Cell & Developmental Biology
Open Access
ISSN: 2168-9296
ISSN: 2168-9296
Review Article - (2012) Volume 1, Issue 6
The larval transfer theory challenges the assumption that larvae and adults gradually evolved from common ancestors. It claims that larvae were later additions to life histories. Instead of evolving from within the same lineages as their associated adults, larvae are adult forms from foreign taxa that were transferred by hybridization. Larval transfer offers explanations for many anomalies in animal development, as shown here by examples from insects and echinoderms. Hybridogenesis (evolution by hybridization) and symbiogenesis (evolution by symbiosis) are examples of evolution by merger of lineages. They are quite distinct from Darwinian ‘descent with modification’, which is evolution within separate lineages.
<Larval transfer is a theory that explains the origins of larvae. It is not a new theory, but as yet, it has been accepted by only a minority of biologists. Larvae are young animals that differ significantly from adults, and they must metamorphose, or change form, to develop into other types of larvae or into adults. Larvae and adults are different phases in the development of some animals.
There are two main theories on the origins of larvae. The first, which I shall refer to as the common ancestor theory, assumes that larvae are young animals that gradually became more and more different from corresponding adults. Under this theory, the larva and the adult evolved from a common ancestor, and they are products of a single genome. The second theory, which I will refer to as the larval transfer theory, proposes that larvae were later additions to life histories, and they originated as adults of distantly related animals. Different species occasionally hybridized to produce new animals, which hatched in a form resembling one parent and then metamorphosed into a form resembling the other. Each hybrid was an animal with larvae, in which the larval form had been transferred from another animal. Under this theory, the larva and the adult each have their own genomes, and metamorphosis is the change-over from the expression of one genome to the expression of the other.
Charles Darwin adopted the common ancestor theory without question [1]. He was convinced that all living things had evolved and that the main method of evolution was by “descent with modification and natural selection”. He assumed that the differences between larvae and adults had evolved gradually, and he thought that larvae showed the true relationships of animals, and were, therefore, very important in classification. He said, for example, that you could tell that a barnacle is a crustacean because it has crustacean larvae. Ernst Haeckel, in a book dedicated to Darwin, carried the common ancestor theory further by proposing that larvae represent ancestral adults [2]. He claimed that ontogeny (development) is a short and rapid recapitulation of phylogeny (evolutionary history), and, although the recapitulation theory has been amended several times, it is still very much alive today. Walter Garstang’s amendment, which proposes that modern larvae represent ancestral larvae rather than ancestral adults, is widely accepted [3].
Frank Balfour was a committed Darwinian, but he disagreed with Darwin on the origins of larvae. He proposed, in his Treatise on Comparative Embryology, that virtually all larvae are ‘secondary’ and have been ‘introduced into the ontogeny of species’ [4]. Balfour was regarded by his contemporaries as one of the greatest biologists of his day and Charles Darwin’s successor. He died on 19 July 1882, aged 30, attempting to climb Mont Blanc, the highest peak in Western Europe. Darwin had died three months previously, at the age of 72.
I had read parts of Balfour’s Treatise, but, until 2000, I had not read the part in which he suggests that virtually all larvae are ‘secondary’ and had been ‘introduced’ into life histories. The two volumes of the Treatise are respectively labelled ‘Invertebrata’ and ‘Vertebrata’, and his general remarks on larvae are in a separate section at the end of the vertebrate volume. For most of my working life, I followed Darwin in assuming that larvae and adults had evolved from common ancestors. Over the years, however, I became aware of more and more examples that did not fit this theory, and I eventually produced my own theory. This claims that larvae were later additions to life histories. The basic forms of all larvae originated as adults in other animal taxa (groups used in classification), and they were transferred by hybridization. The first larvae resulted when eggs of one species were fertilized by sperm of another species, resulting in a ‘sequential chimera’ [5]. The original chimera was a fabulous monster with parts of several animals. My sequential chimera is a hybrid in which the forms of the respective parents are expressed in succession. All descendants of this hybrid were animals with larvae. My early publications on the subject apply larval transfer to animals in eight phyla, but not all these larvae are traced back to unrelated adults [5-7]. My later publications claim that all larvae originated as remotely related adults [8,9]. My notion of larval transfer is virtually the same as Balfour’s concept of secondary larvae that have been introduced into life histories, but I go further than Balfour in suggesting the sources of larvae and the means of transfer. He and I also agree with each other, but disagree with Haeckel, on why radially symmetrical adult echinoderms have bilaterally symmetrical larvae (see later). I agree with Haeckel that modern larvae represent former adults, but, while Haeckel thought these were ancestral adults, I claim instead that they are only remotely related to the adults that will succeed them in ontogeny. I insist that larvae do not recapitulate the phylogeny of adults. There are dozens of types of larvae, but here I shall concentrate on the larvae of insects and echinoderms.
Caterpillars are well known as the larvae of butterflies and moths, which are lepidopterans, but sawflies (which are hymenopterans) and scorpionflies (which are mecopterans) also have caterpillar larvae. Caterpillars grow in a series of moults, separated by intermoults, instars or stages. Caterpillars crawl, have no wings, and usually have three pairs of jointed thoracic legs and a variable number of unjointed abdominal prolegs. The adults that develop later do not moult or grow. They can fly or settle, have two pairs of wings, three pairs of jointed thoracic legs and no abdominal legs or prolegs. Metamorphosis from a caterpillar to an adult insect takes place in an inactive pupa or chrysalis (Figure 1).
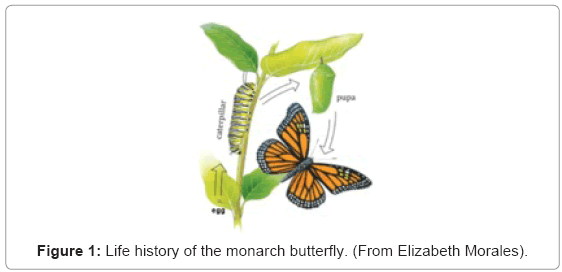
Figure 1: Life history of the monarch butterfly. (From Elizabeth Morales).
The organs and tissues of the caterpillar break down in the pupa, and they are replaced by a soup of liquid, stem cells and ‘imaginal discs’. These are small discs that form during the last caterpillar stage, and the cuticle, legs, wings and nerves of the imago (adult) develop from them. All the other adult organs, including the heart, gut and digestive gland, grow from stem cells in the pupal soup. No part of the larva becomes a part of the adult. The developing animal dismantles the larva then starts again to produce the adult, and I question whether this complex procedure could have evolved by a series of small changes, each subject to natural selection. In contrast, dragonflies and damselflies (Figure 2A) manage the transition from larva to adult without it.

Figure 2: A, adult damselfly. B, larval damselfly. C, adult thysanuran. (From Google Images).
Of course, the larva of a dragonfly or damselfly (called a nymph) (Figure 2B) is not a caterpillar. It is aquatic, and it resembles an adult thysanuran (three-pronged bristletail) (Figure 2C), a so-called wingless insect that has no larva. I claim that an ancestor of dragonflies and damselflies acquired larvae by hybridization with a thysanuran. The dragonfly or damselfly nymph with its thysanuran counterpart is not an isolated case. This gives an important clue to the origins of larvae. All larvae have, or had, adult counterparts with no larvae. The adult counterpart of a caterpillar is an onychophoran or velvet worm [10]. Caterpillars of most butterflies, moths, sawflies and scorpionflies have three pairs of jointed thoracic legs and a variable number of unjointed abdominal prolegs (Figure 3A). A velvet worm looks like a caterpillar, but all its trunk appendages are unjointed prolegs, and there is no clear distinction between thorax and abdomen (Figure 3B). The larva of the primitive moth Micropterix, however, resembles a velvet worm: all its legs are unjointed prolegs, and there is no clear distinction between thorax and abdomen (Figure3C). I claim that Micropterix has retained the original larval form of insects that pupate, and jointed thoracic legs and the loss of both types of legs in some insect larvae evolved later. I suggest that an early onychophoran, not long (on the evolutionary timescale) after it had emerged from the sea, hybridized with an ancestor of pupating insects, which had not yet acquired larvae or pupae. This hybrid was the first insect with caterpillar-like larvae. It had two genomes, an onychophoran genome and an insect genome, which were integrated within one organism. The onychophoran genome was expressed first as the larval caterpillar. The onychophoran tissues and organs were too different from insect tissues and organs to permit smooth metamorphosis, but it devised the complicated but effective pupal method of ‘start-again metamorphosis’. This hybrid survived, but there were probably many other hybrids that failed to produce pupae and perished. An ancestor of dragonflies and damselflies acquired larvae from a thysanuran, also by hybridization, but in this case there was less difference between the animals that hybridized, and ‘start-again metamorphosis’ was unnecessary. Imaginal discs were mentioned earlier as the progenitors of most outer structures of insects that pupate. These discs form during the last caterpillar stage, but imaginal cells, the precursors of imaginal discs, occur in all caterpillar stages. In all caterpillars except those about to pupate, the caterpillar’s immune system treats the imaginal cells as alien and destroys them. Only in the last caterpillar stage, when the caterpillar’s immune system breaks down, do the imaginal cells survive and aggregate into discs [11]. This scenario is readily explained in terms of the larval transfer theory, which postulates that caterpillars are derived from an onychophoran genome. The caterpillar’s immune system is, therefore, derived from an onychophoran immune system, and it regards imaginal insect cells as alien and destroys them. Under the common ancestor theory, it is difficult to explain how and under what selection pressure the ancestral immune system should have evolved into two systems, one of which destroys cells crucial to metamorphosis. I have suggested experiments and genetic investigations to test my claim that caterpillars are transferred onychphorans [10].

Figure 3: Caterpillar larvae and an adult onychophoran. A, Luna moth caterpillar with 5 prolegs including anal clasper. B, adult onychophoran, Peripatus. C, anterior end of caterpillar larva of archaic moth Micropterix. (From: A, Wikipedia proleg; B, Google Images; C, SEM by Donald R Davis, SI, NMNH, USA).
Most caterpillars have fewer abdominal prolegs than onychophorans, and I suggest that grub-like larvae evolved by the loss of all abdominal appendages and reduction of all legs produced maggots. A wide range of insect larvae can thus be derived from the presumed onychophoran-like ancestor. I claim that the larvae of dragonflies, damselflies and mayflies are transferred thysanurans (three-pronged bristletails), and other insect larvae had their origins in diplurans (two-pronged bristletails). Dipluran larvae, also known as campodeiform larvae, occur in several families of beetles (coleopterans), in some lacewings (neuropterans), and in some caddis-flies (trichopterans). The distribution of dipluran larvae bears no relation to the classification of adult insects. This is unexplained in terms of common ancestry, but it is readily explicable in terms of larval transfer by chance hybridizations.
The American meloid beetle, Epicauta vittata, goes through three larval and two pupal phases between egg and adult, a process known as hypermetamorphosis (Figure 4). The egg hatches as a dipluran larva (known as a triungulin), which metamorphoses into a caraboid larva (resembling the larva of a carab beetle), which is followed by an inactive pseudopupa (or coarctate larva), which is succeeded by a scarabaeoid larva (resembling that of a scarab beetle), which pupates, and the pupa transforms into the adult beetle. This complex life history seems to defy explanation in terms of the gradual accumulation of small mutations demanded by the common ancestor theory. If larvae show the true relationships of animals, as Darwin claimed, Epicauta appears to be related to diplurans, caraboid beetles and scaraboid beetles. Its bizarre life history, however, is explicable in terms of larval transfer. I suggest that different ancestors of Epicauta hybridized with members of each of these taxa, and the respective types of larvae were successive additions to the life history of the evolving beetle. The genetics of Epicauta have not yet been studied in sufficient detail to show these inferred transfers.
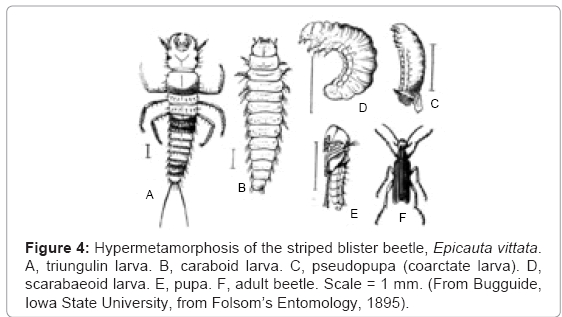
Figure 4: Hypermetamorphosis of the striped blister beetle, Epicauta vittata. A, triungulin larva. B, caraboid larva. C, pseudopupa (coarctate larva). D, scarabaeoid larva. E, pupa. F, adult beetle. Scale = 1 mm. (From Bugguide, Iowa State University, from Folsom’s Entomology, 1895).
The foregoing examples demonstrate that the larval transfer theory offers explanations of facts in the development of insects, including some that are otherwise unexplained. Larval transfer, however, applies to all animals with larvae. For example, ‘start-again metamorphosis’ and a form of pupation also occur in bryozoans (moss animals), and cases of more than one type of larva in a life history occur in several orders of insects, some crustaceans and some echinoderms [7].
The Echinodermata is the phylum that includes starfish and sea-urchins, which are radially symmetrical animals, with no left and right sides. Larval echinoderms, however, are bilaterally symmetrical, with left and right sides. Haeckel ingeniously suggested that original echinoderms were bilateral and that echinoderm larvae have retained this type of symmetry while the adults evolved radial symmetry in response to a sedentary way of life. The evidence, however, does not support Haeckel’s view, but it is consistent with the conclusions of Balfour and myself that early echinoderms were radial and without larvae [4-6]. They eventually acquired bilateral larvae by transfer, but not until after the phylum had diverged into sea-lilies, brittle-stars, starfish, sea-urchins, sea-cucumbers, and fourteen other classes of extinct echinoderms. The order in which the classes acquired larvae bears no relationship to the order in which they evolved. This explains why the affinities of echinoderm larvae are quite different from those of adult echinoderms. Some echinoderms with no planktonic larvae go through a brief bilateral phase, which suggests that they are descended from forms with larvae. However, a different form of direct development occurs in a few brittlestars, in brooding heart-urchins of the genus Abatus, and in the only known genus of sea-daisies, Xyloplax [7,12]. These echinoderms have no trace of a bilateral phase in their development, and the formation of the mouth and coelom is quite different from that in echinoderm larvae. This is consistent with the view that these species are not descended from forms with larvae, and they have retained the original method of echinoderm development. Genes of some of these species are currently being studied. In all echinoderms with larvae, the juvenile echinoderm, radial from the start, begins to grow from stem cells within the bilateral larva, and it eventually migrates to the outside of the larva (Figure 5A). Most echinoderms go through a stage like this, in which the juvenile can move its arms (if any) and tube-feet quite independently of the swimming movements of the larva. Indirect development in echinoderms involves ‘overlapping metamorphosis’, in which the larva and juvenile develop side-by-side. This also occurs in some polychaete worms, some nemertean worms, and some urochordates [7]. The independence of the juvenile and larva is carried further in the starfish Luidia sarsi, in which the juvenile drops off the swimming larva (Figure 5B) [11]. The larva can continue swimming for three more months, and during this time the bilateral larva and the radial juvenile, which developed from the same egg, may be miles apart. This remarkable independence of larva and juvenile is unexplained in terms of the single genome permitted by common ancestry, but it is consistent with the plural genomes required by larval transfer. Acorn-worms, also known as tongue-worms or enteropneusts, are a class of bilaterally symmetrical hemichordates, each with a tripartite body and gill slits (Figure 6A). I have proposed that acorn-worm larvae and most echinoderm larvae were transferred, directly or indirectly, from an ancestor of Planctosphaera pelagica (Figure 6B). This species, which can attain a diameter of 25 mm [13], resembles a giant larva of an acorn-worm, but even the largest specimen shows no sign of metamorphosis, and I am convinced that it is an adult. The close affinities between acorn-worm larvae and echinoderm larvae have been accepted since the late 19th century [14]. On the other hand, the smooth metamorphosis in acorn-worms is strikingly different from the overlapping metamorphosis in echinoderms, and I question whether these two methods could have evolved from an ancestral method, as the common ancestor theory implies. Under larval transfer, however, larvae were later additions to life histories, and each animal that acquired larvae had to devise its own method of metamorphosis. The 18S ribosomal gene has been frequently used to produce ‘molecular phylogenies’ of animals. In the case of acorn-worms and echinoderms, however, the resulting phylogenetic tree does not show the order in which they evolved (Figure 7). The tree implies that radial echinoderms evolved from bilateral ancestors of acorn-worms, as Haeckel said. On the other hand, sea-cucumbers were the last of the extant classes of echinoderms to appear in the fossil record, not the first as indicated in Figure 7. In addition, the anatomy of the relevant echinoderms makes it highly unlikely that ancestral sea-cucumbers gave rise to starfish and sea-urchins. Larval transfer, however, offers a solution to this anomaly. The 18S tree does not reflect the order in which adult acorn-worms and echinoderms evolved, but it does reflect the order in which (I claim) they acquired larvae [8,13]. 18S ribosomal RNA does not carry recipes for larvae, but it appears that the acts of hybridization which transferred DNA recipes for larvae to acorn-worms and echinoderms also transferred the 18S gene. In this case at least, genetic phylogeny is in accord with larval transfer. True evolutionary histories must take account of extant and fossil adults, and must include evaluation of genetic evidence that is not restricted to one gene. Larvae must not be ignored, and the taxonomist must bear in mind that larval phases and the genes prescribing them were added through hybridization during the evolution of animals with larvae. Both adults and larvae evolve, but not necessarily at the same rate. Hybridization unavoidably complicates biological classification, but “nature ….. does not consider the convenience of taxonomists” [8].
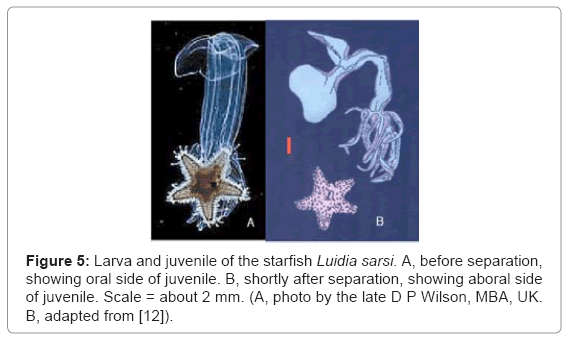
Figure 5: Larva and juvenile of the starfish Luidia sarsi. A, before separation, showing oral side of juvenile. B, shortly after separation, showing aboral side of juvenile. Scale = about 2 mm. (A, photo by the late D P Wilson, MBA, UK. B, adapted from [12]).

Figure 6: A, the acorn-worm, Dolichoglossus. Scale = about 2 cm. B, Planctosphaera pelagica, from the Gulf Stream. Scale = about 5 mm. (A from [6]. B, photo by LP Madin, WHOI, USA).
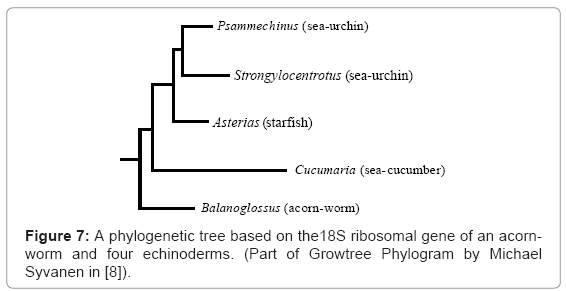
Figure 7: A phylogenetic tree based on the18S ribosomal gene of an acornworm and four echinoderms. (Part of Growtree Phylogram by Michael Syvanen in [8]).
Larval transfer is an example of evolution by hybridization, or hybridogenesis. Another example of hybridogenesis is component transfer, which accounts for the occurrence of such organs as lophophores, the feeding apparatus of brachiopods, bryozoans, entoprocts, phoronidans and pterobranch hemichordates [12]. Component transfer was a vital factor in the Cambrian explosion, which was a relatively rapid increase in numbers, size and complexity of animals about 550 million years ago. Hybridogenesis involves the merger of two or more genomes, which evolved in separate lineages. Genomes also merge in symbiogenesis or evolution by symbiosis, which explains the origin of eukaryotic cells (of plants, animals and fungi) from bacteria and archaea. An English translation is now available of the 1924 book on symbiogenesis by the Russian botanist Boris Kozo-Polyansky [15]. Independently of Kozo-Polyansky, and some decades later, Lynn Margulis reached similar conclusions on the origin of eukaryotic cells by symbiogenesis [16-18]. Hybridogenesis and symbiogenesis are quite distinct from Darwinian ‘descent with modification’ [1], which is evolution within separate lineages. Organisms have evolved by all three methods, and they may be subject to natural selection however they evolved.
I am very grateful to Michael Syvanen for the Growtree Phylogram, part of which is reproduced in Figure 7 and to Farley Fleming for constructive comments and suggestions.