Journal of Horticulture
Open Access
ISSN: 2376-0354
ISSN: 2376-0354
Research Article - (2015) Volume 2, Issue 3
The presence of leaves in citrus inhibit axillary bud break which were released, in several citrus cultivars, as soon as leaves were removed. Since even the presence of small petiole alone was as effective in inhibiting bud growth in Satsuma trees as the whole leaf, this indicates that the supply of sugars or photosynthesis play little role in regulating bud break in Owari Satsuma trees. It appears that physical injury through complete defoliation may trigger hormonal stimulation that in turn initiates the metabolic processes leading to bud break. Majority of bud break from defoliation developed vegetative growth, but, as anticipated, chilling treatment in defoliated plants resulted in majority of buds breaking into flowers. Since differentiation of buds into vegetative or reproductive structures occur in the absence of leaves it indicates that in grapefruit, and possibly other citrus cultivars, metabolic processes leading to flowering occur within the resting bud tissue. Changes in the levels of specific polyphenols occur when buds begin to sprout as vegetative shoot, marked by increase in chlorogenic acid and naringenin levels. When buds develop into reproductive structures then such developmental change accompany dramatic rise in the levels of hesperidin, naringenin, and Apigenin-7-glucoside. In general, the results clearly show a strong inhibition of axillary bud sprouting even with the presence of a small portion of the leaf. Very distinct changes in the levels of specific polyphenols were associated with bud sprouting to vegetative shoot and when it develops into flower.
<Keywords: Citrus greening; Defoliation; Flowering; Flushing; Huanglongbing; Polyphenols
The Huangalongbing (HLB), also called citrus greening, is a lethal disease that causes enormous economic losses in Florida and other citrus growing areas of the world [1,2]. This disease is spreading to other southern states in the US and was detected in Texas in 2012 [3]. The disease causing bacteria (Candidatus Liberibacter spp.) spreads through an insect vector Asian citrus psyllid (ACP); Diaphorina citri Kuwayama. The ACP feeds on phloem sap of leaves of different ages but it primarily colonizes young emerging buds of new flush where it oviposit and the nymphs inhabit [2]. At present, there is no cure for the HLB, and therefore, the spread of the disease is primarily managed through controlling psyllid population [4,5]. Thus, to find effective and efficient ways to control psyllid population one needs to study interactions between psyllids and the new flush for which a continuous supply of new citrus flush is needed for researchers. In addition, understanding and managing flushing in field grown citrus trees will provide valuable tool for efficient and effective management of psyllid populations in the field by appropriate timing of spray applications of insecticides.
Bud break and dormancy has been extensively studied in various plants and it is well known that several environmental factors (e.g., day-length, temperature, water stress, nutrients) play important role in controlling bud break [6-14]. In addition, it has long been known that those correlative factors such as apical dominance play important role in controlling axillary bud break [15]. For example, removal of apex in dicots triggers sprouting of axillary buds, and hence, tip pruning in macadamia trees has been shown to produce new flush [16]. The effects of defoliation at various levels in different cultivars have given mixed results ranging from no effect to promotion of bud sprouting [17].
For obvious economic reasons majority of studies on bud break in citrus are related to flowering and are thoroughly discussed in various review articles [18-20]. Only a few old studies describe environmental regulation of new flushes [21,22]. Limited studies on the effect of rootstock on bud break have also been conducted [23,24]. However, predicting and management of flushing in citrus is crucial for developing efficient and effective methods to control citrus greening disease. In this study, experiments are described that promote flushing in citrus through defoliation and temperature changes. In addition, changes in the levels of polyphenols which have been implicated in bud break of different plants species [25-28] were also investigated to determine if they can be used as markers. Therefore, in this study we conducted a series of experiments to induce flushing in citrus under controlled conditions throughout the year and to explore biochemical markers for bud break that could be used to time flushing in the field.
Defoliation experiments
Two-year old citrus cultivars (Rio-Red grapefruit, Owari Satsuma, Mars orange all on sour orange rootstocks, and Kaffir limes seedlings) were purchased from a local nursery and grown in 16-liter pots filled with a mixture of peat: perlite: vermiculite (1:1:1 by wt.), supplied with nutrients, as described previously [29]. The trees were grown in a nursery block under 30% shade during summer and then in the greenhouse maintained at 29 ± 2°C from November till end of February at the USDA-ARS facility in Weslaco, Texas. By keeping trees under warm conditions during winter there was minimal bud sprouting in spring (late February-March). Defoliation experiments were conducted in May-June and after defoliation the trees were placed back in greenhouse maintained at 30 ± 2°C. The defoliation was conducted by removing the leaf from the base joining stem except in one experiment where different portions of leaves were removed (Figures 1-3). The numbers of buds sprouted after defoliation was counted 10-13 days after defoliation. At least six replicate trees were used for each treatment.
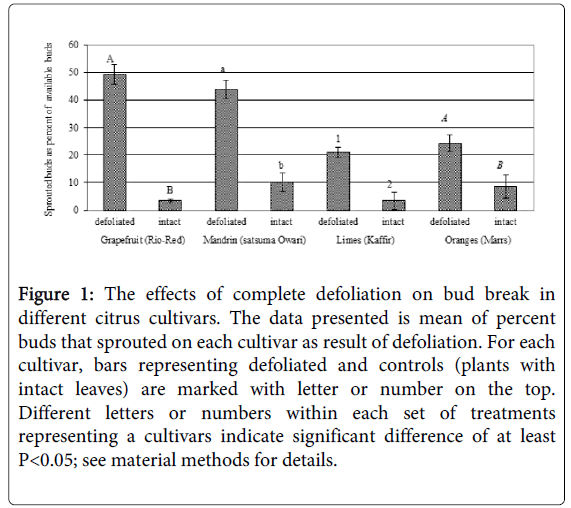
Figure 1: The effects of complete defoliation on bud break in different citrus cultivars. The data presented is mean of percent buds that sprouted on each cultivar as result of defoliation. For each cultivar, bars representing defoliated and controls (plants with intact leaves) are marked with letter or number on the top. Different letters or numbers within each set of treatments representing a cultivars indicate significant difference of at least P<0.05; see material methods for details.

Figure 2: Pictures of representative of Rio Red grapefruit plants at the start of experiment (A), immediately after defoliation (B), one week after defoliation (C), and development of foliage three weeks after defoliation (D).
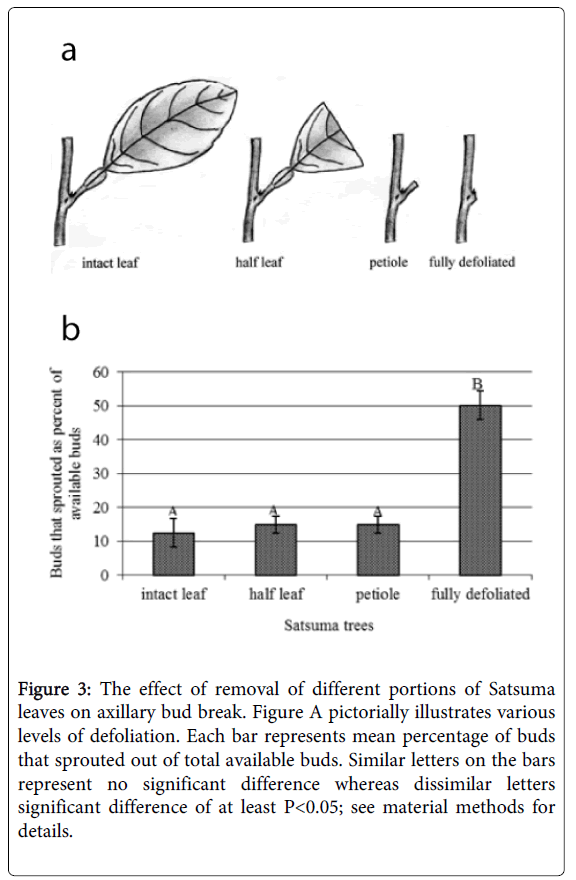
Figure 3: The effect of removal of different portions of Satsuma leaves on axillary bud break. Figure A pictorially illustrates various levels of defoliation. Each bar represents mean percentage of buds that sprouted out of total available buds. Similar letters on the bars represent no significant difference whereas dissimilar letters significant difference of at least P<0.05; see material methods for details.
In the first defoliation experiment the effect of complete removal of leaves on bud break in various citrus cultivars was determined as shown in in (Figures 1 and 2). In a second defoliation experiment the effect of complete and partial removal of leaves as shown in (Figure 3A and 3B) was tested on axillary buds. Again six replicate plants were used for each treatment in each experiment.
Defoliation and chilling experiment
For chilling treatment, trees were placed in an indigenously designed and constructed plant growth chamber for 8 weeks [30]. The daily temperature changes in the growth chamber were as follows: (0:00-5:00 time)=2.8°C; (5:00-8:00 time)=7.2°C; (8:00-10:00 time)=(12.8°C; (10:00-18:00 time)=19.4°C; (18:00-19:00 time)=15.6°C; (19:00-20:00 time)=12.8°C; (20:00-21:00 time)=7.2°C; (21:00-24:00 time)=2.8°C. The temperature regime and the period of incubation were selected based on preliminary tests that showed these to be optimal conditions for induction of bud break. The buds breaks occurred a week after the trees were transferred to greenhouse maintained at 30 ± 2°C. Six replicate Rio-Red trees were defoliated and immediately transferred to the chilling growth chamber for 8 weeks, thereafter, the number of buds that sprouted into vegetative shoot or flowers were counted after 10-13 days. For non-chilled controls, six defoliated plants were kept in the greenhouse and the sprouted buds were counted after 10-13 days.
Separate chilling experiments were conducted to study changes in polyphenol levels in buds. For this purpose, six replicate Rio red trees were subjected to defoliation but kept under greenhouse conditions (maintained at 30 ± 2°C), and additional six trees were defoliated and the given chilling treatment described above. The lateral axillary buds were collected 4-5 days after the defoliated plants that were given 8 week chilling treatment were brought to warm greenhouse, and also from control plants that were defoliated but kept in greenhouse; i.e., without chilling treatment.
Extraction and analyses of polyphenols
Plant samples were taken four days after defoliation when the buds were just beginning to sprout. Several buds with small bark (approximately 3-4 mm) were collected from six replicate trees of defoliated and six replicate tree of non-defoliated control trees and stored at -80°C until needed for analyses. Plant samples were pulverized in liquid nitrogen using our previously published method [28]. Three replicate sample of pulverized plant material were extracted in 80% methanol. A 60 mg aliquot of frozen pulverized plant material was homogenized in 2 ml of 80% methanol with a polytron (Brinkman model PT 3100) at maximum speed for 30 second three times with 10-15 second intervals followed by 10 seconds of sonication with a Branson 4C15 probe at high power (Branson 150I, Fisher Scientific, Danbury, CT). The mixture was centrifuged for 30 min at 20,000 rpm at 20°C using an Eppendorf (5417R, Westbury, NY) refrigerated centrifuge. After centrifugation the supernatant was decanted into a separate 2 mL microfuge tube and the pellet was re-extracted two more time with 500 µL of 80% methanol as described above. The supernatant liquids from three extracts were pooled and the pooled extract was dried with speedvac (SPD1010, Thermo Savant, Holbrook, NY). The dried samples were reconstituted in 500 µL of 80% methanol, and filtered through a Costar® Spin-X® filter (0.2 µm). The filtrate was stored at -80°C until used for analysis by HPLC. Triplicate sample of leaves were extracted for each treatment and each leaf sample was extracted in triplicate and each extract was chromatographed on HPLC in triplicates.
Separation of phenolic compounds present in methanolic extracts, as described above, was performed using a Waters Xterra MS C18 (5 µm particle size) (3.9 mm x 150 m) maintained at 35°C and eluted at 1 mL/min as described previously [31]. The column was fitted in Waters Alliance 2695 HPLC system equipped with a 2996 photo-diode array detector. Phenolic compounds were identified by matching retention times (tr) and the UV spectra in the extract with the peak and the spectra of known standard compounds. For quantitative measurement of different polyphenols in the extracts, regression curves were developed using known standard compounds. All experiments were performed in triplicate, and for HPLC analysis, each sample was analyzed by triplicate injections.
Data analysis
Statistical analysis on the means of triplicate experiments were performed using the ANOVA procedure of the InStat® software, version 3.0 (GraphPad, San Diego, CA). The Tukey’s test was used for assessing significance. Significance was reported p <0.05; i.e., including p values that may be even <0.01. To show whether or not the results of two treatment significantly different, letters or numbers are added on top of each bar (representing a specific set of treatments) where same letter indicate no significant difference but dissimilar letters indicate significant difference at “at least p <0.05”.
Defoliation experiments
Complete removal of leaves significantly increased bud break in several cultivars [including Rio Red grapefruit, Owari Satsuma mandarin, Kaffir lime, and Mars oranges when compared to none defoliated control plants without any chilling or temperature change (Figure 1). However, percentage of bud break was much more pronounced in Rio Red grapefruit and Owari Satsuma (cv) trees (Figure 1). Figure 2 shows Rio red plant at the time of defoliation (A), immediately after defoliation (B), and abundance of new flush (that be used to study interactions with ACP) a week after defoliation (C), and production of foliage (D) three weeks after defoliation.
In the subsequent experiment, various portions of the leaves were removed from Satsuma trees to study how much portion of the leaf was necessary to suppress bud break (Figure 3A). Partial defoliation was ineffective in promoting bud break; i.e., even when a small petiole was left intact it completely inhibited bud break in Satsuma cultivar (Figure 3B).
In fact, there was no significant difference in the percentage of buds breaks (buds that sprouted as percent of available buds) between treatments where the whole leaves was kept intact and in the treatment where the entire lamella was removed leaving only a stump of petiole intact (Figure 3A).
Defoliation plus chilling experiment
Since it known for a number of plant species that chilling treatment can promote bud break, additional experiments were conducted to study the effect of chilling treatment on bud break in in defoliated Rio Red grapefruit.
Chilling treatment as described in material and methods was given for 8 weeks before transferring the plant back to greenhouse to initiate bud break. Total number of buds that sprouted with defoliation plus chilling (78% of the available nodes) were significantly higher (p<0.05) than the buds that sprouted with defoliation alone (67%) (Figure 4). While all the buds that sprouted by simple defoliation in the greenhouse were vegetative (67% vegetative and 0% flowering), majority of buds in defoliated plants that were given 8 weeks of chilling period sprouted into flowers (47% of the available buds) while significantly less sprouts turned into vegetative shoots (31% of available buds) (Figure 4).
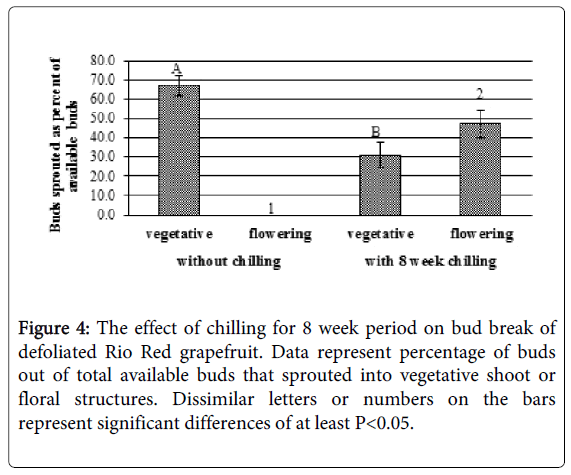
Figure 4: The effect of chilling for 8 week period on bud break of defoliated Rio Red grapefruit. Data represent percentage of buds out of total available buds that sprouted into vegetative shoot or floral structures. Dissimilar letters or numbers on the bars represent significant differences of at least P<0.05.
Changes in polyphenol levels during vegetative bud break after defoliation
Levels of chlorogenic acid and naringenin significantly (p<0.05) increased (28% and 20%, respectively) during bud break when Rio Red trees were defoliated that normally produce vegetative shoot (Figure 5B). There were no significant changes in the levels of hesperidin and apigenin-7-glucoside at the early stages of bud break in Rio Red tress from the defoliation treatment (Figure 5A). The levels of naringin and naringin DHC, however, decreased (21% and 28%, respectively) with the vegetative bud break from defoliation in Rio Red trees (Figure 5A and 5B).
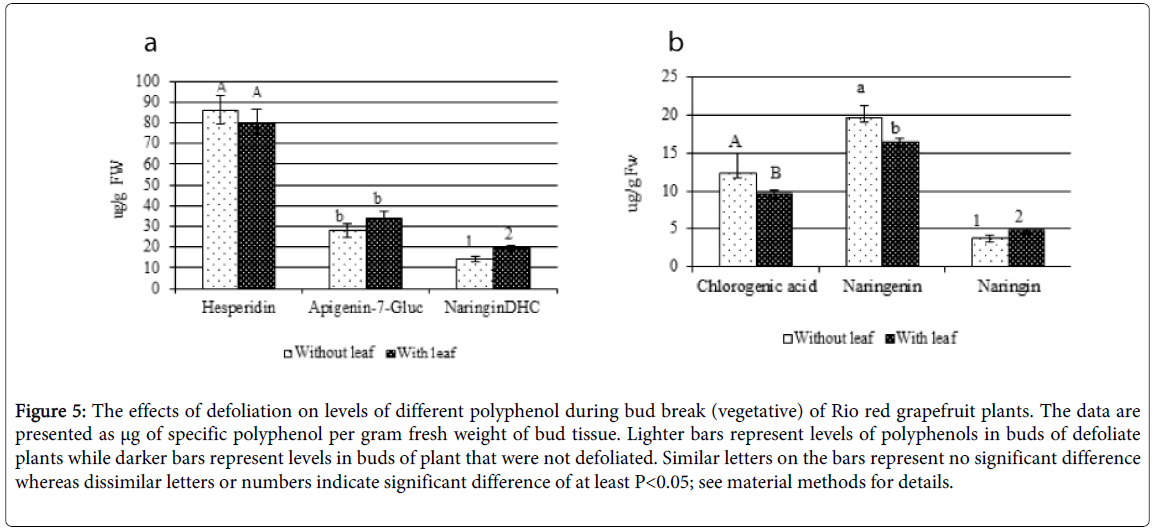
Figure 5: The effects of defoliation on levels of different polyphenol during bud break (vegetative) of Rio red grapefruit plants. The data are presented as μg of specific polyphenol per gram fresh weight of bud tissue. Lighter bars represent levels of polyphenols in buds of defoliate plants while darker bars represent levels in buds of plant that were not defoliated. Similar letters on the bars represent no significant difference whereas dissimilar letters or numbers indicate significant difference of at least P<0.05; see material methods for details.
Changes in polyphenol levels in bud break after defoliation and chilling and after defoliation at warm temperatures
Pronounced increases in levels of several polyphenols were observed in defoliated Rio-Red sprouting buds subjected to 8 weeks of chilling treatment compared to the buds that were induced to sprout by defoliation alone under warm temperatures in the greenhouse (Figure 6A and 6B). For example, levels of hesperidin, naringenin, and apigenin-7-glucoside considerably increased (by 246%, 190% and 150% respectively) in the sprouting buds of defoliated trees that received 8 weeks of chilling treatment compared to trees that were defoliated alone and kept at warm temperatures (Figure 6A and 6B). Chilling had no effect on the levels of naringin DHC, and chlorogenic acid (Figure 6A and 6B).
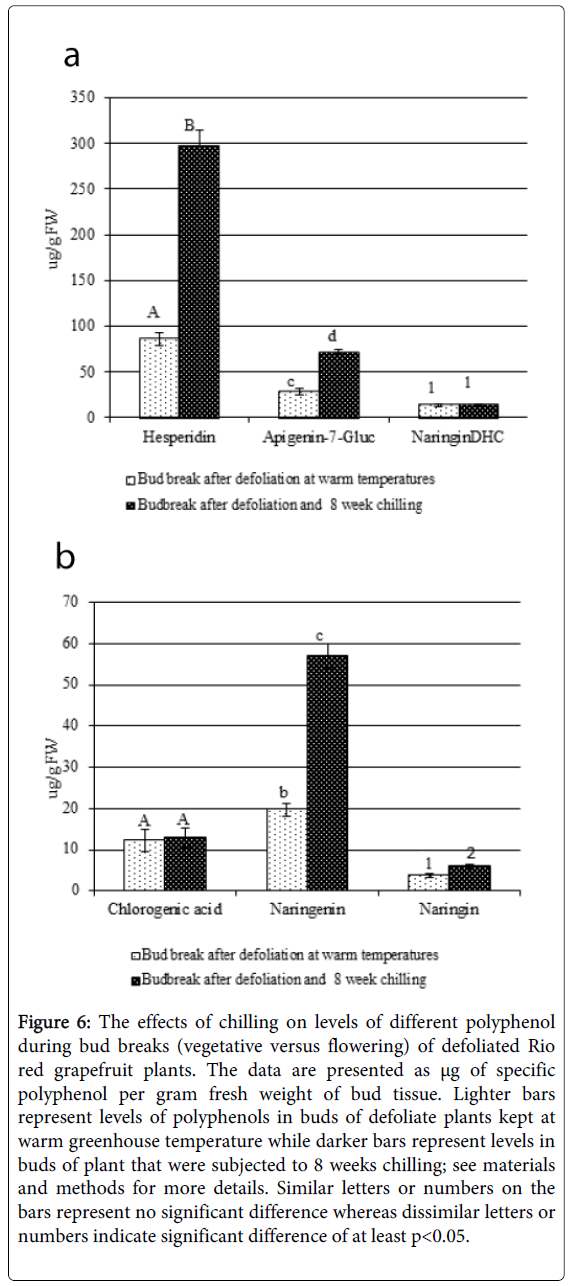
Figure 6: The effects of chilling on levels of different polyphenol during bud breaks (vegetative versus flowering) of defoliated Rio red grapefruit plants. The data are presented as μg of specific polyphenol per gram fresh weight of bud tissue. Lighter bars represent levels of polyphenols in buds of defoliate plants kept at warm greenhouse temperature while darker bars represent levels in buds of plant that were subjected to 8 weeks chilling; see materials and methods for more details. Similar letters or numbers on the bars represent no significant difference whereas dissimilar letters or numbers indicate significant difference of at least p<0.05.
Extensive and uniform new flushing can be induced in various citrus cultivars by simple removal of leaves; this is much more pronounced in Rio Red grapefruit and Owari Satsuma than others (Figures 1 and 2). Similar stimulatory effect of leaf removal and chemicals to induce bud break has been noted in other plant species such as peach, apple and grape plants [32]. In addition, it has been previously documented that removal of apical meristems induces lateral bud sprouting [33]. Thus, tip pruning has been used to induce lateral bud sprouting in macadamia but buds did not sprout uniformly [16]. Our results show that simple defoliation in citrus can be an effective method to induce flushing at will for research related to citrus greening disease and to investigate metabolic regulation of bud break. The ability to manipulate and predict bud break would be invaluable for efficient farm management practices such as proper scheduling of spray applications of insecticides to control ACP. The fact that even the presence of a petiole was as effective in suppressing lateral bud growth, just as the presence of the whole leaf inhibited bud break, it indicates that nutritional factors such as carbohydrates supply from leaves play little role in the bud break of citrus axillary buds.
Our findings are supported by reports that photosynthetic activity within the growing bud tissue could supply sugars needed for bud break in Valencia oranges [34]. In other species, however, supply of carbohydrates from leaves was considered important for bud break [35]. The dramatic difference between intact and removed petiole on bud break would relate to possible hormonal stimulus as result of wounding effect; and of course, production of hormonal stimuli in response to wounding have been reported in the literature [36-38]. Since the development of leaves after bud break occurs quite quickly (Figure 2), the plant soon becomes capable producing photosynthates for the plant needs even after complete defoliation; although complete defoliation of entire plant may not be necessary and only few branches may be defoliated to produce new flush.
While simple defoliation could effectively cause bud burst in citrus, chilling treatment of defoliated trees for 8 weeks further significantly (p<0.05) increased the percentage of bud breaks (Figure 4) and the observation is consistent with results obtained through winter defoliation in apples [39]. In addition, it is not surprising that 8 weeks of chilling resulted in the production of flowers from the bud burst since induction of flowering with various levels of chilling is well known phenomenon in many plant species including citrus [40-42].
The fact that chilling of defoliated trees shifted bud break from vegetative to reproductive bud growth indicates that metabolic changes within bud tissue from external stimuli are primarily responsible for flower induction in Rio Red grapefruit and this observation is similar to what we observed for flower induction in olives; i.e., developmental change in auxiliary buds could occur independent of leaf metabolism [43].
Levels of several polyphenols significantly changed when defoliation induced vegetative bud break; however, the changes were quite dramatic when chilling treatment was given to defoliated plants that resulted in reproductive bud growth (Figures 4-6). For example, there was no significant change in levels of hesperidin, Apigenin-7-glucoside, and only 20% increase of Naringenin during the sprouting of vegetative buds compared to non-sprouting control buds, while their levels increased by 246%, 150%, and 190% respectively in buds sprouting to produce flowers compared to buds sprouting to become vegetative shoot (Figures 5 and 6). Naringin also increased by 62% in buds sprouting to produce flowers, but in buds sprouting to become vegetative shoot (i.e., defoliation but no chilling) its levels actually decreased by 21%. Increases in poly phenols during reproductive bud break and development have been reported for different plant species [17,44]. The only polyphenols that actually increased significantly during the sprouting of vegetative buds were chlorogenic acid and naringenin, and out of these two only naringin continued to remain higher in flowering buds compared to vegetative buds; the chlorogenic acid levels remained unchanged in buds that turned into reproductive structures (Figures 5 and 6). Thus, increases in the levels of chlorogenic acid were distinctly related to bud break in Rio Red grapefruits that may be a good biological marker of vegetative growth if there was no accompanying dramatic increase in hesperidin, apigenin-7-glucoside, and naringenin. However, past studies have given contrasting result regarding correlations between chlorogenic acid levels and developmental changes such as vegetative versus reproductive growth. For example, higher levels of chlorogenic acid were found in leaves of flowering olive trees versus nonflowering tree but exogenous application of chlorogenic acid had no effect on floral differentiation [45-48]. High levels of chlorogenic acid were found in Lonicera japonica flowers, but in Asparagus officinalis, flowering was closely related to decreases in chlorogenic acid levels. In Chrysanthemum morifolium light quality that increased chlorogenic acid had no effect on induction of flowering, and application of chlorogenic acid had no response on dormant buds of peaches [49]. Exogenous applications of chlorogenic acid on peaches had no effect on bud break, but in Impatiens balsamina the application of chlorogenic acid alone and in combination with gibberellic acid promoted the initiation of floral buds [50,51]. These reports indicate that the role of chlorogenic acid could be different in different plant species. Our findings therefore, provide a valuable information regarding rise in the levels of chlorogenic alone (or in combination with naringenin) in dormant grapefruit buds as indicator of vegetative bud sprouting, and since it does not increase in flowering buds it is valuable pointer for flushing as opposed to flowering.
Phillips in 1962 [52], reported that Naringenin inhibit the bud breaking effect of gibberellin, and therefore, concluded that naringenin acts as modulator of bud break in peaches; but, Luna et al. [53] could not find any positive correlations between levels of naringenin and rest period in peach buds. However, Biggs [50] reported that exogenous application of naringenin had positive effect on bud break in peaches. Both increase and decrease in the levels of naringenin related to bud growth have been reported in different experimental conditions [22,54]. Apparently, studies with different plants under different experimental condition and sampling protocols are responsible for lack of consistency regarding correlations between naringenin levels and the bud break. Our experiments designed to capture changes in naringenin levels, along with other polyphenols, just at the time of bud break in grapefruit provide an accurate estimation of indicator molecules such as naringenin, hesperidin, and apigenin-7-glucoside that dramatically increase at the onset of flowering bud break. Thus, while rise in levels of chlorogenic acid, in combination with rise in Naringenin, indicate vegetative bud break in grape fruits, a dramatic rise in hesperidin, in combination with apigenin and naringenin indicate the onset of flowering bud break. This is valuable information that may be used to develop specific sensor for predicting bud break in grapefruit [55,56].
In general, this study clearly demonstrates that the presence of leaves in different citrus cultivars inhibit axillary bud break which were released soon after defoliation. Since even the presence of small petiole alone was as effective in inhibiting bud growth, just as the whole leaf, it indicates that supply of sugars or photosynthates from leaves play little role in regulating bud break in grapefruit. On the other hand, it appears that physical injury through complete defoliation may trigger hormonal stimulation that in turn starts the metabolic process leading to bud break. Majority of bud break from defoliation tend to develop vegetative buds, but, as anticipated, chilling treatment results in majority of bud breaks into flowers. Since differentiation of buds into vegetative or reproductive structures occurred in the absence of leaves (i.e. strictly by chilling defoliated plants induced flowering) it indicates that in grapefruit metabolic process leading to flowering occur within the resting bud tissue. Changes in the levels of specific polyphenols occur when buds begin to sprout as vegetative shoot, marked by increase in chlorogenic acid and naringenin levels, or develop into reproductive structures that accompany dramatic rise in hesperidin, naringenin, and Apigenin-7-Glucoside. Thus, pattern with which changes in levels of different polyphenols occur at the onset of bud break in grapefruit provide good indications for bud break into vegetative or reproductive structures.