Journal of Cell Science & Therapy
Open Access
ISSN: 2157-7013
ISSN: 2157-7013
Review Article - (2017) Volume 8, Issue 5
Yamanaka and Takahashi’s astonishing discovery in 2006 revolutionized the world of stem cells. Simplicity and reproducibility of IPSC’s cells opened doors for extensive therapeutic advancements and potential clinical trials particularly in the field of Regenerative medicine. In 2012, nobel prize was presented to both researchers for the breakthrough in reprogramming somatic cells to a pluripotent state with the expression of “Yamanakas cocktail” of OKSM quartet transcription factors: sex determining region y box2 (sox2), octamer binding transcription factor4 (oct4), krupple like factor (klf4) and myelocymatosis oncogene (c-myc). Rationale of iPS use is attributed to its unlimited cell source circumventing ethical hindrance. This comprehensive review will summarize mechanisms of IPS production and cell therapy applications. Moreover, appreciate the improvements and initiatives in iPS technology, scrutinize the scientific claim of indistinguishable resemblance of iPS and eSC, evaluate the constraints during iPS manipulation and analyze safety of these pluripotent cells in clinical scenarios.
Keywords: Reprogramming; Constraints; Worldwide initiatives; Induced pluripotent stem cells; Remodelin; Gene editing
iPS: Induced Pluripotent Stem Cells; hiPSC: Human Induced Pluripotent Stem Cells; OKSM: oct4, sox2, c-myc; ESC: Embryonic Stem Cells; SOX: Sex Determining Region; Ybox2, KLF4: Krupple Like Factor 4; Cmyc: Myelocytomatosis Oncogene; GMP: Good Manufacturing Practice; ECM: Extracellular Matrix; RPE: Retinal Pigment Epithelium; HLA: Human Leukocyte Antigen; CiRA: Center for iPS Research and Application; CIBK: Clinical iPS Cell Bank of Kyoto; DMF: Drug Master File
Genetic manipulation of somatic cells led to the discovery of iPS which could differentiate into all three primary germ layers (endoderm, ectoderm and mesoderm) [1,2]. IPS generated by the OSKM transcription factors appeared morphologically similar to eSC and exhibited similar gene expression profiles and selfrenewal characteristics [3-6,]. iPS can be identified by immunohistochemistry, RT-PCR and expression of immunological cell surface markers such as stage specific embryonic antigens (SSEA), Tra-1-60 and Tra-181 [1,7]
Benefits of iPS are due to cells being highly proliferative, biocompatible and universally accessible. iPS have aided in disease remodeling “disease-in-a-dish”, drug screening and designing tailored therapies for individual patients [2,8,9]. For therapeutic purposes disease specific iPS of any lineage can be generated [1,9]. Since IPSC’s are autogeneically available, both immunological compatibility and ethical constraints are bypassed [10].
Downside of iPS usage is retention of an epigenetic memory of their cell of origin which affects the differentiation process [9-12]. Somatic cell nuclear transfer and transcription factor based reprogramming methods yield induced pluripotent cells and simultaneously reset genomic methylation.
Discernible gene expression patterns and DNA methylation patterns were observed in a study on hiPSC derived from fibroblasts, adipose tissue and keratinocytes [13]. Low passage induced pluripotent stem cells derived by factor based reprograming harbor low passage residual DNA methylation signatures characteristic of their somatic tissue of origin, which promotes differentiation along lineages related to the cell of origin, while restricting alternative cell fates. Such an “epigenetic memory” of the donor tissue could be reset by differentiation and serial reprogramming, or by treatment of iPSC with chromatin modifying drugs. Another experiment corroborated this conclusion that in a given iPS clone DNA methylation patterns could indicate the origin of the former phenotype [2,14]. Furthermore, undifferentiated iPS remaining at site of interest expand into a teratoma [2,9,15]. Teratoma has become the gold standard assay to define bona fide induced pluripotent stem cells capable of generating tumoral disorganized structures containing tissues representing the three germ layers. Despite the importance of this assay to prove pluripotency the underlying mechanism of teratoma transition to teratocarcinoma in vivo is not yet understood. There is no standard procedure to test/ characterize teratomas to control puripotency. This needs to be further tested as characterizing teratomas is crucial in a therapeutic patient based iPS treatment.
Transcriptional regulators
Commencing with a pool of 24 pluripotency associated candidate genes Yamanaka identified minimally required core set of 4 genes: the OSKM cocktail to generate iPS from mouse fibroblasts through retroviral transduction [6,8,10]. However, overexpression of these regulators exhibited neoplastic potential which has been validated in various researches. Aberrant expression of oct4 is linked to hepatocellular carcinoma and murine epithelial dysplasia [10,16,17]. Sox2 overexpression has been reported in serrated polyps, mucinous colon carcinoma, human squamous cell lung tumours [18,19]. Elevated expressions of klf4 is linked to breast cancer and the cmyc oncogene has strong correlation with various human cancers [19]. This incited search for less carcinogenic gene set sufficient for reprogramming. Substitution of klf-4 and cmyc was necessary due to extensive malignant potential. Yamanakas group also identified that myc is unnecessary for generation of iPSC’s [20]. Essrb, orphan nuclear receptor, found in ES cells is reported to replace klf-4 [10,21]. P53 blockade also allowed reprogramming in the absence of klf-4. Yu et al. utilized Homeobox protein NANOG and RNA binding protein LIN-28 along with oct4 and sox2 to generate IPSC’s from human somatic cells [15]. NANOG doesn’t trigger reprogramming but is essential for acquiring state of full pluripotency [22].
Reprogramming enhancers
Frequency of iPS production is typically well below 1% imposing limitations to mechanistic studies and clinical translation. Myc family including L-myc and N-myc enhance reprogramming but is disregarded due to its oncogenetic nature. Use of Glis-1, NANOG and Sall-4 have shown to promote direct reprogramming of somatic cells [23,24]. NANOG augments the reprogramming efficiency which is quantified by NANOG positive colonies in a cell culture [21].
Furthermore, small chemical compounds which either affect chromatin modifications or transduction pathways enhance reprogramming or replace transcription regulators include i) BIX-01294, histone lysine methyltransferase inhibitor [25]. ii) TSA and SAHA, deacetylase inhibitors [26]. iii) AZA and RG108, DNA methyltransferase inhibitors [26,27] iv) BayK8644, L-type Calcium channel agonist [25] v) Dexamethasone, steroid glucocorticoid [26]. vi) PD0325901 and CHIR99021, inhibitors of the MEK and GSK respectively [28,29]. vii) A-83-01, TGF β inhibitor [30].
Mechanism of iPS Generation
Comprises of integrating vectors, non-integrating vectors, excisable vectors and vector-free systems. Conventional viral integrating systems (retroviral transduction) were commonly employed to generate iPS from mouse fibroblasts or human somatic cells [6,7,10,31]. Retroviral integration into host genome requires high division cycles which is observed in a narrow spectrum of cells [10,32]. Retroviral vectors constitutively express transgenes under the control of promoter localized in long terminal repeat (LTR) which is terminated once the somatic cell transforms into its pluripotent state [8,10,32,33]. Partially reprogrammed cells show increased levels of exogenous pluripotent transgenes found in the retrovirus though fully reprogrammed cells exhibit constant retroviral attenuation [34]. Decrease in exogenous expression of oct4, sox2, klf4 and c-myc is reported in the retroviral construct as cell approaches pluripotency correlating with a decline in reprogramming efficiency. Due to above mentioned pitfalls, lentiviruses have been utilized for iPS generation but they lead to a differentiation blockade since they are less efficiently silenced in pluripotent cells than retroviruses [9,13]. Doxycycline-inducible lentiviral vectors have aided in selection of fully reprogrammed cells since drug withdrawal halts proliferation of exogenous factor dependent cells [33,35]. Nevertheless, issues of insertional mutagenesis and leaky transgene expressions due to retroviruses which precipitate harmful effects if considered in a therapeutic in vivo setting led to exploration of non-integrating (viral-free) systems for iPS generation [6,8,10].
Non-integrating systems
Viral integrating methods of iPS generation are unsuitable in a clinical setting. With application of adenoviruses, sendai viruses, expression plasmids, episomal vectors, liposomal magnetofection and minicircle vectors various virus free iPSC lines have been formed by passing the risk of insertional mutagenesis. Although non-integrating systems are safer than conventional use of viruses for delivery of reprogramming factors this system has limited application due to low grade efficiency [36-42].
Excisable vectors
Cre-loxP technology: The Cre-loxP system provides an optimized protocol for generating transgene free human induced pluripotent cells with retroviral transduction of a single vector consisting of the coding sequence of transcriptional regulators linked via picornaviral 2A plasmids [6,9,43]. Once reprogramming is achieved, this cassette is excised by mRNA transfection with Cre-recombinase. Ease and efficiency of CreloxP system with minimal genomic modification facilitates the reprogramming process of transgene free iPS. Cremediated excision of the provirus eliminates all transgenes but lox- P remnants are found at each integration site, possibly distressing the neighboring genes [44].
Piggy bac transposons: This system requires only inverted terminal repeats flanking transgene and temporary enzymatic expression which catalyzes introduction and excision of mobile genetic elements from the host genome [9,45]. Benefitting from the natural propensity of the piggy-bac systems seamless excision, it is validated that each PG insertion can be removed from fully reprogrammed iPS cell lines.
Vector free systems
Reprogramming proteins: Genomic manipulation is unavoidable in DNA-based reprogramming, so alternate systems suggested application of proteins as delivery tools for the generation of transgene-free iPS [46]. By delivering Yamanakas cocktail of transcriptional regulators fused with a cell-penetrating peptide (CPP), Kim et al. achieved efficient iPS from human fibroblasts [47]. A study on the midbrain dopaminergic (DA) neurons differentiated mouse embryonic stem cells and protein based iPS, resulting in a stable DA neuron-specific marker expression observed in the protein based iPS [48]. An unsuccessful attempt at outgrowing stem cell like colonies into iPS by reprograming proteins concluded that partial reprogramming was a common response to protein-based reprogramming technique [49]. In contrast use of self-penetrating proteins and truncated proteins seems an attractive alternative [50]. It’s unclear whether protein transduction can be used for adult cells [51]. Also, applicability of protein based iPS is restricted due to technical difficulties in production and reprogramming inefficiency.
mRNA: Warren et al. exploited synthetic mRNAs for reprogramming which resulted in higher efficiencies than the established protocol [52]. However, increased workload and the need of a tissue incubator with 02 controls are drawbacks of this system [53]. Rosa et al. resolved the issues of interferon-mediated innate immune response and poor protein yield by modifying the ribonucleotides used [54].
MicroRNAs have also shown to control pluripotency, escalating the reprogramming process [6]. MiRNA 302 is of particular importance and has been reported in various studies [55-58]. Mouse and human miR302/367 iPS cells have exhibited similar characteristics to Oct4/ Sox2/Klf4/Myc-iPS cells, including pluripotency marker expression, teratoma formation, and, for mouse cells, chimera contribution and germline contribution [59]. MiR-155 has been identified as a key player for the in vitro differentiation of iPSC toward hematopoietic progenitors [60].
These improvements in reprogramming system have helped achieve stable stem cell lines with high efficiencies. However, their potential in human iPS is still under research and careful monitoring of proteins and mRNA interaction with each transcription factor in adult iPS still needs to be evaluated.
Optimized Cell Cultures: Feed Less, Discover More
Since the revolutionary discovery began by the stem cell pioneer numerous methods were introduced and routinely undertaken to manufacture stem cell lines. However optimized conditions to achieve ideal iPS lines for human therapies are yet to be designed. Despite the tremendous potential of iPS technology, several issues with iPS manufacturing have been identified. Feeder cells for iPSC maintenance in culture have impeded the translation of this innovative technology into clinical therapy because of their xenogeneic constituents. To overcome these ethical barriers, use of synthetic coatings and bioreactors that support proliferation of iPS in xenogeneic-free environment have been suggested [2,61]. Generation of iPS from adult adipose stem cells in feeder free environment is made possible by switching to mTeSR1 culture medium [62]. Application of a cost effective, high quality E8 medium has also supported higher reprogramming efficiencies for viral and episomal approaches. Freshly isolated fibroblasts in E8-based fibroblast medium on vitronectin have consistently reprogrammed at a higher efficiency (60 to ~1,000 iPS cell colonies per 106 transfected fibroblasts) than the commercially available established stem cell lines [63]. E8 medium supporting higher reprogramming efficiencies for both viral and episomal approaches has been validated [64].
Reprogramming events require mechanical isolation of iPS for colony expansion with major contamination by unreprogrammed somatic cells. Colony picking though prone to bacterial contamination has been routinely used to enrich stem cells but becomes impractical when numerous lines are considered. Dual advantage of E8 medium and the EDTA dissociation method has unveiled a faster expansion of large cell volume cultures. Furthermore, EDTA dissociation has enabled enrichment of potential iPSC’s in an overcrowded reprogramming culture in which otherwise a routine secondary passaging is needed [64,65].
Sigma aldreich’s PluriSTEM and human ES/iPS medium is a specially formulated defined media utilizing activin-A, tgfB1 and bFGF which aids in promoting stem cell self-renewal with increased cell viability and enhanced cell proliferation in single cell passaging. This feeder-free, serum free culture system requires less cell feeding lowering down the total cost and culture period [66].
iPS lines need to be strictly generated under GMP (Good manufacturing practice) system protocols if considered for use in a clinical setting. GMP system ensures products are continuously produced and controlled according to quality standards. Thus, minimizes the risks related to pharmaceutical production that cannot be eliminated through testing the final product. To date, no fully cGMP-compliant cell line has been reported where the entire manufacturing process, from tissue sourcing to cell expansion and banking processes as well as documentation, raw materials, staff training, cell therapy facility, and quality control (QC) testing is validated [67].
Recently a biotech company, Lonza put forth a novel, robust and reproducible GMP compliant reprogramming guide which could bypass quality control impediments for efficient iPS production. Lonzas Nucleofector™ technology for efficient, non-viral delivery of reprogramming factors to the L7 Culture System for feeder-free, xenofree culture of human iPS, is currently being utilized by leading scientists all over the world [68].
Disease remodeling
Researchers worldwide are struggling to understand the mechanism of action of various debilitating diseases and remedies to fight them. Previously experimental animal models have been exploited to understand the underlying mechanism of various human pathologies. However marked differences in biochemical variation, anatomical complexity and physiological responses limit the evaluation of new therapies in humans. Mice have become the gold standard for research but simply put mice aren’t humans. Stem cells have been employed to recapitulate pathologies in vitro commonly referred to as “disease-in-adish” [69]. The iPS technology can improve our interpretation of the combined effect of genetic malfunctioning and environmental changes on the severity of diseases. Power of iPS technology lies in understanding mono and polygeneic pathologies through disease remodeling [70]. iPS allows a deeper insight by capturing the disease in its early stage and monitoring the cellular events throughout disease development [9]. Moreover, patient specific iPS can help tailor individualized therapies.
Numerous iPSC lines have been generated so far from pathological diseases e.g., Degenerative illnesses like parkinsons disease [71], alzeihmers [72] and amyotrophic lateral sclerosis [73]. Blood Disorders such as sickle cell anemia [74], fanconis anemia [75], hemophilia A [76] and β-thalassemia [77]. Genetic diseases: Downs syndrome [78], familial dysautonomia [79] and Huntington’s disorder [80].
3D organoids
3D cell clusters depend on extracellular matrix in culture for selforganization, resembling endogenous tissue architecture and duplicating organ functionality [81]. These 3D functional organoids have improvised our understanding of early human development and improved drug screening methodologies organoids including mini gut, mini heart (repopulating decellularized mice heart through iPS), mini brain (developing cortex, central telencephalon and choroid plexus), small lungs, mini eyes (optic vesicle like structures) have been formulated for disease remodeling purposes [82-89].
Lack of vascularity, absence of native interactive microenvironment, limited growth potential due to lack of constant nutrients, rigid nature of ECM, absence of immune cells limiting the observation of inflammatory response to drugs are associated pitfalls of these cell clusters [81,90].
Spinning bioreactors and shaking culture platforms have been suggested to provide better nutrient supply and improve the growth of organoids [91]. Co-culturing with endothelial cells has aided vasculogenesis in organoids [92]. In vitro Organoids are the biggest technological breakthrough that might in future overcome the dire urgency of organ transplants for critically ill patients.
Gene editing implication in disease remodeling
Discovery of site specific nucleases Zinc finger nuclease (ZFN), transcription activator-like effector nucleases (TALENs) along with CRISPR-Cas9 system has enhanced gene editing efficiency in iPSCs by rupturing the DNA double-strand at the site of gene modification enabling knock to in/knock out of one or more genes [93]. High system efficiency, ease of use and cheaper costs than predecessors has made CRISPR-CAS a popular editing tool. Researchers can introduce specific disease-causing alleles into wildtype iPSC’s with the assistance of CRISPR-CAS and eliminate genetic defects in patient iPS for isogenic controls in disease remodeling [94]. Successful progression of iPS and CRISPR-CAS amalgamation is justified by stable iPS proliferation and wholesome numbers of modified clones that can be harvested after genetic modification in culture. Immunodeficiency centromeric region instability, facial anomalies syndrome (ICF) and pancreatic cancer models have been generated using the CRISPR-CAS editing system [95,96]. Seamless gene correction of β-thalassemia mutations in patient-specific iPSCs has been possible using CRISPR/ Cas9 system [97,98].
Drug screening platform
Drug testing is an enormous challenge faced by pharmaceutical companies with a 90% failure due to unpredictable adverse effects unidentified in the preclinical phase [99]. $2billion per drug is an estimated cost of the entire process of drug screening. The average time to introduce new drug to the market, from the start of clinical testing to FDA approval, is 8.5 years, and the clinical success rate is 21.5% [100]. Researchers can take advantage of recapitulating human pathologies in vitro to evaluate cellular responses to different chemicals. Moreover, any effective drug against hiPSC can be compared with various cell lines of patients with similar pathologies to validate results. iPS are a powerful platform for drug screening to understand cellular responses to neurons, cardiocytes and hepatocytes which are most susceptible to drug toxicity [101].
Drug development employs three main strategies after establishing disease-specific iPS: high-throughput screening (HTS) of drugs, candidate drug approach or patient-specific therapy [102]. In HTS, extensive compounds are tested on differentiated cells, followed by phenotypic analysis. Making use of large bank of compounds can help identify therapies in vitro by this approach which is faster and economically less burdening than conventional drug testing. By contrast, both candidate drug approach and patient-specific therapy use small numbers of potential drugs to attenuate the disease. These approaches are useful when the disease mechanism is known and potential therapies are available.
However, substantial safety assays are required before drugs discovered by these methods can be prescribed (Figure 1).
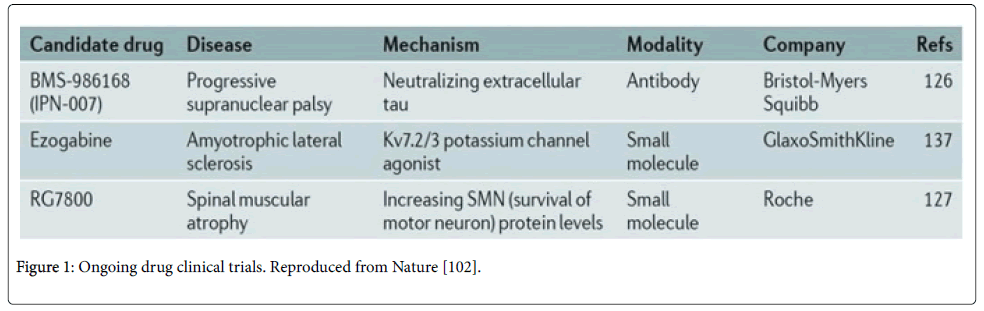
Figure 1: Ongoing drug clinical trials. Reproduced from Nature [102].
Worldwide Banking Initiatives
Tedious large-scale efforts nationally and internationally are taking place to build an iPS reservoir for therapeutic purposes. Californian institute of regenerative medicine in San Francisco initiated cell repository to bank iPS lines for research on childhood degenerative illness, eye blindness and therapeutic purposes. The first 285 lines were distributed by CIRM in September 2015. In 2013, CIRM awarded Cellular Dynamics International Inc. (CDI) $16 million to create iPS lines for each of 3,000 healthy and diseased volunteer donors across 11 common diseases and disorders to be made available through the CIRM hiPSC Repository. CDI will expand each iPS line to generate sufficient aliquots of high quality cryopreserved cells for distribution via CIRM [103].
IMI is the world's biggest public-private partnership (PPP) in the life sciences. The (EBiSC) European Bank for induced pluripotent Stem Cells was a two-year project operated by IMI. EBiSC’s goal is to establish a centralized facility where academics, biotech companies, and big pharmaceutical companies can store and access high-quality, well-characterized iPSC’s covering a range of disease areas as well as cells from healthy donors. For its part, the bank will provide standardized protocols for the storage, retrieval, culture, and differentiation into different cells types, plus a searchable catalogue where cells can be requested based on specific characteristics or disease areas [104].
Another IMI project, StemBANCC (Stem cells for biological assays of novel drugs and predictive toxicology) began in October 2012 and aims to generate 1500 iPS lines from 500 people by end of September 2017, characterize them in terms of their genetic makeup and metabolic profiles. Moreover, make them available to researchers to study various diseases including dementia and diabetes mellitus, allow drug testing and efficacy for safer drug development. All cell lines will undergo rigorous quality checks [105].
In a unique collaboration, both IMI’s EBiSC and Stem-BANCC team members incited the search to improve understanding of a rare pathology called inherited erythromelalgia (IEM). IEM is a debilitating condition without adequate available medical treatment. It involves patients suffering from extreme pain symptoms made worse by heat. Underlying etiology is a genetic mutation leading to over activity of the sodium channels in patients’ sensory neurons. Five subjects were tested with an experimental drug targeting blockade of sodium channels to reduce the pain experienced by IEM patients. Study concluded that drug reduced pain upon exposure to heat in most sufferers. Same patients allowed scientists to transform their blood cells into iPS, which were deposited in the EBiSC project stem cell bank. Next, the StemBANCC scientists transformed stem cells into sensory neurons and evaluated their characteristics. Prior to drug therapy, neurons exhibited hyper-excitability and increased response to heat: increased severity of the disease led to increased number of hyperactive neurons. Post treatment, hyper-excitability of tested neurons diminished, mimicking the drug effect that occurred in patients. Rarely is it possible for drug developers to scrutinize drug responses of the relevant cells from individual patients involved in clinical trials of the same experimental drug. These results illustrate the power of stem cell technology to transform drug development, formulating therapies tailored to individual patients. This bridges the translational gap between preclinical models and clinical evaluation [106].
In UK, human induced pluripotent stem cell initiative (hiPSCi) combines diverse constituents in genomics, proteomics, cell biology and clinical genetics to form a national high content iPS resource for interpreting cellular genetics. HiPSCi’s 2017 target is to generate over 500 iPS lines from healthy volunteers and several hundred lines from individuals with genetic disease to discover the impact of genomic variation on cell phenotype and identify new disease mechanisms.
Kyoto University’s Center for iPS Research and application (CiRA) in japan has also invested heavily in developing an iPSC bank. Just 75 iPS lines will cover 80% of the Japanese population by 2020 says Yamanaka, the leading director of (CiRA), Kyoto University [107]. Prior to clinical therapy, donor iPS can be matched to patients through human leukocyte antigens haplotyping based on cell surface proteins regulating immune responses. Yamanaka states that matched cells will obviate the need of large doses of immunosuppressive drugs causing cell rejection, the main hurdle faced during embryonic stem cell therapy for transplantation.
HLA iPS banks will ensure safer therapies since this hybrid model will select optimally matched cells producing closer to ideal graft materials, minimizing the risk of immune responses [108]. In japan, creating an HLA matched iPS bank was faster, cheaper and less tedious as few as 50 stem cell lines could cater 90% of the Japanese populations due to narrow genetic diversity [109,110]. In contrast, an iPSC bank from 150 selected homozygous HLA-typed volunteers could match 93% of the United Kingdom population [108]. Although initial efforts seem daunting with increased startup costs, HLA matched iPS cells could reduce use of lifelong immunosuppression and improve patients’ quality of life.
An international collaboration in the form of GAIT (global alliance for iPS based therapy) aims to formulate a GMP iPS-haplobank in harmony with mutually recognized iPS banks for maximizing utilization and offering hope in the future to facilitate iPS therapy globally [111-113].
These dedicated efforts for developing and improving wide spread availability of iPSC globally will help in developing innovative therapies and provide limitless opportunities for scientists to scrutinize underlying molecular mechanisms of genetic, epigenetic and environmental pathologies (Figure 2).
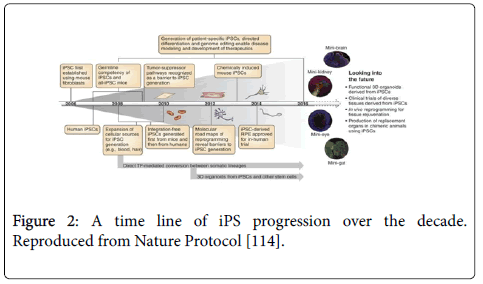
Figure 2: A time line of iPS progression over the decade. Reproduced from Nature Protocol [114].
Ips and Esc: Strikingly Similar Yet Remarkably Different
Various researches have undergone to analyze iPS and eSC lines. Great similarities emerged in histone modications and genome expressions of these two cell lines [9,115]. Comparative transcriptome analyses using microarray indicate that hESCs and hiPSCs are highly alike on global scale, with gene expression patterns clustering together [116]. Gene expression analysis study in mice revealed that majority iPS formed low grade chimeras but only cells with an ESC equivalent expression of the dlk1-dio3 locus exhibited high grade chimeras (the key indicator of pluripotency) and generated all iPS mice after blastocyst injection [9,117]. Despite the incomparable origin and generation methods, comparison of these cell lines is exceptional. This could be attributed to the man-made nature of both cell lines. Cells from the inner cell mass are to some degree manipulated under culture conditions to generate ESC which are otherwise non- existent in physiological conditions [118]. However, conflicting studies using targeted bisulfite sequencing of three human ESC clones and four iPS lines showed extensive differences in DNA methylation patterns [118,119]. Expression microarrays have been used to report various genes being differentially expressed in both stem cell lines [120]. IPS and ESC debate is still on but the important concern should be focusing on the functional characteristics of these cells in vivo as they can be categorized as part of the same big experimental approach i.e. the therapeutic potential of these cell lines in regenerative medicine in the future (Figure 3).

Figure 3: Various IPSC banking initiatives [102].
Reprogramming Constraints: Fighting the “M Triad” (Money, Mutations and Manufacturing)
The biggest hurdle in generating commercially available iPS is extensive culture period and heavy burdening investments in the entire process. A standardized protocol for cost effective production of iPS has to be carefully formulated. Complexity of iPS application in field of regenerative medicine has made manufacturing lengthy and tedious. Cutting down costs is a problematic task which needs identification of important factors which can be modified efficiently and those which are absolutely necessary for iPSC fabrication. Mahendra and Anthony suggested potential cost-effective strategies which can reduce financial investments in iPS technology substantially.
• Defining iPS as an input material since it’s not the end product in iPS therapy and should be regulated as other manufacturing products.
• Fabricating stem cell lines under Good Manufacturing Practice Protocols (already discussed).
• Developing a Drug Master File (DMF) accessible to investigators that will dispense the need of repeated testing and validation of material suppliers and assays. This would require companies to share data or permit referencing to a DMF benefitting all involved parties and cutting down costs effectively.
• Provide kits for iPS generation.
• Use modular manufacturing protocol and manufacture products from same master cell bank.
• Calibrating material and developing comparability assays by consensus.
• Widen approved use based on function of particular disease.
• Implement haplobanking.
• Simplification of the approval process already approved in other areas.
• De-risk development [111].
Other pluripotency pitfalls
Various somatic mutations, copy number variations are associated with iPS lines [121-123]. These potential complications are a hurdle in iPS therapeutic applications. Extensive genetic screening would be required to ensure safety of these cells in clinical scenarios. Genome sequencing of hiPS cell lines will help screen out cell lines with increased mutational load or mutations implicated in development, disease or tumorigenesis. Thorough studies on mutation rates and distributions during in vitro culturing and reprogramming of hiPS cells will be vital to aid in establishment of clinical safety standards for genomic integrity.
Prior to commencing the 1st ever human clinical trial with iPSC, rigorous safety trials (in mouse and monkeys) took place to validate application of retinal epithelial sheets without subsequent tumorigenesis [124,125]. The swift journey from skin to eyes started in September 2014, when ophthalmologist Masayo Takahashi collaborated with the stem cell pioneer Shinya Yamanaka and transplanted RPE (retinal pigment epithelium) sheets generated from patients iPS cells into the right eye of the female suffering from age related macular degeneration. Takahashi had claimed to stop the degeneration process and the patient experienced an improvement in vision [126]. The trial was halted due to unexplained mutations observed by Yamanaka in the transplanted RPE and the patients’ iPS [126,127]. This trial proved that several safety assays need to be established to validate preclinical studies before they are safe to begin future clinical trials. Each reprogramming stage needs to be carefully validated and monitored for future approval of any clinical trial with iPS.
Despite the pitfalls of this system iPS holds great potential for further improvement in advancing the field of regenerative medicine. If gmp compliant manufacturing obstacles, financial hurdles, reprogramming inefficiencies and genetic instabilities are overcome iPS could be tailored for individualized therapeutic products. This road to progress has been challenging yet still very promising. This world has witnessed great revolutionary advancements owing to the massive potential of these cell lines and will continue to do so in the near future.