Cell & Developmental Biology
Open Access
ISSN: 2168-9296
ISSN: 2168-9296
Review Article - (2013) Volume 2, Issue 4
I challenge the widely held assumption, reflected in current classifications, that larvae and corresponding adults evolved from common ancestors and that animals with lophophores evolved from an ancestral lophophorate. Types of animal development are defined, and their origins are discussed in relation to the common ancestor hypothesis and hybridogenesis, the proposal that new life forms have been generated by hybridization. Echinoderms are commonly classified by their larval characteristics as bilateral enterocoelous deuterostomes. However, some echinoderms without larvae develop as radial schizocoelous protostomes, which would place them in a different superphylum from those with larvae. Current classifications also place some hemichordates in a different superphylum from other hemichordates. A genetic study concludes that lophophorates are not descended from a common ancestor, thus invalidating the clade Lophotrochozoa. I propose that lophophorates and barnacles are chimeras, with components from different phyla. The evidence consistently supports the view that both larvae and lophophores were later additions to life histories. F M Balfour (1851-1882) recognized that larvae were later additions to life histories, and there were no echinoderm larvae until after the establishment of the classes of that phylum. He has been ignored, and, as a result, animal taxonomy has been on the wrong road since the late 19th century.
<Conventional assumptions on the evolutionary origins of larvae and lophophores have led to fundamental flaws in inferred relationships between animal phyla. These are reflected in current classifications, which place a minority of echinoderms in a different superphylum from the majority, and one class of hemichordates in a different superphylum from other hemichordates. In this paper I review hypotheses on the origins of larvae, lophophores and barnacles, and I call for a radical reassessment of higher-group animal taxonomy.
Darwin explained much of organic evolution in terms of “descent with modification through natural selection” [1]. He assumed that all evolution is gradual, and that larvae had gradually evolved from the same stocks as corresponding adults. Balfour, by contrast, proposed that virtually all larvae were later additions to life histories, and they had been ‘transmitted’ from unknown sources [2]. He deduced that there were no echinoderm larvae until after the establishment of the classes of that phylum. Most modern biologists follow Darwin and ignore Balfour, but there is a wealth of evidence that supports Balfour’s views. I claim that the phylogenetic origins of larvae and lophophores have much in common, and both were later additions to life histories. The origins of the various types of development that animals may experience during their life histories are here discussed in relation to the common ancestor hypothesis and the larval transfer hypothesis.
The common ancestor hypothesis assumes that larvae and corresponding adults evolved from common ancestors. It was accepted without question by Darwin [1], and Haeckel used it as the basis for his ‘biogenetic law’, which asserts that ontogeny recapitulates phylogeny and larvae represent ancestral adults [3]. Garstang amended Haeckel’s biogenetic law by claiming that larvae represent ancestral larvae, not adults [4]. Today the common ancestor hypothesis with Garstang’s amendment is widely accepted. Bilateral echinoderm larvae metamorphose into radial adults. Haeckel’s explanation of this phenomenon is that ancestral echinoderms were bilateral and that echinoderm larvae have remained so while the adults evolved radial symmetry as an adaptation to sessile life.
The larval transfer hypothesis states that larvae were later additions to life histories. They originated as adults in other taxa, and their genomes were transferred by hybridization. More than a century after Balfour, and after 40 years of accumulating examples of animal development that defy explanation in terms of the common ancestor hypothesis, I, like Balfour, deduced that the basic forms of most larvae were later additions to life histories, and, contrary to Haeckel, ancestral echinoderms were radial, and there were no echinoderm larvae until after the establishment of echinoderm classes.
An echinoderm then acquired bilateral larvae by hybridizing with an enteropneust hemichordate with tornaria larvae, and further hybridizations led to the spread of bilateral larvae to most echinoderms [5]. I subsequently proposed that the basic forms of all larvae originated as adults in other taxa and their genomes were transferred by hybridization [6-8]. This would have required less than ten hybridizations between members of different phyla, and rather more between more closely related animals. Larval transfer is a type of saltational evolution, but it is an addition to gradual evolution, not a substitute for it. Larvae and adults have evolved gradually, and continue to do so, as Darwin described. All developmental stages of all organisms may be subject to natural selection, however they evolved.
Some items in this section are defined in more detail in Williamson [9]. Hybridogenesis is the generation of new life forms and new life histories by hybridization, which is the interbreeding of different organisms at all levels of relationship. It includes larval transfer (defined above) and component transfer, the transfer of genetic prescriptions for parts of animals (e.g., lophophores) by hybridization.
Animals that do not pass through a larval phase are said to undergo direct development, in contrast to those with one or more larval phases, which undergo indirect development. A larva is a hatched immature animal that must metamorphose to enter the next phase in its life history. The larval form usually disappears completely at metamorphosis. Metamorphosis is a marked change in form, but what constitutes a marked change is largely subjective. Different methods of metamorphosis are discussed later. An embryo is an unhatched form, still within the egg membrane. A protomorph is the hatched form of some arthropods that becomes part of the juvenile and adult. A paralarva occupies a different habitat and shows limited morphological differences from the juvenile. A juvenile is an immature form that grows into a mature adult without metamorphosis.
Under the larval transfer hypothesis, larvae and adults each have their own genomes, and metamorphosis is the change-over from the expression of the larval genome to the expression of the adult genome. If there is more than one larval phase in the life history of an animal, metamorphosis is the change-over from the expression of one larval genome to the expression of either the next larval genome or the adult genome.
A number of terms have been employed to describe the development of insects, but they have seldom been applied to comparable types of development in other taxa. Some insects metamorphose in a relatively inactive phase known as a pupa, in which the larval tissues and organs break down. The adult digestive and circulatory systems grow from the stem cells that result from the breakdown of larval tissues and organs, but the nervous system and outer parts of the adult grow from imaginal discs, which have never been part of the larva. The origin of imaginal discs is discussed in Williamson [8].
An imago is a mature insect. Butterflies, flies, beetles and other insects that metamorphose in the pupal phase are said to be holometabolous, while dragonflies and other insects that have larvae but not pupae are said to be hemimetabolous, and species with no larvae, such as thysanurans and other wingless hexapods, are ametabolous. Some insects have two or more types of larvae in their life histories, and they metamorphose more than once. They are said to be hypermetamorphic, and they undergo hypermetamorphosis. The aquatic larvae of hemimetabolous insects are called nymphs or naiads, but the term ‘nymph’ is also frequently used for the immature stages of cockroaches, locusts, bugs and other ametabolous insects with gradual development.
Some beetles (Coleoptera), lacewings (Neuroptera) and caddisflies (Trichoptera) have campodeiform larvae, named after Campodea, a genus of diplurans (two-pronged bristletails). Such larvae resemble Campodea or other diplurans. Most types of development that occur in insects also occur in some other taxa. In some bryozoans, for example, the planktonic larva settles and undergoes histolysis and cytolysis until no larval organs or tissues remain. The resulting stem cells then divide and differentiate to produce the sessile juvenile. Such bryozoans could be said to be holometabolous, but, in practice, this term is restricted to insects. The occurrence of two larval phases in one life history is not uncommon in animals other than insects, and euphausiid, sergestid and penaeid shrimps (Crustacea) usually pass through four larval phases before the juvenile. Such animals could be said to undergo hypermetamorphosis, but, in practice, this term also is restricted to insects.
Metamorphosis is obviously not confined to insects, but some of the best known examples concern this group. The various types of insect development are all explicable in terms of larval transfer, but they pose problems for common ancestry. There is, for example, no current explanation of how the pupal method of drastic metamorphosis could have evolved gradually, as required by the common ancestor hypothesis, and other cases difficult to explain under common ancestry are discussed below. Under the larval transfer hypothesis, one or more ancestors of holometabolous insects acquired larvae by hybridizing with onychophorans, also known as velvet worms [10]. The onychophoran genome was expressed first, as a caterpillar-like larva. The onychophoran tissues and organs were too different from insect tissues and organs to permit smooth metamorphosis, but at least one hybrid devised the complex but effective pupal method of ‘start-again’ metamorphosis, in which no larval tissues or organs change directly into adult tissues and organs. The descendants of the insect/onychophoran hybrid evolved, and in some cases larval evolution included reduction and loss of appendages to produce grubs and maggots as the larvae of some types of insects.
Under the larval transfer hypothesis, one or more ancestors of dragonflies (Odonata) and other hemimetabolous insects acquired larvae by hybridizing with thysanurans (three-pronged bristletails). In this case, however, there was less difference between the animals that hybridized, and pupation and start-again metamorphoses were unnecessary. Common ancestry has no explanation of the similarities between dragonfly larvae and thysanurans. It is not an example of convergent evolution, because the larva does not benefit from the resemblance, and nature would not select for it.
Animals with more than one larval phase in their ontogenies are particularly interesting in the present context. The American striped blister beetle, Epicauta vittata (family Meloidae), passes through three larval phases and two pupal phases before adulthood (Figure 1) [11]. The egg hatches as a campodeiform larva called a triungulin. After several moults, the triungulin matamorphoses into the second type of larva, which resembles that of a caraboid beetle. The last stage of the caraboid larva pupates into a pseudopupa, also called a coarctate larva. The third type of larva, which resembles that of a scarabaeoid beetle, emerges from the pseudopupa, and it also pupates. The second pupa gives rise to the adult beetle. If, as Darwin maintained, larvae hold infallible clues to classification, E. vittata is concurrently a scarabaeoid beetle, a caraboid beetle, and a dipluran. The common ancestor hypothesis has no explanation of this very indirect development, with two pupal phases. Under the larval transfer hypothesis, the Meloidae and the Scarabaeidae evolved from a common ancestor with scarabaeoid larvae. These larvae, I suggest, had evolved from caterpillars, which originated as transferred onychophorans. The scarabaeoid larva pupated to metamorphose. An ancestor of Epicauta with scarabaeoid larval and pupal phases then hybridized with a caraboid beetle, thus acquiring genetic instructions for a caraboid larva, pupation, and an adult caraboid. The form of adult caraboid beetle is not expressed, but its genes should be sought in the non-coding DNA of E. vittata. A later ancestor of Epicauta then hybridized with a dipluran, and it thus acquired campodeiform larvae (triungulins). The triungulin is sufficiently similar to the caraboid larva, which follows it in ontogeny, to allow metamorphosis without pupation.
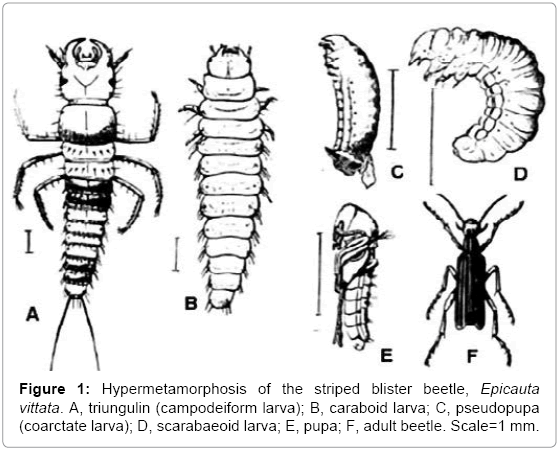
Figure 1: Hypermetamorphosis of the striped blister beetle, Epicauta vittata. A, triungulin (campodeiform larva); B, caraboid larva; C, pseudopupa (coarctate larva); D, scarabaeoid larva; E, pupa; F, adult beetle. Scale=1 mm.
The occurrence of campodeiform larvae bears no relation to the classification of adult insects. This is unexplained in terms of common ancestry, but it is readily explicable in terms of larval transfer by chance hybridizations between diplurans and a variety of insects [7].
If larvae and corresponding adults had evolved from common ancestors, only one genome would be involved in any life history. If, on the other hand, the basic forms of all larvae were transferred by hybridization from animals in other taxa, the genomes of animals with larvae, or whose ancestors had larvae, would have evolved from hybrids with a mixture of two or more genomes. Geneticists are urged to consider these possibilities in their analysis of genomes. Several examples that are unexplained under the common ancestor hypothesis but are explicable in terms of larval transfer are discussed below.
Lack of larvae may either be an ancestral or a derived condition, and echinoderms include examples of both. Adult echinoderms are radially symmetrical (Figure 2A) [12], but echinoderm larvae are bilaterally symmetrical (Figure 2B-F). In echinoderm larvae, the blastopore does not become the mouth (deuterostomy), and the coelom forms from pouches in the archenteron (enterocoely) (Figure 2B). Echinoderms with no free larvae but whose late embryos show bilateral symmetry, deuterostomy or enterocoely have almost certainly evolved from ancestors with larvae. Some such cases show indications of bilateral symmetry, even when there is no embryonic alimentary system and the terms protostome and deuterostome are inapplicable. By contrast, other direct developing echinoderms not only show no trace of bilateral symmetry at any stage of development, but the blastopore becomes the mouth (protostomy), and the coelom forms from splits in the mesenchyme (schizocoely).
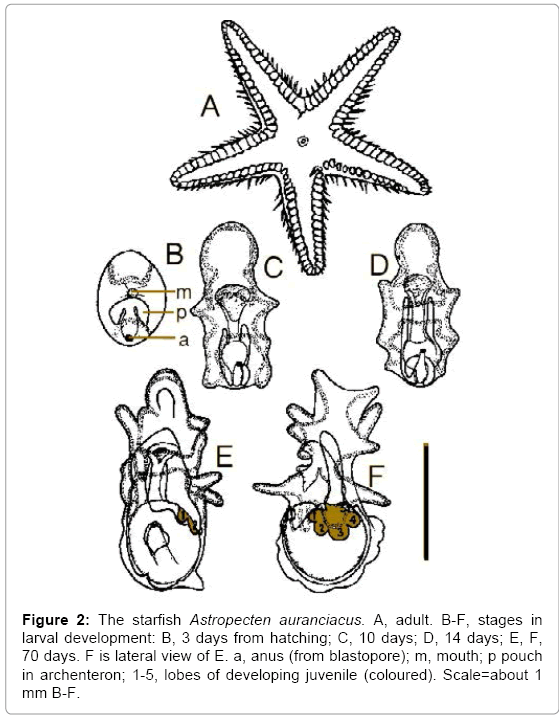
Figure 2: The starfish Astropecten auranciacus. A, adult. B-F, stages in larval development: B, 3 days from hatching; C, 10 days; D, 14 days; E, F, 70 days. F is lateral view of E. a, anus (from blastopore); m, mouth; p pouch in archenteron; 1-5, lobes of developing juvenile (coloured). Scale=about 1 mm B-F.
Kirk’s brittlestar, from New Zealand, is an egg-laying species in this category (Figure 3) [13], while the sub-Antarctic species of Abatus (Echinomorpha, Spatangoidea) and sea-daisies of the genus Xyloplax (Asteromorpha, Concentricyclomorpha) are brooding echinoderms that, like Kirk’s brittlestar, develop as radial schizocoelous protostomes [14,15]. The proposal that these echinoderms have either retained the ancestral method of development or have reverted to it offers the only known explanation of the fact that some echinoderms develop directly as radial schizocoelous protostomes while all echinoderm larvae are bilateral enterocoelous deuterostomes [7].
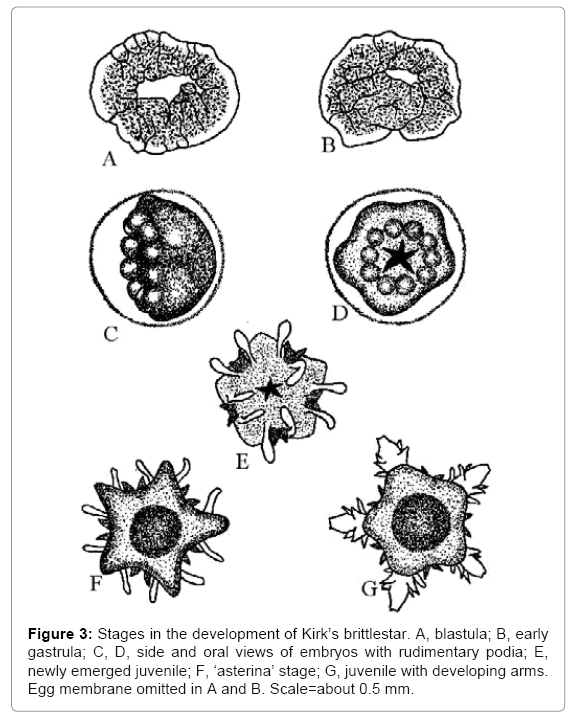
Figure 3: Stages in the development of Kirk’s brittlestar. A, blastula; B, early gastrula; C, D, side and oral views of embryos with rudimentary podia; E, newly emerged juvenile; F, ‘asterina’ stage; G, juvenile with developing arms. Egg membrane omitted in A and B. Scale=about 0.5 mm.
Cephalopod molluscs do not undergo metamorphosis during development, and they have no true larvae. The adults are carnivores, but some hatch as planktivorous paralarvae, which lack some adult features. For example, the bilobed tail fin of the squid Grimalditeuthis bonplandi (Figure 4) [16] is absent in newly hatched paralarvae, and it then develops gradually.
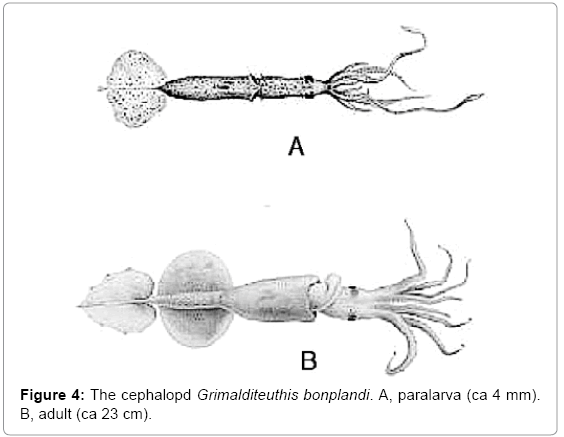
Figure 4: The cephalopd Grimalditeuthis bonplandi. A, paralarva (ca 4 mm). B, adult (ca 23 cm).
The term paralarva was originally coined for cephalopod examples [17], but it is equally applicable to the first free-living form of Branchiostoma (Amphioxus), in which the mouth is turned to the left and there is a single row of gill slits on the right side. Branchiostoma gradually becomes symmetrical as it grows. On my opinion, paralarvae did not result from larval transfer, and the paralarva, juvenile and adult are expressions of the same genome.
The larval transfer hypothesis maintains that larval forms were acquired by hybridization, and the genomes of animals with indirect development are derived from the combined genomes of two or more ancestral animals. I have, however, drawn a distinction between true larvae, which disappear at metamorphosis, and protomorphs, which are also hatched forms that differ from corresponding adults but persist throughout later life [15]. Nauplii provide examples of protomorphs and larvae.
Nauplii are unsegmented arthropods with three pairs of appendages and usually a small median eye. The first pair of appendages are uniramous, the others biramous. Metanauplii are late nauplii with the same functional appendages as nauplii, but also with rudiments of some somites and appendages that will become functional in the next developmental phase. Müller and Walossek [18] and Zhang et al. [19] used the term metanauplius for a number of nauplius-like Cambrian forms with more than three pairs of functional appendages. This, however, is a different concept from the original definition of metanauplius, and it requires a different name. ‘Paranauplius’ has been suggested [20]. I claim that some nauplii are or were adults, some protomorphs and others larva [15].
Müller and Walossek [21] described several species of Cambrian nauplii, but they found no Cambrian metanauplii, and all members of the same species were of uniform size. The lack of metanauplii and growth stages is consistent with the proposal that these nauplii were adult members of the Naupliomorpha, a class of non-crustacean arthropods [6]. I suggest that Palaeozoic adult nauplii occasionally hybridized with crustaceans, and the nauplius became either a protomorph or a larva in the hybrid and its descendants.
Many branchiopod crustaceans hatch as nauplii that persist in later life, as in Leptestheria syriaca (Figure 5) [22]. This animal grows through a series of moults, with each moult adding segments to the body, and the change in form is gradual. The nauplius is retained as the anterior part of the body, but the third pair of appendages becomes gnathobase mandibles. In Leptestheria and other branchiopods the nauplius is a protomorph, rather than a true larva. Comparable gradual development from protomorphs occurred in the Cambrian species Martinssonia elongata and related forms [23] and in Palaeozoic trilobites. Martinssonia and trilobites were non-crustacean arthropods. Martinssonia hatched as a paranauplius, with one pair of uniramous and three pairs of biramous appendages. Trilobites hatched as protaspides, each with one pair of uniramous and (probably) four pairs of biramous appendages [9]. Protomorphs are unknown in animals other than arthropods.
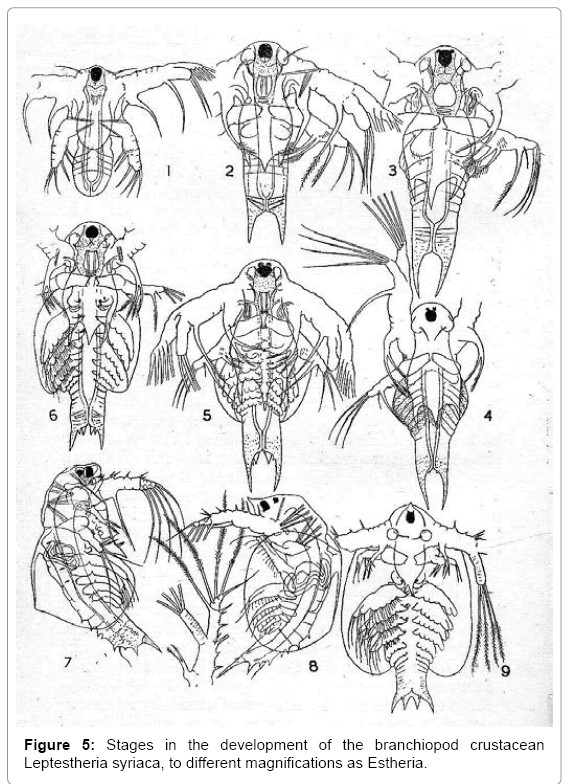
Figure 5: Stages in the development of the branchiopod crustacean Leptestheria syriaca, to different magnifications as Estheria.
Development in most arthropods with larvae includes a wellmarked metamorphosis at a single moult, in which the larval form is completely lost. Some larval features, however, are retained when a copepod metamorphoses from the last nauplius stage to the first copepodid (juvenile). The nauplius eye persists, and the second pair of appendages continues to generate swimming or feeding currents.
Under larval transfer, the genomes for protomorphs or larvae were later additions to the genomes for adults. Animals with protomorphs are concurrent chimeras, in which the two genomes are expressed together. Animals with larvae are sequential chimeras, in which the one or more larval genomes are expressed before the adult genome, and each developmental phase is separated from the subsequent one by a metamorphosis.
Balfour deduced that larvae were later additions to life histories, and there were no echinoderm larvae until after the classes of that phylum had evolved [2]. It is now known that all echinoderm classes were established by the end of the Ordovician [24]. Balfour’s conclusion gives no indication of when larvae were acquired after the classes had evolved, but there are examples of recently acquired echinoderm larvae [7]. Attention was drawn earlier to some direct developing brittlestars, heart-urchins and starfish that develop as radial schizocoelous protostomes. These have either retained the ancestral method of development or they have reverted to it. I have suggested that Balfour’s deduction on echinoderms is equally applicable to all animals with larvae and no animal in any phylum with classes acquired larvae until after the classes of that phylum were established [9]. I maintain that early hybridizations resulted in protomorphs in some cases and component transfers in others. Later hybridizations, after the classes of the respective phyla were established, produced animals with larvae. I suggest that there is a limit to the number of genes that can be expressed simultaneously.
Early animals had fewer genes than later animals, and the original genome and an addition could be expressed concurrently. Later animals had less spare gene capacity, and additional genomes could only be expressed sequentially, as larvae. I postulate that all or most animal phyla originated in the Cambrian explosion, when many concurrent chimeras resulted from hybridizations between disparate early animals [9]. The Cambrian explosion might, eventually, be depicted as a complex reticulum in which many lineages fuse, but it cannot be portrayed solely as one or more bifurcating trees.
Some examples from the Burgess Shale of Cambrian animals that show features of two or more modern phyla (Figure 6A-D) [25-28]. Laggania and Anomalocaris (Figure 6A and B) were originally regarded as members of a hitherto unknown phylum [29], but Collins proposed a new arthropod class, the Dinocarida, to accommodate them and some related Chinese Cambrian forms [30]. Whether they fall within the Arthropoda depends on how this major taxon is defined, but the gaping mouth with inner ‘teeth’ resembles that of a chordate cyclostome rather than an arthropod. Amiskwia sagittiformis (Figure 6C) had a mollusc-like head and a chaetognath-like body. Nectocaris pteryx (Figure 6D) had cephalopod-like tentacles and eyes, a bivalved shell and a chordate body. Laggania, Anomalocaris, Amiskwia and Nectocaris are extinct chimeras, with components from different phyla, and barnacles (Cirripedia: Thoracica) are living chimeras.
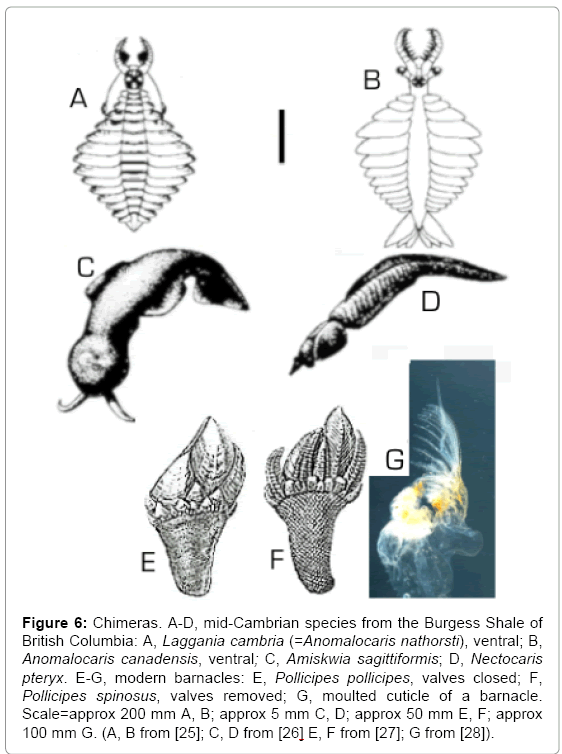
Figure 6: Chimeras. A-D, mid-Cambrian species from the Burgess Shale of British Columbia: A, Laggania cambria (=Anomalocaris nathorsti), ventral; B, Anomalocaris canadensis, ventral; C, Amiskwia sagittiformis; D, Nectocaris pteryx. E-G, modern barnacles: E, Pollicipes pollicipes, valves closed; F, Pollicipes spinosus, valves removed; G, moulted cuticle of a barnacle. Scale=approx 200 mm A, B; approx 5 mm C, D; approx 50 mm E, F; approx 100 mm G. (A, B from [25]; C, D from [26] E, F from [27]; G from [28]).
Barnacles have been classified as crustaceans (phylum Arthropoda) since about 1830, but they are only partly crustacean. Arthropods have chitinous cuticles, which they moult to grow, but the capitular plates of barnacles (usually of calcite) and associated tissues, the base, and (in pedunculate barnacles) the stalk do not have chitinous cuticles, and they grow without moulting. A barnacle with closed valves (e.g. Figure 6E) shows only non-chitinous non-arthropod features, but within the valves is a shrimp-like animal (Figure 6F), which periodically moults its chitinous cuticle. Darwin said, “Even the illustrious Cuvier did not perceive that a barnacle was, as it certainly is, a crustacean; but a glance at the larva shows this to be the case in an unmistakable manner” [1: 440]. I say, ‘Even the illustrious Darwin did not perceive that only part of a barnacle was, as it certainly is, a crustacean; but a glance at the cast cuticle shows this to be the case in an unmistakable manner’.
The moulted cuticle (Figure 6G) shows only the shrimp-like part of a barnacle, and all outer parts are conspicuous by their absence. I propose that the original barnacle was a hybrid between a shrimp-like arthropod and a sessile beaker-shaped non-arthropod with hard plates near the open end. The genomes of the two animals that hybridized were expressed together to create a concurrent chimera. Descendants of this hybrid acquired nauplius larvae (but not the cypris stage) by a further hybridization. I suggest that the juvenile of the crustacean that hybridized with a non-arthropod to create a cirripede is today represented by the non-feeding cypris. This form persists through metamorphosis, and it is, therefore, a type of protomorph rather than a larva. At metamorphosis, the cypris moults to the next developmental stage, with a functional alimentary system, and the non-arthropod part of the hybrid genome is expressed for the first time. Thereafter the crustacean part moults to grow, and the non-arthropod part grows without moulting. The Cambrian fossil Priscansermarinus barnetti was described as a probable lepadomorph barnacle [31]. It resembles a modern Lepas, with one pair of capitular plates, but the inner parts are not well preserved in known specimens. If it has no crustacean components within the capitulum, one of its relatives could have hybridized with a crustacean to produce the first barnacle.
Animals with lophophores are also chimeras. Lophophores are feeding organs consisting of a ring, a horseshoe-shape, or a coil of hollow ciliated tentacles surrounding the mouth. They occur in bryozoans, phoronidans, brachiopods and pterobranch hemichordates, and similar organs are present in entoprocts. No one has suggested a phylogeny deriving these lophophorate taxa from a common ancestor, and even if consideration is limited to brachiopods, phoronidans and bryozoans, LSU and SSU sequences of ribosomal RNA indicate that “the Lophophorata is not a monophyletic entity” [32]. There is thus strong evidence that lophophores were not inherited from a common ancestor, but the genes that prescribe them could have been acquired by distantly related animals from the same source by hybridization. The derivation of lophophores from coelenterates is discussed later. Some bryozoans go through an inactive ‘pupal’ phase in their development, with presumptive lophophore cells in a different capsule from presumptive body cells (see below). This suggests that, in this phylum at least, lophophore genes are a separate entity within the genome.
Animal metamorphosis covers three main methods of changing form during development. These are smooth metamorphosis, overlapping metamorphosis and start-again metamorphosis. Smooth metamorphosis occurs when the cells of most larval tissues and organs are incorporated into the juvenile without reverting to stem cells. Examples include (a) metamorphosis from a tornaria larva to an enteropneust hemichordate (Figure 7A-D) [33], and (b) from a Müller’s larva (a type of trochophore) to a planarian flatworm. The first example contrasts with the drastic metamorphosis from a bipinnaria larva (which resembles a tornaria) to a starfish. The second example contrasts with the equally drastic metamorphosis from a trochophore larva to a polychaete worm, a mollusc, an echiuran, a sipunculan or a nemertean. Garstang described the smooth metamorphosis from Müller’s larva in verse [34]:
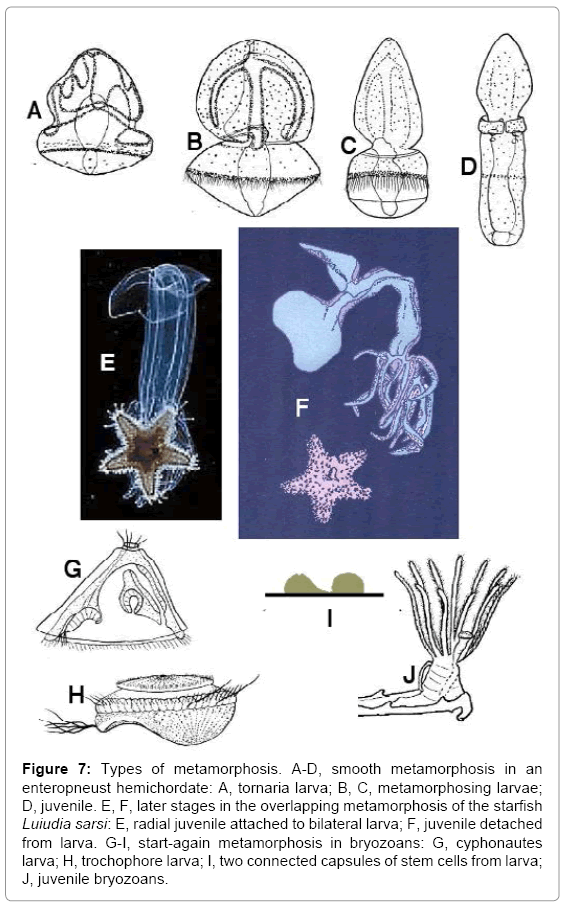
Figure 7: Types of metamorphosis. A-D, smooth metamorphosis in an enteropneust hemichordate: A, tornaria larva; B, C, metamorphosing larvae; D, juvenile. E, F, later stages in the overlapping metamorphosis of the starfish Luiudia sarsi: E, radial juvenile attached to bilateral larva; F, juvenile detached from larva. G-I, start-again metamorphosis in bryozoans: G, cyphonautes larva; H, trochophore larva; I, two connected capsules of stem cells from larva; J, juvenile bryozoans.
Johannes Müller’s larva is the primal Trochophore
That shows how early worms grew up from fry in days of yore:
No drastic metamorphosis! -- Each youngster keeps her skin:
Her larval frills are not thrown off, but eaten from within.
I, however, claim that all larvae originated as adults in other taxa, and the ‘primal trochophore’ was a rotifer [7].
All echinoderms with larvae undergo overlapping metamorphosis. The pentaradial juvenile grows from stem cells lining one of the coelomic sacs of the bilateral larva (Figure 2E and F). The developing juvenile migrates to the outside of the larval body (Figure 7E), where it can move its arms (in brittlestars and starfish), and spines and tube-feet (in all echinoderms), quite independently of the swimming movements of the larva. In most echinoderms the larva with attached juvenile settles at this stage, and much of the dying larva is absorbed by the juvenile. In the starfish Luidia sarsi, however, the juvenile drops off the swimming larva, and the two phases, from the same egg, can live separately for at least a further three months (Figure 7F) [35]. The only known explanation of this phenomenon is in terms of In terms of larval transfer. This maintains that the genome of every animal with a larval phase is a combination of two genomes, inherited from a hybrid. The development of Luidia sarsi illustrates that the two genomes may show remarkable independence. Overlapping metamorphosis also occurs in doliolid urochordates with tadpole larvae, in polychaete annelids with trochophore larvae, and in nemerteans with pilidium larvae [7,15]. In each case the juvenile breaks free from the larva, and both phases live separately for a while.
Start-again metamorphosis occurs when all larval cells revert to stem cells, which then differentiate into juvenile or adult tissues and organs. Examples from holometabolous insects were discussed earlier, and bryozoans provide further examples. Phylactolaematan and stenolaematan bryozoans have short-lived larvae or none, but most gymnolaematan bryozoans hatch as either cyphonautes larvae (Figure 7G) or trochophore larvae (Figure 7H), both of which can remain planktonic for weeks or months. A cyphonautes has a triangular bivalved inarticulate chitinous shell, in contrast to the unshelled trochophore, and the cilia are arranged very differently in the two types of larvae. The occurrence of two dissimilar types of larvae within one class of animals is unexplained in terms of common ancestry of adults and larvae, but it is consistent with larval transfer, which postulates that larvae were later additions to life histories, transferred by hybridization. The source of cyphonautes larvae, however, is currently undiscovered. Both cyphonautes and trochophore bryozoan larvae undergo startagain metamorphosis to transform to juveniles. The larva of whichever type settles, and the larval cells revert to stem cells. These cells form two interconnected capsules (Figure 7I), one of which develops into the main body of the bryozoan, the other into the lophophore [36]. This observation shows that the lophophore can show considerable independence from the rest of the animal.
The late Lynn Margulis (1938 - 2011) (pers. comm.) proposed that the source of lophophores was one or more coelenterates that had not yet acquired cnidae to become cnidarians. The acquisition of cnidae by coelenterates is discussed in Shostak and Kolluri [37]. The occurrence of lophophores as feeding organs probably resulted from hybridizations between such coelenterates and several distantly related animals during the early Palaeozoic. As postulated above, early hybridizations produced protomorphs or component transfers, while later hybridizations, after the classes of the major phyla were established, generated animals with larvae. The occurrence of lophophores in several unrelated phyla is an example of early hybridizations that produced component transfers, while the occurrence of cyphonautes and trochophore larvae in one of the three classes of bryozoans is an example of later hybridizations that led to the acquisition of different types of larvae.
Hemichordates provide examples of the results of both early and late hybridizations. The phylum Hemichordata is usually regarded as consisting of the classes Enteropneusta, Pterobranchia and Planctosphaeromorpha, but I consider the Planctosphaeromorpha to be a separate phylum. Adult enteropneusts (Figure 8A) [37,38] are worm-like solitary animals, each with a trimerous body, consisting of a proboscis, a collar, and a trunk with numerous pharyngeal slits. Many species have tornaria larvae (Figure 8B), each of which is a spheroidal deuterostome with two bands of surface cilia, one convoluted and one straight. The smooth metamorphosis from a tornaria to a juvenile enteropneust was mentioned earlier. Adult pterobranchs (Figure 8C) are small and colonial, each with a spatulate proboscis, a collar bearing a prominent lophophore, and a recurved trunk with one pair of gill slits or none. The class Pterobranchia was established by Lankester for the genus Rhabdopleura, which he assigned to the Polyzoa (Bryozoa) because of its trochophore-like feeding apparatus [39]. After a detailed study, Halanych concluded that “the tentaculated arms of pterobranchs are homologous to the lophophores of brachiopods, phoronids and bryozoans” [40]. Pterobranch larvae (Figure 8D) are very different from tornarias. They were regarded as non-feeding trochophores by Hyman [41], and the recent observation that the coelom is a schizocoele [42] supports this view. If adult pterobranchs have lophophores, and pterobranch larvae are non-feeding trochophores, pterobranch hemichordates are clearly lophotrochozoans, but this fact is ignored in current classifications.
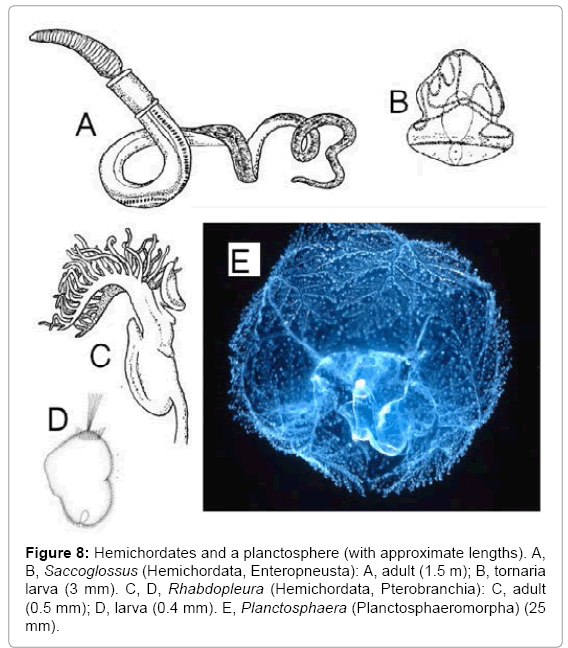
Figure 8: Hemichordates and a planctosphere (with approximate lengths). A, B, Saccoglossus (Hemichordata, Enteropneusta): A, adult (1.5 m); B, tornaria larva (3 mm). C, D, Rhabdopleura (Hemichordata, Pterobranchia): C, adult (0.5 mm); D, larva (0.4 mm). E, Planctosphaera (Planctosphaeromorpha) (25 mm).
Planctosphaera pelagica is the only known planctosphere (Figure 8E) [43]. It is usually included in the Hemichordata because it resembles a giant tornaria larva, but a specimen of 25 mm, eight times the diameter of any known tornaria, showed no sign of metamorphosis [44]. I regard it as an adult in a separate phylum, and I claim that an ancestor of Planctosphaera hybridized with a former enteropneust to give the first enteropneust with tornaria larvae [7]. I suggest that early hemichordates resembled enteropneusts without larvae. One such hemichordate then hybridized with a coelenterate, and the hybrid was a hemichordate with a lophophore: an example of component transfer. Descendants of this hybrid evolved into pterobranchs, and the phylum Hemichordata then had two classes: Enteropneusta and Pterobranchia. The enteropneust branch either had acquired the coelenterate genome, which was not expressed, or it had acquired other genes, so that when one such enteropneust hybridized with a planctosphere, the planctosphere genome was expressed first, as a tornaria larva. A pterobranch acquired trochophore larvae by hybridizing either with a rotifer resembling Trochosphaera or with an animal with trochophore larvae.
Since their description in the 19th century, lophophores have been widely assumed to be inherited from an ancestral lophophorate, but there are now several facts that question that supposition, and they are consistent with the suggestion that lophophores also were later additions to life histories. The ‘clade’ Lophotrochozoa was proposed in 1995 on evidence from 18S ribosomal RNA [45], and it has been widely accepted. In 2006, a study on LSU and SSU ribosomal genes showed that the Lophotrochozoa is not a valid clade [32], but this finding has been largely ignored. Molecular taxonomists should note that analysis of different genes can lead to diametrically opposed conclusions. K M Halanych was one of the geneticists who set up the Lophotrochozoa in 1995, and he was also one of those who invalidated it in 2006. He has clearly changed his mind. The Lophotrochozoa was erected and demolished on evidence from ribosomal RNA, quite independently of the component transfer hypothesis, but component transfer offers a solution to the current confusion.
More evidence on component transfer would probably be gained by experimental hybridizations between animals with lophophores and others without, or between animals with different types of lophophores. More evidence on larval transfer would probably be gained by experimental hybridizations between animals with larvae and others without, or between animals with different types of larvae. Crosses between ascidians and sea-urchins are described in Williamson and Boerboom [46].
Current orthodox classifications (e.g. Figure 9) [47] group echinoderms with hemichordates because the larvae of echinoderms and enteropneust hemichordates are bilateral enterocoelous deuterostomes. This, however, excludes those brittlestars, heart-urchins and sea-daisies without larvae that develop as radial schizocoelous protostomes (see above, under ‘Single or plural genomes’). A new superphylum would have to be erected for these direct developing echinoderms. Such classifications also ignore pterobranch hemichordates, which have lophophores and trochophore larvae (see above, under ‘The origins of lophophores’). This would place pterobranchs in the Lophotrochozoa, i.e., in a different superphylum from enteropneusts. This is another case of splitting a phylum into two superphyla, which is absurd. Such classifications also ignore methods of metamorphosis. The smooth metamorphosis of enteropneust hemichordates (Figure 7AD) is strikingly different from the overlapping metamorphosis of echinoderms with larvae (Figures 2B-F and 7E and F). It is highly improbable that smooth, overlapping and start-again types of metamorphosis could have evolved from a common ancestral type. The major phyla of the controversial clade Lophotrochozoa are the Annelida and the Mollusca, neither of which have lophophores nor both of which contain classes without larvae.
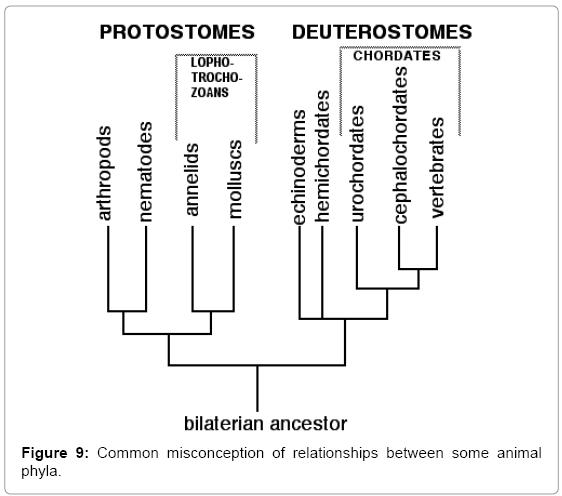
Figure 9: Common misconception of relationships between some animal phyla.
It is thus seen that current conventional classifications (e.g. Figure 9), which are based on the assumption that larvae and lophophores were both inherited from common ancestors, contain a number of illogicalities. A revised zoological taxonomy should recognize that that larvae and lophophores were later additions to life histories, introduced by genome fusions resulting from hybridizations. As a result, neither similar larvae nor presence of lophophores indicate that corresponding adults evolved from common ancestors. Ideally, this revised taxonomy should show that lophophorates and barnacles are chimeras, with components from different phyla, but, as yet, there is no concise method of displaying this. The resulting classification would be more complex than any current model, but nature does not consider the convenience of taxonomists.
Symbiogenesis is the generation of new life forms by symbiosis [48-50]. Hybridogenesis is the generation of new life forms and new life histories by hybridization [7,15]. Both are saltational forms of evolution involving mergers of genomes. They are independent of evolution within separate lineages, such as gradual evolution by “descent with modification” envisaged by Darwin [1] and evolution by larger increments, as described by Eldredge and Gould [51]. All organisms, however they evolved, may be subject to natural selection. Symbiogenesis was responsible for the creation of eukaryotic cells (of protoctists, plants, fungi and animals) from prokaryotes (bacteria and archaea), and for the acquisition of cnidae by coelenterates. Hybridogenesis was responsible for the evolution of complex animals from simple animals, for the acquisition of organs such as lophophores by some animals, and for the acquisition of larvae by many animals. Darwin [1] persuaded biologists that organisms have evolved. He also proposed a method of evolution, but this, as Darwin himself stated, is not necessarily the only method. I agree with Theissen [52] that “the complete dismissal of saltational evolution is a major historical error of evolutionary biology tracing back to Darwin that needs to be rectified”.
Charles Darwin died on 19 April 1882, aged 73, of coronary thrombosis and heart failure. Francis (Frank) Balfour died three months later, on 19 July 1882, aged 30, attempting the ascent of Mont Blanc. He had recently recovered from typhoid fever. Balfour was “regarded by his colleagues as one of the greatest biologists of his day and Charles Darwin’s successor” [53]. Despite the difference in age, the two men were friends, and they held each other in great esteem. Balfour had stayed at Darwin’s house on several occasions. Had both men co-existed for a few more months, Balfour would probably have persuaded Darwin to adopt his (Balfour’s) views on larvae and the evolution of echinoderms. Had Balfour not died on Mount Blanc, he would have taken up the Professorship of Animal Morphology, created for him by the University of Cambridge. He would have put forward, in publications and lectures, his views that larvae were later additions to life histories and no echinoderms acquired larvae until after the classes of the phylum were established. He was already impressed by analogies between tadpole larvae and adult lampreys [2], and he would probably have soon linked all larvae to adults in other taxa. Given the evidence, most biologists would have accepted Balfour’s views rather than the supposed links between evolution and development based on the assumption that larvae and adults have common ancestors. The larval transfer hypothesis would have been widely accepted by the end of the 19th century, and my publications over the last 25 years would have been superfluous.
I am very grateful to Farley Fleming for constructive comments.