1 and Foxa2 may exert distinct functions in UB development. Notably, after UB merged with the thyroid proper most UB-derived cells co-expressed Foxa1 and Foxa2 (data not shown).
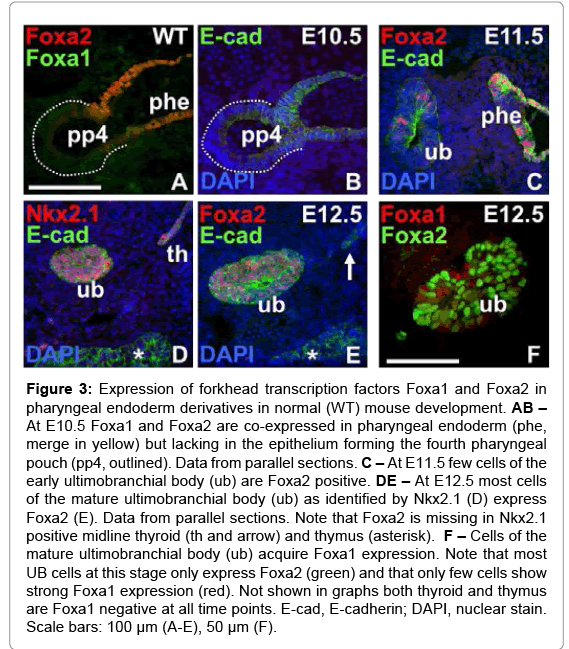
Figure 3: Expression of forkhead transcription factors Foxa1 and Foxa2 in pharyngeal endoderm derivatives in normal (WT) mouse development. AB – At E10.5 Foxa1 and Foxa2 are co-expressed in pharyngeal endoderm (phe, merge in yellow) but lacking in the epithelium forming the fourth pharyngeal pouch (pp4, outlined). Data from parallel sections. C – At E11.5 few cells of the early ultimobranchial body (ub) are Foxa2 positive. DE – At E12.5 most cells of the mature ultimobranchial body (ub) as identified by Nkx2.1 (D) express Foxa2 (E). Data from parallel sections. Note that Foxa2 is missing in Nkx2.1 positive midline thyroid (th and arrow) and thymus (asterisk). F – Cells of the mature ultimobranchial body (ub) acquire Foxa1 expression. Note that most UB cells at this stage only express Foxa2 (green) and that only few cells show strong Foxa1 expression (red). Not shown in graphs both thyroid and thymus are Foxa1 negative at all time points. E-cad, E-cadherin; DAPI, nuclear stain. Scale bars: 100 μm (A-E), 50 μm (F).
As previously reported [19], the initial patterning of pharyngeal arches and pouches is normal but the pouch identity is lost in Shh null mutants. Esophageal atresia further aggravates the anatomy of the pharyngotracheal junction in these mice [15]. We found that Foxa2 and Nkx2.1 were co-expressed bilaterally in a restricted zone of the endoderm located close to the pharyngeal opening of the prospective trachea (Figure 4A and 4B) at a time when the UB normally should exist as free bodies. The Foxa2+/Nkx2.1+ endoderm coalesced into a solid multi-layered structure that extended below the departure of the trachea thus forming the dorsal aspect of a blinded pocket to the pharyngeal cavity (Figure 4C and 4D). Further inferiorly, at the level of the thyroid rudiment, this assembly of cells was again divided into left and right domains of the endoderm (Figure 4E and 4F) before coexpression of Foxa2 and Nkx2.1 disappeared (Figure 4G and 4H). As there were no other parts of the pharyngeal endoderm in Shh deficient embryos that at this stage expressed Foxa2 and only a few Nkx2.1+ cells were observed elsewhere (Figure 4B; notably as shown in Figure 5 the only other tissue that expressed both markers was the distal trachea), this Foxa1+/Nkx2.1+ domain most likely constitutes the UB homologue that retained a bipartite shape reminiscent of the paired pouch origin and that failed to bud off. Unlike the mature wild-type UB, in which only few cells were Foxa1+ (Figure 3F), most if not all Foxa2+ cells present on both sides of the tracheal outlet and in the merged mid-piece co-expressed Foxa1 (Figure 4B and 4C, compare with AC), whereas the more distally located cells of the presumed UB homologue did not (Figure 4F and 4H, compare with EG). Similarly, the proximal trachea and associated pharyngeal endoderm were Foxa1+ whereas the distal portions of these tissues as well as the thyroid proper were entirely Foxa1 negative (Figure 4B, 4D, 4F and 4H).
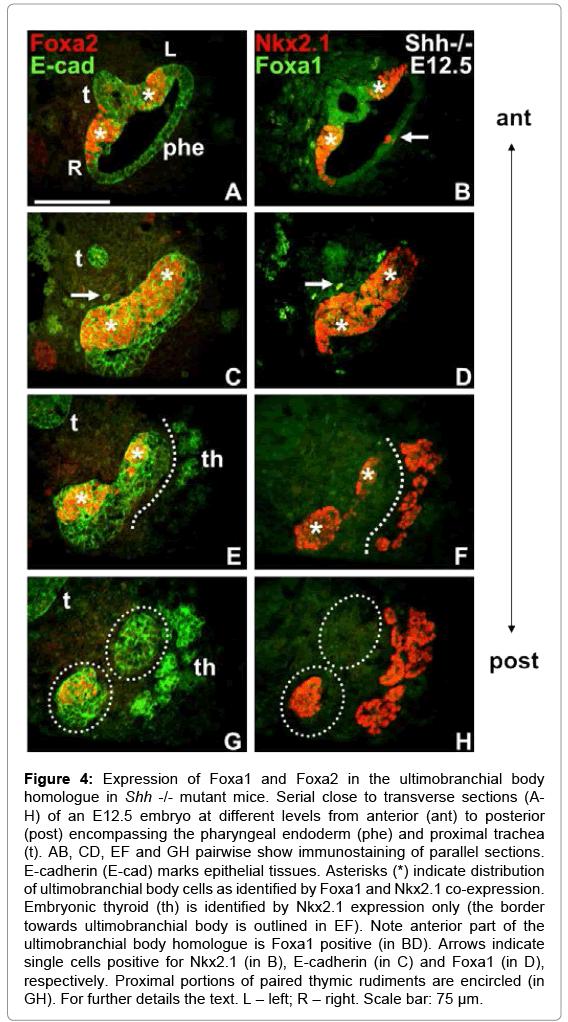
Figure 4: Expression of Foxa1 and Foxa2 in the ultimobranchial body homologue in Shh -/- mutant mice. Serial close to transverse sections (AH) of an E12.5 embryo at different levels from anterior (ant) to posterior (post) encompassing the pharyngeal endoderm (phe) and proximal trachea (t). AB, CD, EF and GH pairwise show immunostaining of parallel sections. E-cadherin (E-cad) marks epithelial tissues. Asterisks (*) indicate distribution of ultimobranchial body cells as identified by Foxa1 and Nkx2.1 co-expression. Embryonic thyroid (th) is identified by Nkx2.1 expression only (the border towards ultimobranchial body is outlined in EF). Note anterior part of the ultimobranchial body homologue is Foxa1 positive (in BD). Arrows indicate single cells positive for Nkx2.1 (in B), E-cadherin (in C) and Foxa1 (in D), respectively. Proximal portions of paired thymic rudiments are encircled (in GH). For further details the text. L – left; R – right. Scale bar: 75 μm.
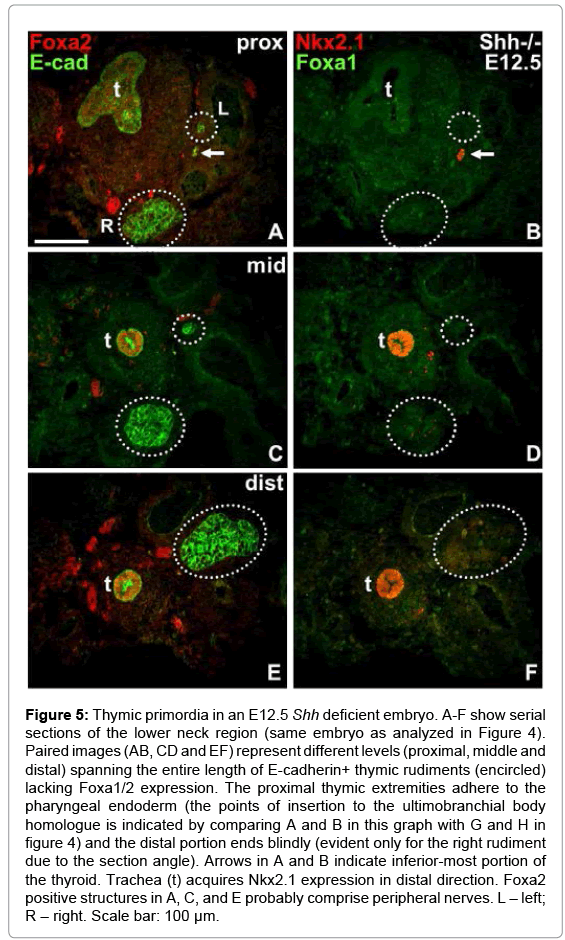
Figure 5: Thymic primordia in an E12.5 Shh deficient embryo. A-F show serial sections of the lower neck region (same embryo as analyzed in Figure 4). Paired images (AB, CD and EF) represent different levels (proximal, middle and distal) spanning the entire length of E-cadherin+ thymic rudiments (encircled) lacking Foxa1/2 expression. The proximal thymic extremities adhere to the pharyngeal endoderm (the points of insertion to the ultimobranchial body homologue is indicated by comparing A and B in this graph with G and H in figure 4) and the distal portion ends blindly (evident only for the right rudiment due to the section angle). Arrows in A and B indicate inferior-most portion of the thyroid. Trachea (t) acquires Nkx2.1 expression in distal direction. Foxa2 positive structures in A, C, and E probably comprise peripheral nerves. L – left; R – right. Scale bar: 100 μm.
Paired thymic primordia adhere to the ultimobranchial homologue in Shh knockouts
As shown in Figure 4E-H, the endodermal pocket harboring the UB homologue did not end by the disappearance of Foxa2+/Nkx2.1+ cells but continued as paired outgrowths located dorsally to the thyroid. Tracking their course in serial sections revealed they formed solid appendages with closed endings that extended into the thoracic cavity running in parallel to the trachea and cardiac outflow tract vessels (Figure 5A-F). The location further supports the notion that these structures indeed are identical to the thymic rudiments. Interestingly, although the proximal thymic stalks occasionally were very thin (see left-sided appendage in Figure 5A, 5C and 5E), they never lost contact with the pharyngeal wall and the UB remnant. In accordance with the original characterization of the Shh-/- phenotype [15], the expression of Foxa2 increased in the distal trachea as the respiratory epithelium also became Nkx2.1 positive (Figures 4 and 5).
Ultimobranchial and thymic progenitor cells form a compound organ that admit C cell precursors to invade the embryonic thymus in mice lacking Shh signaling
We re-examined E15.5 in Shh-/- embryos for localization of the UB remnant using the combination of Foxa2/Nkx2.1immunostaining as a reliable identification marker. This showed that UB cells remained positioned between the endoderm proper and the thymus that now formed a single solid structure (Figure 6A and 6B). Interestingly, cells co-expressing Foxa2 and Nkx2.1 invaded the thymic tissue beyond the interface zone (Figure 6A and 6B). In some section profiles it was obvious that thymus connected to pharynx via a convoluted extension of the endoderm that opened into a pouch-like cavity, which on one side communicated with the pharyngeal lumen and on the other side faced the base of the thymic body (Figure 6C). It is likely that this cavity emerged from the inferior pharyngeal pocket recognized earlier in development (Figure 4C and 4E). Notably, many cells in the lining epithelium expressed both Foxa2 and calcitonin (Figure 6C and 6D), suggesting both were of ultimobranchial origin and possibly identical. In thymus calcitonin positive cells assembled in the pharyngeal pole (Figure 6E) and along the proximal surface (Figure 6F), but was lacking in the more distal tissue portions (Figure 6G and 6H). Calcitonin immunoreactivity was never found in the embryonic thyroid facing thymus (Figure 6D-H).
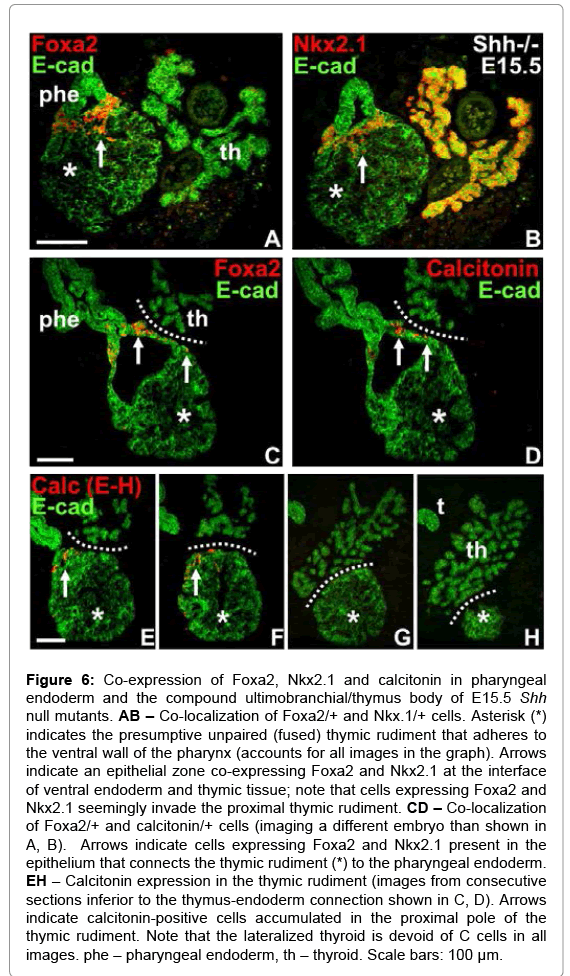
Figure 6: Co-expression of Foxa2, Nkx2.1 and calcitonin in pharyngeal endoderm and the compound ultimobranchial/thymus body of E15.5 Shh null mutants. AB – Co-localization of Foxa2/+ and Nkx.1/+ cells. Asterisk (*) indicates the presumptive unpaired (fused) thymic rudiment that adheres to the ventral wall of the pharynx (accounts for all images in the graph). Arrows indicate an epithelial zone co-expressing Foxa2 and Nkx2.1 at the interface of ventral endoderm and thymic tissue; note that cells expressing Foxa2 and Nkx2.1 seemingly invade the proximal thymic rudiment. CD – Co-localization of Foxa2/+ and calcitonin/+ cells (imaging a different embryo than shown in A, B). Arrows indicate cells expressing Foxa2 and Nkx2.1 present in the epithelium that connects the thymic rudiment (*) to the pharyngeal endoderm. EH – Calcitonin expression in the thymic rudiment (images from consecutive sections inferior to the thymus-endoderm connection shown in C, D). Arrows indicate calcitonin-positive cells accumulated in the proximal pole of the thymic rudiment. Note that the lateralized thyroid is devoid of C cells in all images. phe – pharyngeal endoderm, th – thyroid. Scale bars: 100 μm.
The erroneous development of third and fourth pharyngeal pouch derivatives in the Shh-/- mouse embryo is summarized in Figure 7. Failure of the UB to bud and fuse with the midline thyroid likely explains why C cell precursors do not colonize the thyroid parenchyma but instead migrate into the embryonic thymus and pharyngeal endoderm and differentiate into C cells in these ectopic locations. This is most likely made possible by the persistent attachment of the thymic and UB rudiments to each other and the pharyngeal endoderm. Ectopic C cells also appear within the Shh deficient tracheal epithelium. Together, the observations support previous notions that Shh plays a significant role determining pharyngeal pouch identity [19] and suggest that Shh restricts the endoderm domain in which the C cell lineage develops and mature.
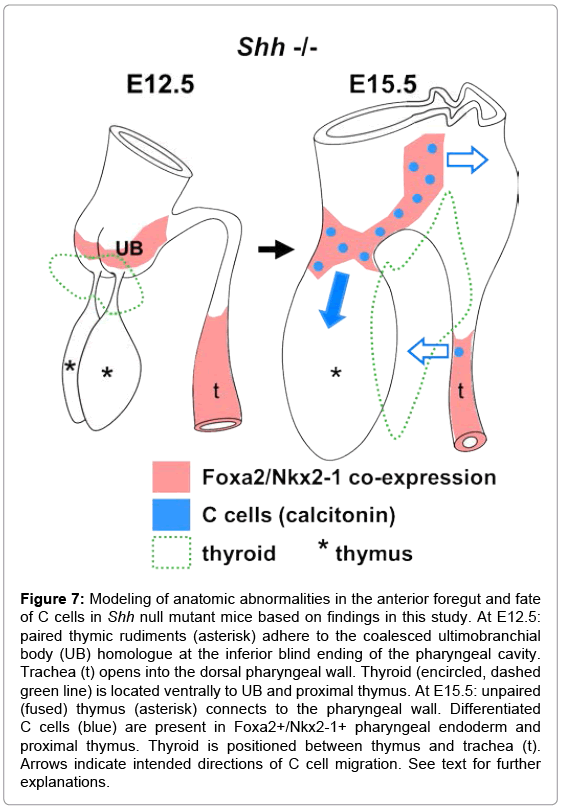
Figure 7: Modeling of anatomic abnormalities in the anterior foregut and fate of C cells in Shh null mutant mice based on findings in this study. At E12.5: paired thymic rudiments (asterisk) adhere to the coalesced ultimobranchial body (UB) homologue at the inferior blind ending of the pharyngeal cavity. Trachea (t) opens into the dorsal pharyngeal wall. Thyroid (encircled, dashed green line) is located ventrally to UB and proximal thymus. At E15.5: unpaired (fused) thymus (asterisk) connects to the pharyngeal wall. Differentiated C cells (blue) are present in Foxa2+/Nkx2-1+ pharyngeal endoderm and proximal thymus. Thyroid is positioned between thymus and trachea (t). Arrows indicate intended directions of C cell migration. See text for further explanations.
Genetic labeling identifies a subpopulation of Shh expressing endoderm-derived cells that might regulate thyroid development cell-autonomously
Although endoderm is the only source of Shh produced within the pharyngeal apparatus [27], Shh is known to be variably expressed along the anterior-posterior axis and missing in the fourth pharyngeal pouch at the primordial stage [19]. The restricted expression resembles that of other foregut budding organs e.g. pancreas and midline thyroid in which the respective placodes specifically lack Shh expression [28]. Together, this suggests that Shh regulates both thyroid and UB primordia by a non-cell autonomous mode of action. However, it cannot be ruled out that Shh is expressed in thyroid progenitor cells at later developmental stages as the embryonic thyroid and UB grow larger and merge. We addressed this by employing a genetic strategy to trace Shh expressing cells and their progeny during thyroid organogenesis. Shh-CRE mice with a CreGFP fusion under control of the Shh promoter [20] were crossed to ROSA26-Reporter (R26R) mice with a β-gal coding region (lacZ) controlled by the Cre recombinase [21] to generate Shh-CRE -/+; R26R -/+ mice. The GFP signal was sufficient to detect strong Shh expression as in notochord and lung bud but was too weak to stain less prominent tissues or scattered Shh+ cells with accuracy. Expression of the reporter gene was therefore detected by immunostaining of β-gal.
At E13.5 β-gal immunoreactivity was exclusively present in locations where Shh expression is expected from previous reports, e.g. floor plate of the neural tube (Figure 8A), notochord (Figure 8A and 8B), trachea and esophagus (Figure 8B). In early embryos focusing on anterior endoderm β-gal+ cells were ubiquitous in the ventral aspect of the foregut including the lung bud whereas most of not all cells of the dorsal endoderm were unlabeled (Figure 8C). A similar restricted expression of Shh in the anterior endoderm was previously reported [19]. The transgene did not disturb the expression of Nkx2-1 in the lung bud (Figure 8D). Together, this indicates that Shh-CRE/R26R recombination faithfully detects past and present Shh expression with a high cell and tissue specificity and without disturbing normal organogenesis in the foregut endoderm.
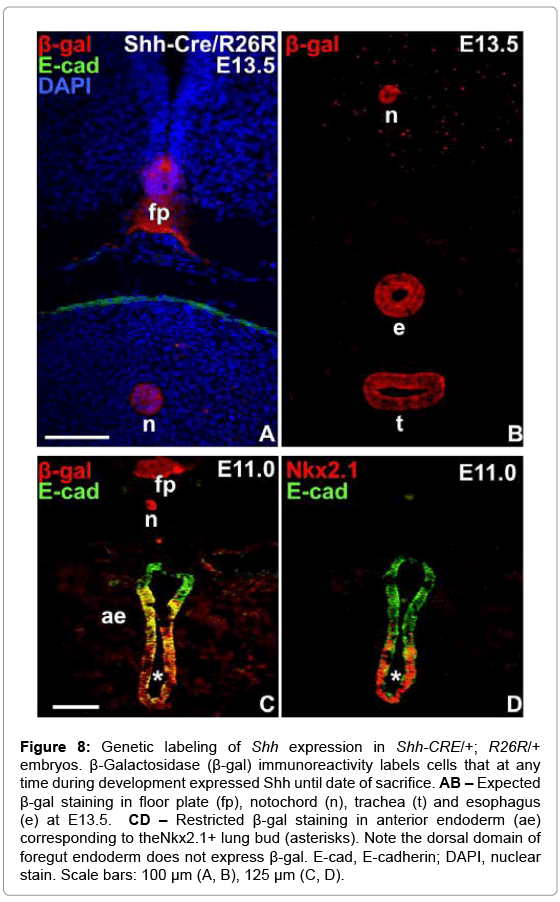
Figure 8: Genetic labeling of Shh expression in Shh-CRE/+; R26R/+ embryos. β-Galactosidase (β-gal) immunoreactivity labels cells that at any time during development expressed Shh until date of sacrifice. AB – Expected β-gal staining in floor plate (fp), notochord (n), trachea (t) and esophagus (e) at E13.5. CD – Restricted β-gal staining in anterior endoderm (ae) corresponding to theNkx2.1+ lung bud (asterisks). Note the dorsal domain of foregut endoderm does not express β-gal. E-cad, E-cadherin; DAPI, nuclear stain. Scale bars: 100 μm (A, B), 125 μm (C, D).
At E11.5 the endoderm of the fourth pharyngeal pouch from which UB develops did not express β-gal apart from a delimited segment of the ventral epithelium that was strongly labeled (Figure 9A). Similar to the domain from which lung budding takes place (Figure 8C) only the ventral portion of the endoderm proper was β-gal+ (Figure 9A). More surprisingly, the vast majority of progenitors forming the midline thyroid bud were negative for β-gal (Figure 9B and 9C), indicating they derived from an endoderm domain that also before the placode stage did not express Shh. As the midline thyroid and UB began to converge at E12.5 it was evident that both primordia were essentially free of β-gal+ cells except in the ventromedial extremity of the UB that contained a group of labeled cells (Figure 9D); their number corresponded well to those identified in the pouch endoderm at E11.5, suggesting they were identical. One day later, at the time of fusion of the two primordia, β-gal+ UB cells were mainly spread in the portion of the UB parenchyma that was in close contact with midline thyroid progenitors identified by their exclusive expression of Pax8 (Figure 9G and 9H). At this stage β-gal+ cells were also detected among Pax8 expressing progenitors located both close to the thyroid-UB interface and in more proximal portions of the midline primordium (Figure 8G and 8H). In all instances, the respiratory epithelium of the trachea was uniformly labeled by β-gal (Figure 9D and 9E), whereas cells in the esophagus gradually acquired reporter gene expression between E12.5-13.5 (Figure 9E and 9F). This indicated that the employed genetic strategy with confidence monitors not only the propagation of cells that expressed Shh earlier in development (e.g. lung bud) but also instantaneous transcription of Shh.
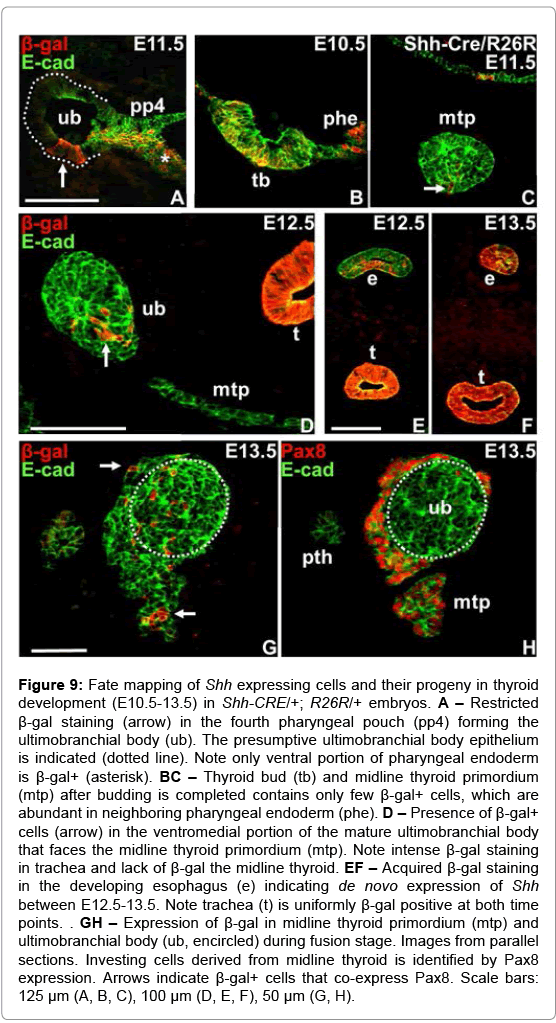
Figure 9: Fate mapping of Shh expressing cells and their progeny in thyroid development (E10.5-13.5) in Shh-CRE/+; R26R/+ embryos. A – Restricted β-gal staining (arrow) in the fourth pharyngeal pouch (pp4) forming the ultimobranchial body (ub). The presumptive ultimobranchial body epithelium is indicated (dotted line). Note only ventral portion of pharyngeal endoderm is β-gal+ (asterisk). BC – Thyroid bud (tb) and midline thyroid primordium (mtp) after budding is completed contains only few β-gal+ cells, which are abundant in neighboring pharyngeal endoderm (phe). D – Presence of β-gal+ cells (arrow) in the ventromedial portion of the mature ultimobranchial body that faces the midline thyroid primordium (mtp). Note intense β-gal staining in trachea and lack of β-gal the midline thyroid. EF – Acquired β-gal staining in the developing esophagus (e) indicating de novo expression of Shh between E12.5-13.5. Note trachea (t) is uniformly β-gal positive at both time points. . GH – Expression of β-gal in midline thyroid primordium (mtp) and ultimobranchial body (ub, encircled) during fusion stage. Images from parallel sections. Investing cells derived from midline thyroid is identified by Pax8 expression. Arrows indicate β-gal+ cells that co-express Pax8. Scale bars: 125 μm (A, B, C), 100 μm (D, E, F), 50 μm (G, H).
Thyroidal C cells and Shh progeny populating the thyroid gland are not identical
According to current understanding the midline thyroid primordium consists of a single progenitor lineage that give rise to the thyroid follicular cells, whereas the UB brings two precursor cell types that differentiate into, respectively, C cells and solid or cystic remnants of the ultimobranchial epithelium that can be found in the adult thyroid gland. However, which cells in the mature UB constitute the C cell precursors has not been elucidated. The prevailing concept of a NC origin of thyroid C cells was recently challenged in studies on Wnt1CRE/Rosa26R mice in which the reporter gene failed to encounter calcitonin-expressing cells [6]. We traced β-gal+ cells in Shh-CRE-/+; R26R-/+ embryos after the thyroid-UB fusion was completed to get an idea of whether UB cells expressing Shh might give rise to C cells. This showed that cells expressing β-gal+ were largely dispersed in the peripheral portions of the growing lobe parenchyma (Figure 10A) in which mature C cells did not yet appear (Figure 10B). Also later on when the UB remnant was largely disintegrated the thyroidal distribution of β-gal+ cells and calcitonin+ cells was distinctly different (Figure 10C and 10D). However, most cells forming small follicles were also devoid of β-gal (Figure 10C), suggesting Shh progeny might distinguish a new subpopulation of parenchymal cells in the differentiated mouse thyroid.
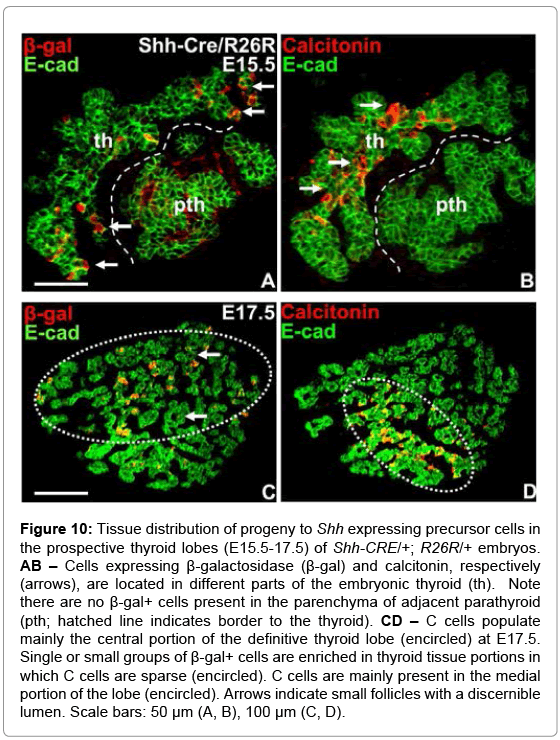
Figure 10: Tissue distribution of progeny to Shh expressing precursor cells in the prospective thyroid lobes (E15.5-17.5) of Shh-CRE/+; R26R/+ embryos. AB – Cells expressing β-galactosidase (β-gal) and calcitonin, respectively (arrows), are located in different parts of the embryonic thyroid (th). Note there are no β-gal+ cells present in the parenchyma of adjacent parathyroid (pth; hatched line indicates border to the thyroid). CD – C cells populate mainly the central portion of the definitive thyroid lobe (encircled) at E17.5. Single or small groups of β-gal+ cells are enriched in thyroid tissue portions in which C cells are sparse (encircled). C cells are mainly present in the medial portion of the lobe (encircled). Arrows indicate small follicles with a discernible lumen. Scale bars: 50 μm (A, B), 100 μm (C, D).
We found in Shh null mutant embryos that C cells are aberrantly distributed in extra-thyroid locations related to abnormal development of the pharyngeal apparatus. The onset at approximately E15 of calcitonin expression in mutants is similar to that of thyroid C cells in wild-type mice. Already at this stage C cells infiltrate the thymic rudiment that erroneously adheres to the pharyngeal endoderm. In late development (E19) calcitonin-expressing cells are more widely spread in the thymic tissue and the pharyngeal endoderm but fail to enter the thyroid apart from minute numbers derived from the adjacent trachea in which C cells ectopically emerge along with thyroid follicular cells, as previously reported [17]. Moreover, the paired UB fail to bud from the pharyngeal endoderm and a persistent connection between the UB epithelium and the thymic rudiment is evident. This likely provides the anatomical basis for misguided migration of C cell precursors in the absence of Shh signaling suggested [6]. Based on fate tracing of migrating NCC to the avian ultimobranchial gland in quail-chick chimeras and the presence of calcitonin immunoreactivity in these cells [1,2,29] it is envisaged that mammalian thyroid C cells originate from the NC. Accordingly, it is supposed that C cell precursors diverge from the stream of cranial NCC that is known to invade and pattern the pharyngeal arch mesenchyme [30], although this has not been possible to confirm due to current lack of specific markers. As Shh is required for both migration and survival of NCC [12,13,31] our findings that differentiated C cells populate pharyngeal derivatives in Shh-/- mice are remarkable and puzzling. In fact, NCC-mediated formation of in particular the posterior pharyngeal arches is strongly dependent on the action of Shh [32]. A clue to this may be provided by comparing the Shh phenotype with that of Tbx1 null mutants in which C cells are completely lacking [18]. Tbx1 is a T-box transcription factor that regulates pharyngeal arch development downstream of Shh by promoting NCC migration to the arches [16,33]. However, unlike Shh Tbx1 is expressed not only in pharyngeal endoderm but also in the subjacent mesoderm, and both are required for pharyngeal pouch formation and organogenesis [34]. This suggest that the more severe pharyngeal malformation encountered in the absence of Tbx1 [4] might be at least partly NCC-independent. Notably, early patterning and segmentation of the pharyngeal pouches do not require participation of NC [35], which probably explains why in the absence of Shh the thymus and UB anlagen yet are specified and committed to develop along the respective cell lineages. Together, this supports the idea of an alternative endoderm origin of thyroid C cells, as previously
We identified forkhead transcription factor Foxa2 as a novel UB epithelium marker. Intriguingly, Foxa2 was initially excluded from the pharyngeal pouch endoderm and gradually expressed in the maturing UB accompanying the expression of Nkx2.1, which previously is known to play a crucial role not only in embryonic thyroid and lung development but also for UB formation [8,36]. Unlike in UB (this study) and endoderm-derived respiratory epithelium [37] Foxa2 is not expressed in the thyroid primordium from the midline anlage [25], indicating that dual analysis of Foxa2 and Nkx2-1 faithfully distinguishes cell linages from the lateral and median thyroid anlagen. This enabled us to identify in Shh null mutants a compound UB homologue that persists as an integral part of the pharyngeal endoderm throughout morphogenesis and that Foxa2+/Nkx2-1+ cells derived from the UB remnant invaded the adhering thymic rudiment in which C cells were also encountered. Thyroid C cells are previously known to express Nkx2-1 [38-40] and preliminary findings in our laboratory indicate that mature C cells in addition co-express Foxa1 and Foxa2 (manuscript in preparation). Thus, although it cannot be ruled out that some NCC in Shh deficient mice yet may get a position close to the caudal pharyngeal wall and attain a differentiated phenotype typical of the UB epithelium these observations further strengthen the hypothesis that C cells might develop from the foregut endoderm. Notably, the presence of calcitonin-expressing cells within the simple epithelium of the pharyngeal cavity further indicates the epithelial nature of mouse C cells, as previously reported [4].
It is previously reported that Foxa2 participates in Shh-mediated activation of Tbx1 and that Foxa2 is down-regulated in pharyngeal endoderm in the absence of Shh [41]. This phenotype was essentially confirmed in the present study with the exception of the UB rudiment that continued to express Foxa2 in mice lacking Shh. Maintained expression of Foxa2 in this domain of the foregut thus probably explains why a rudimentary UB and later on C cells develop in Shh null mutants, presumably by promoting Tbx1 transcriptional activity, but not in Tbx1-deficient mice [9]. The normal UB acquired Foxa1 expression much later than Foxa2. In Shh null embryos the UB homolog showed a more restricted distribution of Foxa1 than Foxa2. Although we do not yet know the function these observations strongly suggest that Foxa1 and Foxa2 regulate distinct steps in UB development. A concerted action of Foxa1 and Foxa2 is established for several other organs originating from foregut endoderm [26].
The Shh-/- mutant UB retained as a placode-like structure in the dorsal aspect of the inferior pharyngeal endoderm long after it normally delaminates. The bipartite shape of the placode further suggested that bilateral assemblies of UB progenitors erroneously coalesce in the midline in the absence of Shh signaling. Moreover, the distal ending of the UB placode connected to the paired thymic rudiments thus forming a compound UB-thymus. These observations support previous notions that Shh is necessary to maintain the boundaries of the inferior pharyngeal pouches [19]. So far no factor(s) solely responsible for the formation the fourth pouch from with UB develops in mouse has been identified. However, loss of Shh is known to alter the expression of Fgf8, Pax1, and Bmp4 in the third pouch leading to confusion of dorsalventral patterning, which in turn triggers expansion of the endoderm domain committed to a thymic fate on the expense of the parathyroid anlage that fails to form [19]. In fact, the pharyngeal domain that expresses thymus-specific transcription factor Foxn1 is larger than solely accounted for by the third pouch, analogous to the expansion gut progenitors adopting a pancreatic fate when Shh signaling is inhibited [42]. It is possible that such enlargement of the thymic placode might infringe on patterning of the fourth pouch resulting in dorsalization and erroneous fusion of the UB rudiments as evident in the Shh mutant.
Lineage identity and propagation of progenitors in pharyngeal pouch endoderm are critically dependent on the balance of dorsalventral regionalization signals generated by morphogens. For example, expression of bone morphogenetic protein 4 (Bmp4) in the third pouch is restricted to the ventral thymic domain whereas Noggin, a natural Bmp4 antagonist, is exclusively present in the parathyroid primordium [43]. Accordingly, conditional activation of Noggin in mouse thymic progenitors leads to defective development of the thymus that becomes dysplastic and ectopically located in the neck [44]. As Bmp4 but not Noggin is also expressed in the fourth pharyngeal pouch [43], it is probable that the UB develop under the influence of a similar morphogenetic mechanism. This notion is supported by recent findings that targeted deletion of Bmp4 in pharyngeal endoderm and NC-derived arch mesenchyme leads to diminutive and dislocated UB that do not fuse with the embryonic thyroid accompanying the severe thymus and parathyroid malformations that predominate in these mutants [45]. Notably, Bmp4 is known to cooperate with or even mediate the action of endodermal Shh in gut development [46,47]. From this we propose that the UB phenotype in Shh deficient embryos may result from inactivation of a Shh -Bmp4 signaling pathway.
The typical action of Shh is non-cell-autonomous as in lung branching morphogenesis [48], but recent observations indicate that Shh may also stimulate cell behavior autonomously [49,50]. We investigated the conditions for such a possibility in thyroid development by tracing genetically Shh expressing pharyngeal endoderm and its progeny in Shh-CRE/R26R mice according to cell fate studies by others [20,51]. This showed, confirming previous expression data [17,19], that the dorsal portion of the anterior foregut does not express Shh. Moreover, the fourth pharyngeal pouch shares this feature apart from a restricted Shh+ segment present in the ventral pouch epithelium. This indicates that the emerging UB consists of at least two populations of endoderm progenitors; a predominant group that never expressed Shh and therefore likely descends from dorsal endoderm, and a smaller subset that either derives from Shh+ anterior endoderm or starts to express Shh expression once the pharyngeal pouch is formed. The fact that the delimited distribution of Shh+ and Shh– UB cells were largely maintained in the mature UB, suggests that the original topological orientation of Shh expressing cells as the UB emerged and delaminated from pharyngeal endoderm might provide directional cues for the subsequent fusion with the midline thyroid.
Genetic fate mapping further showed that progenitors committed to a thyroid follicular cell fate originate in Shh negative endoderm and that this property persists in the growing thyroid bud. It is previously known that the nascent thyroid bud increase in size without apparent cell proliferation indicating that additional cells most likely derive from pharyngeal endoderm outside the definitive placode [22]. This may be analogous to the pre-patterning of mouse liver progenitor cells in multiple distinct domains of the anterior endoderm before the liver bud is established [52]. In chick embryos patterning of the dorsal pancreatic anlage requires that Shh is actively repressed in the dorsal endoderm conducted by activin and Fgf2 secreted form the notochord [53]. It is likely although not formally proven that a similar mechanism regulates early thyroid development. Nonetheless, the fact that the thyroid bud emerges from a Shh negative placode surrounded by pharyngeal endoderm that ubiquitously express Shh suggests that further growth of the bud before it delaminates is manufactured by recruitment of progenitors from a more distant location, presumably by planar movement from the lateral or dorsal endoderm of single or small groups of cells, than previously appreciated.
It is previously known that progenitors of the follicular lineage differentiate normally in Shh deficient mice but the thyroid gland is severely hypoplastic [17]. As Shh is not expressed in the thyroid bud this suggests that Shh exerts a non-cell-autonomous effect possibly related to branchial arch vessel development in close proximity to the embryonic thyroid [17,54]. However, in the present study we observed signs of de novo Shh expression in a subset of Pax8+ progenitors present in the distal portions of the midline thyroid previous to fusion with the UB. In late thyroid development these cells did not mix with the UB remnant but were scattered throughout the prospective lobes accompanying the parenchymal growth cones that develop into follicles from E15 onwards [22]. These findings open for a possibility that Shh produced within the thyroid itself in addition might act as an intrinsic growth regulator.
In conclusion, we show here that Shh is required for correct routing of C cell precursors to the embryonic thyroid gland. In the absence of Shh signaling mature C cells instead appear in ectopic locations preferentially in the pharyngeal endoderm and developing thymus. The probable cause of the misguided migration of C cells is failure of the UB, which normally brings C cells to the thyroid, to bud off from the pharyngeal pouch and adherence of the retained UB homolog to the thymic rudiment. A possible cell-autonomous role of Shh in UBthyroid fusion is suggested from genetic labeling of Shh expression in the respective progenitor cell lineages.
This study was supported by grants from the Swedish Reseach Council, The Swedish Cancer Society, and Sahlgrenska University Hospital.