Cell & Developmental Biology
Open Access
ISSN: 2168-9296
ISSN: 2168-9296
Research Article - (2013) Volume 2, Issue 3
The field of regenerative medicine requires a reliable source of therapeutic cells in the context of muscular dystrophies. Cell therapies are among the treatment options being considered with the advances in stem cells and scaffolding domain. Umbilical cord tissue, Dental pulp and adipose tissues are abundant sources for clinical grade mesenchymal stem cells. These cells can differentiate into multiple lineages. The isolation, characterization, transdifferentiation, cell- gel formations and clinical applications of stroma derived mesenchymal progenitors are compared with each other source to evaluate the regenerative capability invitro. Dental pulp derived mesenchymal stem cells showed more propensity towards myogenic transdifferentiation although the abundance of the source and ease of higher cell yield was observed with cord and adipose tissue.
<Muscular dystrophy and mesenchymal progenitor cells
Muscular dystrophy [1] is a group of inherited disorders that involve muscle weakness and loss of muscle tissue, which get worse over time. Muscular dystrophies are passed down through families. All of the muscles may be affected or only specific groups of muscles may be affected typically as pathological symptoms in this disease pattern. The mainstay of treatment [2,3] includes steroids, physiotherapy, braces and wheelchairs, spinal surgery, and breathing aids. Very recently cell therapy was explored with muscle regeneration [4-8] as the qualitative measurement in coping with the progressive degeneration. Mesenchymal progenitor cells [9-11] are the latest addition to the battery of therapeutic cell types and are recognized by two important properties of extensive self renewal ability and multi-lineage differentiation potential into mesenchymal and non mesenchymal cell lineages. Adipose tissue [10,11], dental pulp [12,13] and umbilical cord tissue [14] derived mesenchymal progenitor cells originate from different anatomic sites while the stemness of the cells depends on the protocols of isolation as well as culture conditions. To obtain each of these clinical sources for proposed therapeutic application, the method to source, harvest the cells and multiply those for specific lineage regeneration invitro/vivo are dependent on appropriateness of the cell type proposed. Several groups have worked on each of these sources and published both in-vitro [15,16] and clinical utility results that address muscular degeneration independently. Combinational treatment approaches combining tissue engineering and stem cells infusion have yielded better procedures in treating Muscular dystrophies [15].
In the current study, we evaluated and compared the multilineage in vitro transdifferentiation capabilities of adipose, dental pulp and cord stroma derived mesenchymal progenitor cells with respect to induced myogenesis along with their cell kinetics and biocompatibility with Polyethylene Glycol – Fibrinogen biomaterial (PEG-FB). The objective of this study was to compare and weigh three different clinical sources and their myogenic transdifferentiation potentials.
Sourcing tissue and cell harvesting
Human subcutaneous adipose tissue (n=3) sample sources were obtained following routine, lipoaspiration. All human biological samples were handled with Institutional Ethics Committee approval and with informed consents of the donors for research. The lipoaspirate (chunks of adipose tissue) was extensively washed with Phosphate Buffer Saline (PBS) by centrifuging at 3500 rpm and digested in trypsin 0.25% solution in 5% CO2 incubator for 1 hour at 37°C. After the incubation, the sample was centrifuged at 3500 rpm for 5 minutes to separate the stromal cell fraction (pellet) from adiposities. The supernatant was discarded, and the cell pellet was suspended in growth medium (DMEM+ 5% Fetal Bovine Serum (FBS), supplemented with growth factors - Vascular Endothelial Growth Factor (VEGF), Epidermal Growth Factor (EGF)). The plastic adherent fibroblastic cells were cultured for three more passages to expand. Cells at passage 4 were detached from the flask by the treatment with trypsin/EDTA for invitro transdifferentiation.
The exfoliated teeth (n=3) collected in phosphate buffer saline [13,17] and antibiotic solution were cut with a sterile classic max hard wire cutter (1.5 mm). Pulp harvested was placed in a sterile petri dish. The tissue was then digested with trypsin 0.25% for 45 minutes at 37ºC with 5% CO2 at 37°C. The neutralized pulp in complete growth medium was cultured for 4 weeks. The adherent cells were passaged thrice and used for invitro transdifferentiation procedure.
Donated Umbilical Cord (UC) tissue (n=3) collected with informed consent from elective deliveries in sterile tube containing phosphate buffered saline with antibiotics. UC was then cut longitudinally, and around 2-3 ml of 0.25% trypsin was added and incubated at 37°C in a humidified chamber with 5% CO2 for 30 mins [18]. After 30 min of incubation, the UC was then neutralized and dissected into small pieces approximately 0.5 cm3 in length. The resulting dissected UC lining was placed into fresh DPBS for explanting. One dissected cord lining piece was cut into 3–4 strips with the gelatinous side up. Three to four explants of UC were seeded onto coated 60 mm petri dish and 1.5 ml of media was added into the plate. Explants and cells were grown in an amalgam of Dulbecco’s Modified Eagle’s medium, 10% fetal bovine serum (GIBCO), pen-strep, 5 μg/ml of EGF and 5 μg/ml of VEGF. The plates were then incubated at 37°C with 5% CO2, with alternative day change of fresh medium. At 70-80% confluence, cells were detached and passaged thrice and plated for in vitro transdifferentiation experiments.
Cellular Growth Kinetics
Population doubling time: For population doubling times [19] at various cell densities, we seeded cells harvested from adipose, dental pulp and cord tissue (P2 to P5) in a 35 mm plate at 1×104, 5×104 or 1×105 cells/plate, and incubated them at 37°C in a humidified atmosphere containing 5% CO2. At confluence they were detached by 0.05% Trypsin-EDTA treatment, counted and reseeded at the initial cell density. This procedure was repeated at every passage for 24 days. The population doubling times were calculated according to the formula PDT=(T-T0) lg2/(lgNt-lgN0), where PDT is the population doubling time, T0 is the starting time of cell culture and T is the ending time of cell culture, while N0 and Nt separately are the cell numbers at the start and the end of each culture, respectively.
Cell viability: Cell viability and cell number was determined using Tali Viability Dead cell Red Kit (Manufacturer: Life Technologies, Invitrogen) with Tali Image-Based by following the instructions in manufacturer’s manual.
Cell surface marking: Immunophenotyping of cells at P3 was carried out using Tali image based cytometry. The cells were dissociated and re-suspended in saline at 1×106 cells/ml concentration. From that sample, 100 μl of cell suspension with added primary anti body at the ratio 1: 20 was analyzed for the following surface markers CD 34, CD45, CD73, CD90, CD105, HLA ABC & HLA DR.
Induction of transdifferentiation in vitro: Adipogenic differentiation (induction) has been performed in monolayer culture after cells became 80% confluent. The P4 cells were cultured in adipogenic differentiation medium (Invitrogen) and maintained in the induction medium for 21 days. To detect fat droplets and quantify adipogenesis cells were stained with Oil-O-red [20,21].
Osteogenic differentiation (induction) has been performed in monolayer culture at 80% cell confluency. The P4 cells were cultured in osteogenic differentiation medium for 21 days. To determine osteogenesis, calcium depositions were detected in induced cells by Von Kossa staining method [20,21].
The harvested cells from the three different sources were reseeded in P4 stage and supplemented with 0.1 μm dexamethazone, 50 μm hydrocortisol and 5% horse serum in Growth medium. Cells were maintained in the induction medium for 45 days [20]. A different stage of myogenesis was quantified by microscopic assessment of stained cells by myoD [22] and dystrophin markers [22] per focus at 20X magnification at day 20 and 45 respectively of the culture period, while morphologically different satellite like cells during the myogenesis induced invitro were quantified by visual average counting from 5 different foci for percentage calculation.
Measurement of cell-gel stability
Cell-Gel encapsulation: Research grade PEG-Fibrinogen (Irgacure 2959, Technion, Israel) was polymerized as per the supplier’s instructions. Briefly, cells were added at the desired density at a final concentration of 7 and 8.3 mg fibrinogen in 1 ml C solution and exposed immediately to UV light at a wavelength of 365 nm, Intensity - 7 mW/cm2 (Vilber Lourmat) for 7 minutes at room temperature as per the supplier manual.
Duration of contact Vs viability of encapsulated cells: 2-3 million cells/ml of the precursor solution with 7, 8.3 mg fibrinogen was dispensed in molds (n=12) greased to the bottom of 60 mm Culture dish. For polymerization the PEG-FB solution with cells was exposed to long-wave laser light (365 nm). The plugged gels (n=3) were degraded in collagenase (type 2) solution (1mg/ml) immediately after polymerization, after 30 min, 1 hr, 2 hr and 3 hr time lapse and evaluated for the viability of cells expressed in the gels by countess method.
Statistical analyses: All data is expressed as means or representative of at least three separate experiments.
Cultured and passaged mesenchymal cells maintained characteristic phenotypic integrity
Adipose tissue, Dental pulp and Umbilical cord tissue samples collected from the donors yielded mesenchymal like fibroblastic cells that could be expanded up to P7. They were spindle shaped with thin and long morphology, adhering to coated tissue culture grade plastic surfaces (Figure 1). The average cell sizes were 10-17 μm (adiposederived), 14-17 μm (dental pulp-derived) and 8-12 μm (cord-derived) respectively at P5. Cells at P4 were used for transdifferentiation assays that showed >90% viability (Figure 2). The calculated population doubling time for both cord and dental pulp derived cells was 48 hr for the first 6 passages and increased to 76hr thereafter, whereas for adipose derived cells the population doubling time was 40.5 hr for the first 5 passages of the culture growth.

Figure 1: Morphology of ADMSCs, adhering to coated tissue culture grade plastic surfaces.

Figure 2: Indicates viability of ADMSCs tested by Propidium iodide method showing 96% by Tali Image based cytometer.
Mesenchymal cells expressed stem cell specific quantitative phenotype
Upon immune-phenotyping of mesenchymal cells, we checked and verified the absence of cell surface antigens HLA DR, CD34, CD45. The same cells from three different sources were uniformly positive for HLA ABC, CD90, CD105, CD73 while the percentage expression by each cell type was differently quantified (Figure 3).
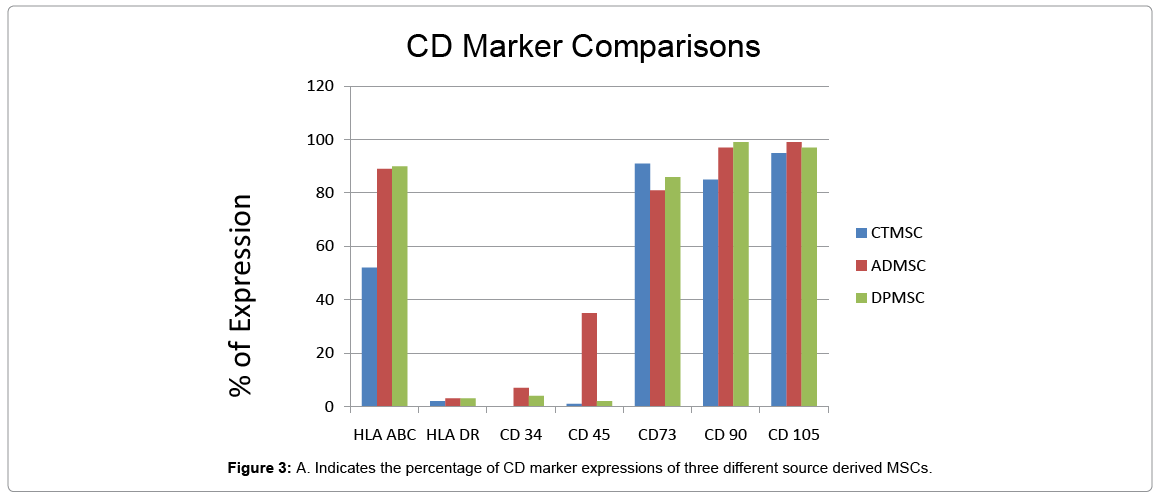
Figure 3: A. Indicates the percentage of CD marker expressions of three different source derived MSCs.
Transdifferentiation capabilities reflect lineage propensity
Adipose, Dental pulp and Umbilical cord tissue derived stromal mesenchymal progenitors differentiated into osteoblasts, adipocytes, chondrocytes (data not shown) and myocytes upon induction. Significant calcium depositions were detected in the induced cells confirmed by Von Kossa staining (Figure 4A1, Figure 5B1 and Figure 6C1). In the adipocyte differentiation medium, the cells showed accumulation of lipid rich vacuoles stained with Oil-O-Red, a fat soluble dye by 28 days (Figure 4A2, Figure 5B2 and Figure 6C2).
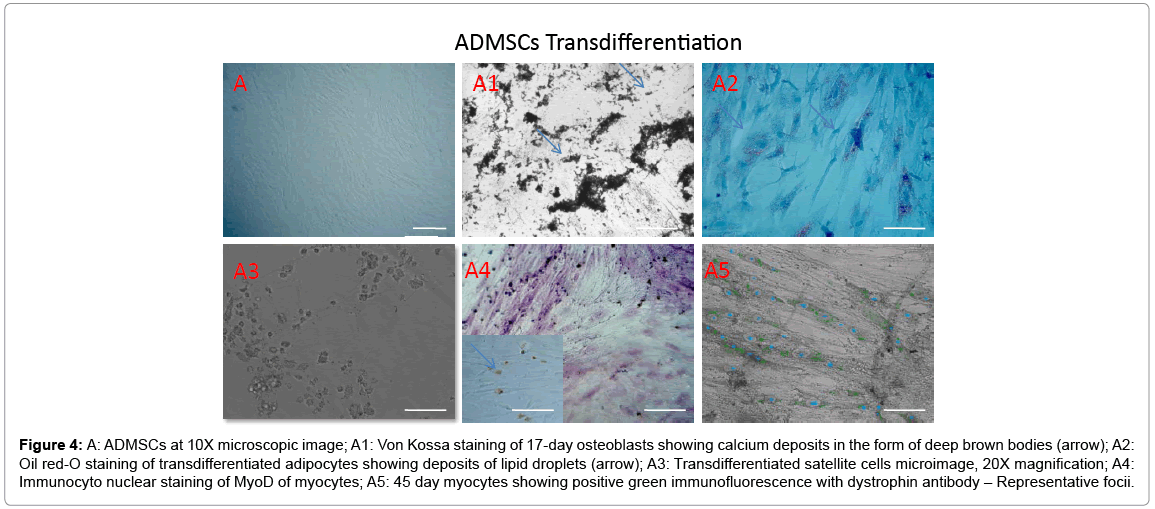
Figure 4: A: ADMSCs at 10X microscopic image; A1: Von Kossa staining of 17-day osteoblasts showing calcium deposits in the form of deep brown bodies (arrow); A2: Oil red-O staining of transdifferentiated adipocytes showing deposits of lipid droplets (arrow); A3: Transdifferentiated satellite cells microimage, 20X magnification; A4: Immunocyto nuclear staining of MyoD of myocytes; A5: 45 day myocytes showing positive green immunofluorescence with dystrophin antibody – Representative focii.
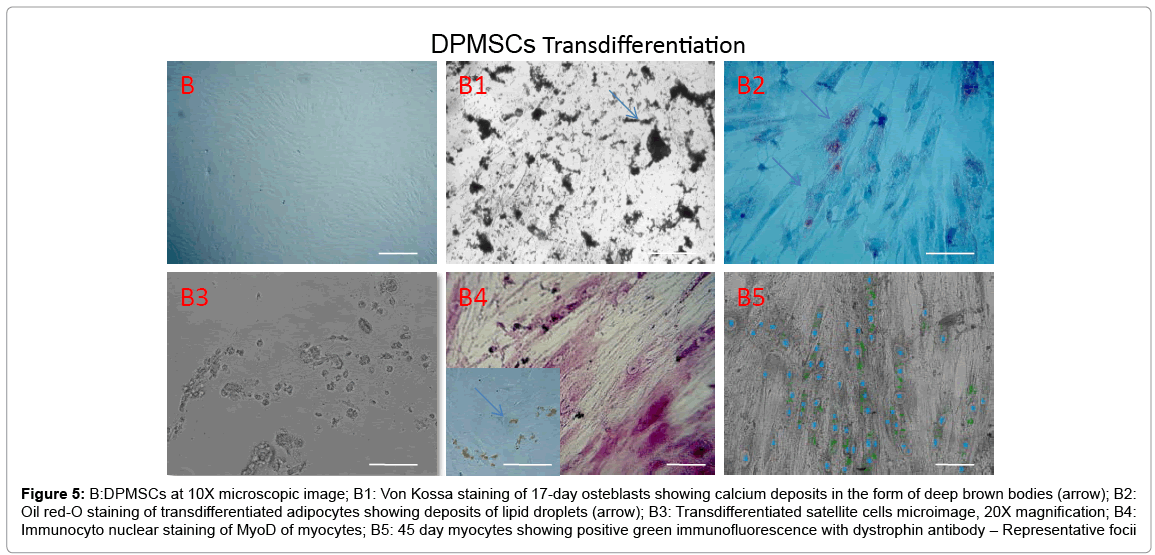
Figure 5: B:DPMSCs at 10X microscopic image; B1: Von Kossa staining of 17-day osteblasts showing calcium deposits in the form of deep brown bodies (arrow); B2: Oil red-O staining of transdifferentiated adipocytes showing deposits of lipid droplets (arrow); B3: Transdifferentiated satellite cells microimage, 20X magnification; B4: Immunocyto nuclear staining of MyoD of myocytes; B5: 45 day myocytes showing positive green immunofluorescence with dystrophin antibody – Representative focii
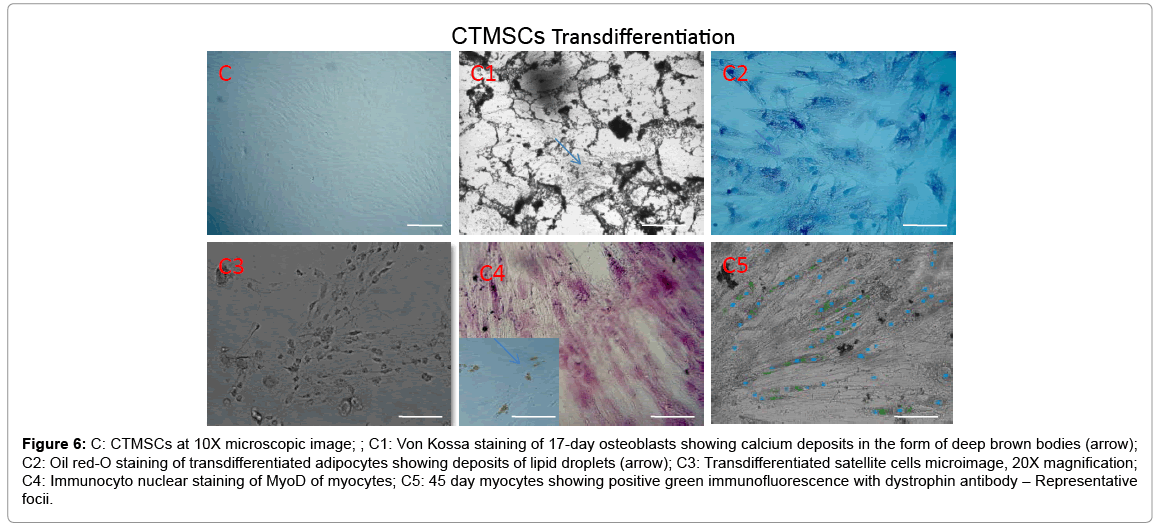
Figure 6: C: CTMSCs at 10X microscopic image; ; C1: Von Kossa staining of 17-day osteoblasts showing calcium deposits in the form of deep brown bodies (arrow); C2: Oil red-O staining of transdifferentiated adipocytes showing deposits of lipid droplets (arrow); C3: Transdifferentiated satellite cells microimage, 20X magnification; C4: Immunocyto nuclear staining of MyoD of myocytes; C5: 45 day myocytes showing positive green immunofluorescence with dystrophin antibody – Representative focii.
Stromal mesenchymal progenitors derived from all the three sources showed a greater tendency of transdifferentiation towards myogenesis with observed changes in the phenotype during induction period (Figures 4-6). Adipose MSCs expressed an average of 58.3% of satellite cells (microscopic enumeration) at the end of 7 days during transdifferentiation process, 67.6% of MyoD positive myocytes while 17% of myotubes with scattered dystrophin expression in cytoplasmic zones. Dental and Cord tissue MSCs expressed 43%, 45% of satellite cells respectively, 59%, 62% respectively of MyoD positive myocytes while 22% and 19% respectively of myotubular structures formation (Table 1). Uninduced MSCs served as a negative control.
| Source | Satellite cells (%) foci = 5 | MyoD positive cells (%) foci = 5 | Dystrophin positive cells (%) foci = 5 |
|---|---|---|---|
| ADMSCs | 58.3 | 67.6 | 17 |
| DPMSCs | 43 | 59 | 22 |
| CTMSCs | 45 | 62 | 19 |
Table 1: Transdifferentiated positive cell profile.
Stable gel-cell ratio supports viability and uniform distribution
7 mg/ml of PEGF-FB with 3 million cells retained viability even after 3 hr of polymerization with U.V exposure for 7 min. There was no significant affect of either the U.V exposure or the time of observation after polymerization at room temperature on the viability of cells evaluated (Table 2).
| Time (post polymerization) | Average % of (2-3 million adipose derived cells) Viability (n=3) | % of (2-3 million cord derived cells) Viability (n=3) | % of (2-3 million dental pulp derived cells) Viability (n=3) |
| 0 Hours | 91 | 92 | 91 |
| 30 Min | 88 | 87 | 89 |
| 01 Hour | 85 | 84 | 80 |
| 02 Hour | 80 | 83 | 80 |
| 03 Hour | 80 | 80 | 80 |
Table 2: Time Vs Viability of cells in PEGF-FB.
For decades, basic research on MSCs and their proliferation and differentiation potential as well as their immunosuppressive properties were repeatedly validated by stand-alone MSCs or by MSCs embedded in biocompatible hydrogels entering clinical applications in regenerative medicine [23]. Several clinical sources of MSCs have been identified and thoroughly characterized by comparing the expressions of function specific markers correlating with their clinical applicability [14,24]. In this context, our study investigated the propensity of MSCs derived from Adipose, Dental pulp and Cord tissue sources towards the myogenic transdifferentiation specifically to compare the appropriateness to address muscular degeneration pathologies along with their biocompatibility in the PEGF hydrogel.
Once primary cell lines from Adipose, Cord tissue and Dental pulp were established, the different MSC populations were characterized following the criteria suggested by the International Society for Cellular Therapy (ISCT). Cytometric principle of Tali, a suspension cell based imaging system was used to assess and evaluate the percentage of cultured cells that do not belong to the hematopoietic lineage (CD45−) CD34, HLA DR, but co express MSC antigens CD105, CD90, and CD73 (Figure 3) along with HLA ABC. Our results substantiate that the cells derived from the above mentioned three different sources handled at P3 were indeed mesenchymal progenitor in nature as per the ISCT criteria with typical surface profile though the quantity wise percentage expression have never been attributed to any specific cell source but with the culture expansion protocols as per the work done by Hagmann et al (2013). De Luca et al. (2013) compared some biological properties of mesenchymal cells isolated from subcutaneous and visceral adipose tissues with the “gold standard” BM-derived MSCs and evidenced the peculiarities that distinguish and characterize MSCs, while in our study, we indicated the characteristics distinguishing surface marker expression on all the primary cell lines harvested and stabilized for transdifferentiation procedures in vitro. Parallely harvested and cultured under suitable conditions, the growth kinetics of mesenchymal stem cells derived from dental pulp and cord tissue as in population doubling was analyzed to be the same while it differed as 40.5 hr for adipose source. The initial harvest period was although long as 22-25 days for dental pulp cells in P0. It was previously reported that different culture media affect growth characteristics, surface marker distribution and chondrogenic differentiation of human bone marrowderived mesenchymal stromal cells. In this study though the harvest and expansion media used was the same for all the three different sources, the type of source of mesenchymal stem cells and the quality of explants were speculated to be contributing factors for differential growth kinetics documented.
Rebelatteo et al. (2008) compared MSCs isolated from Bone Marrow (BM), Umbilical Cord Blood (UCB), and Adipose Tissue (AT) and presented their similar capacities for chondrogenic and osteogenic differentiation with marked variation in their adipogenic differentiation potential. However, in our results, though we document the similar transdifferentiation of chondrogenic, osteogenic and adipogenic capabilities with no statistical differences observed when quantified (data not presented), variation was observed in myogenic invitro transdifferentiation of adipose, dental pulp and cord tissue derived MSCs. The dental pulp source at P4 yielded 22% of dystrophin positive expression enumerated from the microscopic observation and averaging of the count from foci in comparison to both the adipose and cord tissue sources. To the best of our knowledge, this distinguishing transdifferentiation myogenic capability among the sources compared was not published before. Several groups have evidenced the tissue specific terminally differentiated cells and stem cell impregnated biocompatible 3-D gels for functional regeneration and repair of the diseased tissues and in our study, we evaluated the biocompatibility of PEGF-FB gel with MSCs derived from all the three different sources in terms of the cellular viability at different concentrations of the gel. We were able to document that at 7 mg/ml of the gel used, the MSCs were uniformly distributed within the biomaterial and the source of cells and window period after polymerization with the cells up to 3 hr at room temperature has no significant or detrimental effect on the cellular viability and the integrity of the complex made. The choice of PEGF-FB as the biomaterial for impregnating the therapeutic MSCs in our study was based on the available data on the clinical trial conducted on just the hydrogel in Europe for cartilage defects. The main purpose of this specific objective on evaluating the PEGF-FB biocompatibility with the MSCs was to simulate the clinical setup to handle the clinical grade cells and the biomaterial for transplantation in muscular dystrophy pathology.
The significance of our results lie within the gamut of study that the clinical application of MSCs for a specific indication may be dependent on the availability of the clinical source, scale up process, proof of concept invitro experiments justifying the regenerating prowess of MSCs and providing post transplantation support. While the laboratory test results of the comparative study assessing the eligibility of sources like adipose, dental pulp and cord tissue has shown that dental pulp source has more proclivity towards myogenic transdifferentiation, the harvest ease and the biopsy size are also significant factors. Umbilical cord tissue and Adipose sources are the most available sources for large scale preparations for allogenic preparations in the cell therapy procedures for diseases where the damaged or degenerating tissues are repaired.
We are grateful to Ambrish Kapadia, Anand Shah, Purvi Shah and Jiten from Parents Society of Muscular Dystrophy, Mumbai for providing partial funding to conduct experiments on adipose derived stem cells expansion, transdifferentiation and to procure the Gel from the supplier. We thank Surya Emani (Exeler Enterprises) for helpful discussions and comments on the manuscript.