Drug Designing: Open Access
Open Access
ISSN: 2169-0138
ISSN: 2169-0138
Research Article - (2014) Volume 3, Issue 3
Long neurotoxin 2 protein from the venom of Naja naja shows a great affinity towards acetylcholine receptors, which produces peripheral paralysis by blocking neuromuscular transmission at the postsynaptic site. In this study, we use a cascade SVM based TAPPred method which found 26 High affinity TAP Transporter peptide regions. TAP is heterodimeric transmembrane protein which transports antigenic peptide proteins to the endoplasmic reticulum where it binds to MHC class I molecules, These MHC-peptide complexes will be translocated on the surface of antigen presenting cells (APCs), where they are recognized by T-cell receptors to elicit an immune response. Therefore, prediction of TAP-binding peptides is helpful in identifying the MHC class I-restricted T-cell epitopes and hence helpful in the designing of synthetic peptide vaccine.
<Keywords: Binding affinity, Antigenic peptides, MHC-Binders, TAP binder, Cascade SVM
Naja najavenom generally contains large quantity of neurotoxins and cytotoxins [1,2]. Long neurotoxin 2 from N. naja shows a great affinity to the acetylcholine receptors. It binds to muscular and neuronal nicotinic acetylcholine receptor leading to ceases breathing, and produces peripheral paralysis [3,4]. Designing of synthetic peptide vaccine requires identification of antigenic regions of protein which are recognized by cytotoxic T lymphocyte (CTL) cells [5,6]. The recognition of such regions by CTLs involve breakdown of a protein [7,8]. into peptides by proteasome complex in cytosol after cleavage, the peptides are translocated by the transporter associated with antigen processing (TAP) into the endoplasmic reticulum (ER), where they bind to MHC class I molecules [9-12]. However, this MHC-peptide were translocated on to the surface of antigen presenting cells (APCs), where they are recognized by T-cell receptors (TCR) of CTLs to elicit an immune response [13]. TAP binders are important for understanding the processing and developing method of endogenous antigen to predict TAP binders [14-16]. N. naja neurotoxin peptide has potential for the development of synthetic peptide vaccine because even single epitope can generate ample immune response. This approach is based on the phenomenon of cross-protection, where an individual with mild toxic effects have strong immunity against severe effects of the same. Therefore, the identification and study of antigenic properties are important for the development of new vaccines [17]. However, this theme is implemented in designing synthetic peptide vaccines [18]. One of the important problems in subunit vaccine design is to search antigenic regions in an antigen that can stimulate T cells called Tcell epitopes [5].
Protein sequence analysis
The antigenic protein sequence of Long neurotoxin 2 from N. naja was retrieved from NCBI protein database [P25669]–to study the MHC class peptide binding, antigenicity, solvent accessible regions that allow potential drug targets to identify active sites.
Prediction of antigenicity
Antigenicity program predicts those segments from Long neurotoxin 2 protein, which are likely to be antigenic by eliciting an antibody response. In this research work antigenic epitopes of Long neurotoxin 2 are determined by using the Hopp and Woods, Welling, Parker, Kolaskar and Tongaonkar antigenicity methods [19-22].
Prediction of MHC binding peptide
MHC molecules are cell surface glycoprotein, which take active part in host immune reactions and involvement of MHC class-I in response to almost all antigens and the variable length of interacting peptides make the study of MHC Class molecules very interesting. The MHC peptide binding is predicted using neural networks trained on C terminals of known epitopes. RANKPEP predicts peptide binders to MHCI molecules from protein sequences or sequence alignments using Position Specific Scoring Matrices (PSSMs). MHC molecule binds to some of the peptide fragments generated after proteolytic cleavage of antigen [23,24].
Prediction of antigenic peptides by cascade SVM based TAPPred method
In the present study, we predicted cascade SVM based several TAP binders which was based on the sequence and the features of amino acids [25]. We found the MHCI binding regions (Table 2), the binding affinity of Long neurotoxin 2 having 71 amino acids, which shows 63 nonamers.
Solvent accessible regions
Solvent accessibility used to identify active site of functionally important residues in membrane proteins. For the study of solvent accessible regions in proteins, different measurement was performed that are useful for the prediction of antigenic activity, surface region of peptides. Emani et al. [26] predicts the highest probability that a given protein region lies on the surface of a protein and are used to identify antigenic determinants on the surface of proteins. Karplus and Schulz [27] predict backbone or chain flexibility on the basis of the known temperature B factors of the a-carbons. By using different scale we predict the hydrophobic and hydrophilic characteristics of amino acids that are rich in charged and polar residues [28-37].
A antigenic sequence is 71 residues long as-IRCFITPDITSKDCP NGHVCYTKTWCDGFCSSRGKRVDLGCAATCPTVRTGVDIQCCS TDDCDPFPTRKRP
Prediction of antigenic peptides
We found the antigenic determinants by finding the area of greatest local hydrophilicity. The Hopp-Woods scale was designed to predict the locations of antigenic determinants in a protein, assuming that the antigenic determinants would be exposed on the surface of the protein and thus would be located in hydrophilic regions (Figure 1). Welling antigenicity plot gives value as the log of the quotient between percentage in a sample of known antigenic regions and percentage in average proteins (Figure 2). We also study Hydrophobicity plot of HPLC/Parker (Figure 3), Kolaskar and Tongaonkar [38] antigenicity methods (Figure 4) and the predicted antigenic fragments can bind to MHC molecule is the first bottlenecks in vaccine design. In these methods we found the antigenic determinants by finding the area of greatest local hydrophilicity. Hydrophilicity is the analysis of the surface properties [39], membrane spanning regions [40], antigenic sites, exposed loops or buried residues of proteins and is used for understanding the interactions that occur between proteins and other macromolecules. The hydrophobicity scales display the distribution of polar and apolar residues along a protein sequence as well as help to predict membrane protein interactions as well as protein structure. Because many of the drugs interact strictly with membrane proteins, thus, for understanding protein interactions through measuring hydrophobicity could give great ideas for synthetic peptide vaccine [41-44].
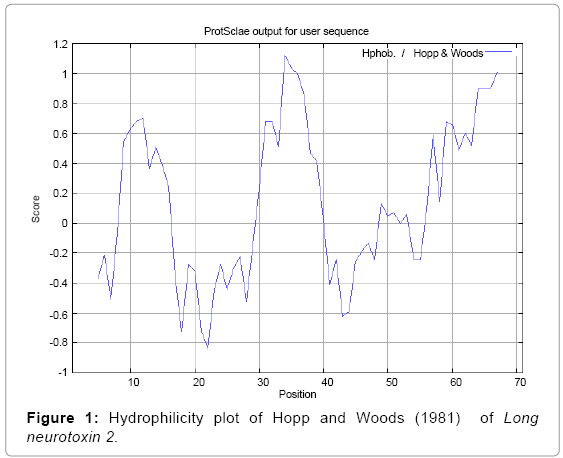
Figure 1: Hydrophilicity plot of Hopp and Woods (1981) of Long neurotoxin 2.
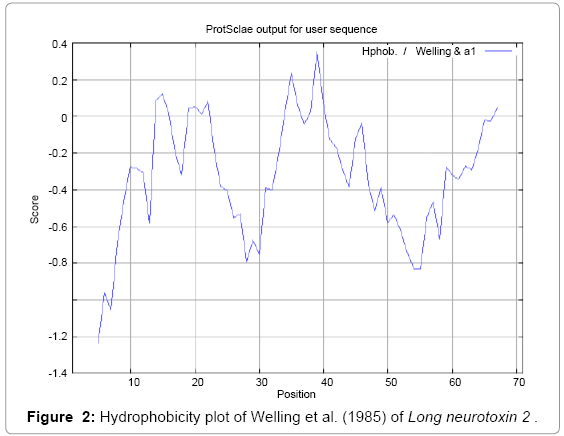
Figure 2: Hydrophobicity plot of Welling et al. (1985) of Long neurotoxin 2 .
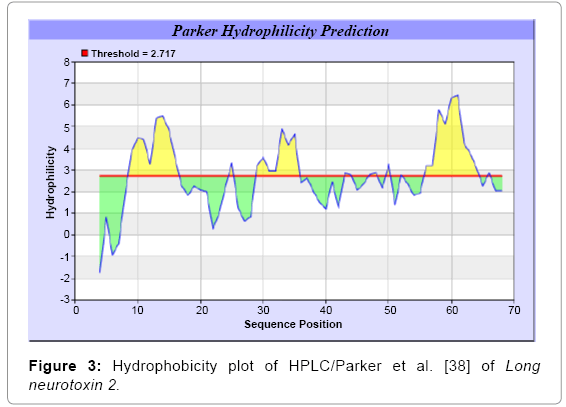
Figure 3: Hydrophobicity plot of HPLC/Parker et al. [38] of Long neurotoxin 2.
Figure 4: Kolaskar and Tongaonkar antigenicity are the sites of molecules that are recognized by antibodies of the immune system for the Long neurotoxin 2.
Prediction of MHC binding peptide
The prediction method was tested with Long neurotoxin 2 on a set of 4 different alleles i.e. H2-Db (mouse) 8mer, H2-Db (mouse) 9mer, H2-Db (mouse) 10mer, H2-Db (mouse) 11mer for MHC class I (Table 1). We also use a cascade SVM based TAPPred method which found 26 High affinity TAP Transporter peptide regions, which represents predicted TAP binders residues which occur at N and C termini from Long neurotoxin 2 (Table 2). Long neurotoxin 2 sequence is 71 residues long, having 63 nonamers. MHC molecules are cell surface proteins that participate in host immune reactions and involvement of MHC-I in response to almost all antigens. Predicted antigenic fragments can bind to MHC molecule is the first bottleneck in synthetic peptide vaccine design [41-44].
| MHC-I Allele | POS. | N | SEQUENCE | C | MW (Da) | SCORE | % OPT. |
|---|---|---|---|---|---|---|---|
| 8mer_H2_Db | 41 | DLG | CAATCPTV | RTG | 746.89 | 21.518 | 40.99 % |
| 8mer_H2_Db | 61 | STD | DCDPFPTR | KRP | 932.03 | 4.607 | 8.78 % |
| 8mer_H2_Db | 58 | QCC | STDDCDPF | PTR | 880.89 | 1.654 | 3.15 % |
| 8mer_H2_Db | 2 | I | RCFITPDI | TSK | 946.14 | 0.021 | 0.04 % |
| 9mer_H2_Db | 12 | ITS | KDCPNGHVC | YTK | 954.08 | 7.581 | 15.05 % |
| 9mer_H2_Db | 57 | IQC | CSTDDCDPF | PTR | 984.03 | 7.131 | 14.16 % |
| 9mer_H2_Db | 13 | TSK | DCPNGHVCY | TKT | 989.09 | 5.283 | 10.49 % |
| 9mer_H2_Db | 41 | DLG | CAATCPTVR | TGV | 903.08 | 3.327 | 6.61 % |
| 9mer_H2_Db | 40 | VDL | GCAATCPTV | RTG | 803.94 | 2.756 | 5.47 % |
| 10mer_H2_Db | 12 | ITS | KDCPNGHVCY | TKT | 1117.26 | 7.127 | 12.11 % |
| 10mer_H2_Db | 60 | CST | DDCDPFPTRK | RP | 1175.29 | 7.065 | 12.00 % |
| 10mer_H2_Db | 39 | RVD | LGCAATCPTV | RTG | 917.1 | 1.576 | 2.68 % |
| 11mer_H2_Db | 11 | DIT | SKDCPNGHVCY | TKT | 1204.34 | 14.334 | 18.03 % |
Table 1: Promiscuous MHC ligands, having C-terminal ends are proteosomal cleavage sites of Long neurotoxin 2.
| Peptide Rank | Start Position | Sequence | Score | Predicted Affinity |
|---|---|---|---|---|
| 1 | 46 | PTVRTGVDI | 8.641 | High |
| 2 | 58 | STDDCDPFP | 8.636 | High |
| 3 | 47 | TVRTGVDIQ | 8.613 | High |
| 4 | 38 | DLGCAATCP | 8.549 | High |
| 5 | 24 | TWCDGFCSS | 8.534 | High |
| 6 | 61 | DCDPFPTRK | 8.499 | High |
| 7 | 50 | TGVDIQCCS | 8.385 | High |
| 8 | 57 | CSTDDCDPF | 8.151 | High |
| 9 | 7 | PDITSKDCP | 8.071 | High |
| 10 | 40 | GCAATCPTV | 8.010 | High |
| 11 | 44 | TCPTVRTGV | 7.852 | High |
| 12 | 36 | RVDLGCAAT | 7.814 | High |
| 13 | 30 | CSSRGKRVD | 7.684 | High |
| 14 | 25 | WCDGFCSSR | 7.557 | High |
| 15 | 29 | FCSSRGKRV | 7.536 | High |
| 16 | 51 | GVDIQCCST | 7.318 | High |
| 17 | 60 | DDCDPFPTR | 7.183 | High |
| 18 | 55 | QCCSTDDCD | 7.139 | High |
| 19 | 33 | RGKRVDLGC | 6.855 | High |
| 20 | 15 | PNGHVCYTK | 6.764 | High |
| 21 | 63 | DPFPTRKRP | 6.707 | High |
| 22 | 16 | NGHVCYTKT | 6.535 | High |
| 23 | 17 | GHVCYTKTW | 6.355 | High |
| 24 | 32 | SRGKRVDLG | 6.349 | High |
| 25 | 43 | ATCPTVRTG | 6.145 | High |
| 26 | 11 | SKDCPNGHV | 6.130 | High |
Table 2: Cascade SVM based High affinity TAP Binders of Long neurotoxin 2.
Solvent accessible regions
We predict Solvent accessibility by using Emani et al. [26] that predicts the highest probability i.e. Maximum 7.361, 32-SRGKRV-37 of a given protein region lies on the surface and Karplus and Schulz [27] predict backbone or chain flexibility which shows Maximum: 1.087 7-17 and 30-37, (Figures 5 and 6). we also predict the hydrophobic and hydrophilic characteristics of amino acids that are rich in charged and polar residues [28-37] (Figures 7-17). These scale predict the protein folding patterns, polar and nonpolar residue with a protein sequence such analysis has the goal of predicting membrane spanning segments hydrophobic or regions that are likely exposed on the surface of proteins (Hydrophilic domain) and therefore potentially antigenic.
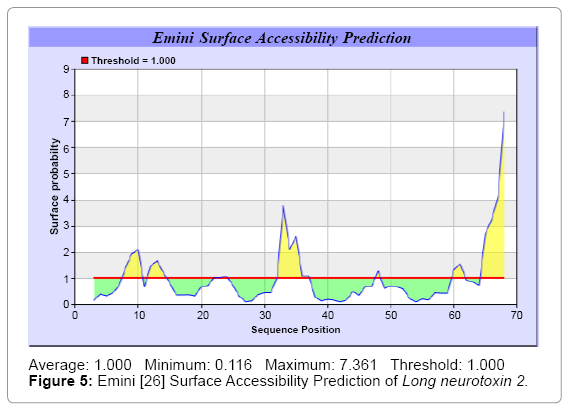
Figure 5: Emini [26] Surface Accessibility Prediction of Long neurotoxin 2.
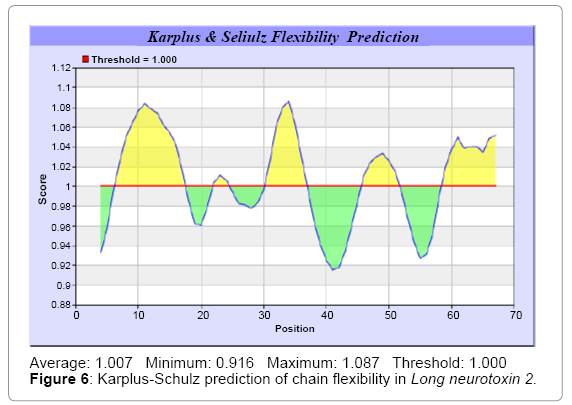
Figure 6: Karplus-Schulz prediction of chain flexibility in Long neurotoxin 2.
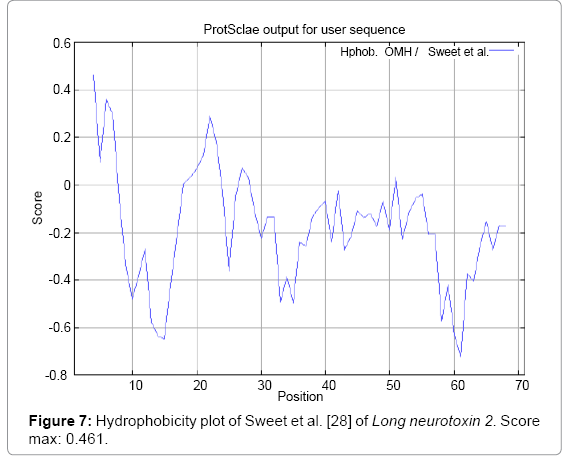
Figure 7: Hydrophobicity plot of Sweet et al. [28] of Long neurotoxin 2. Score max: 0.461.
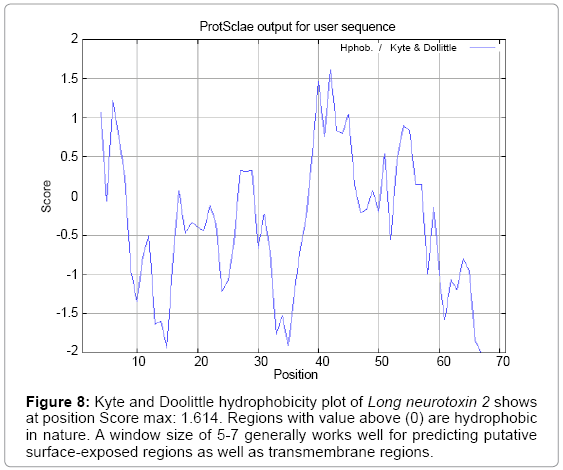
Figure 8: Kyte and Doolittle hydrophobicity plot of Long neurotoxin 2 shows at position Score max: 1.614. Regions with value above (0) are hydrophobic in nature. A window size of 5-7 generally works well for predicting putative surface-exposed regions as well as transmembrane regions.
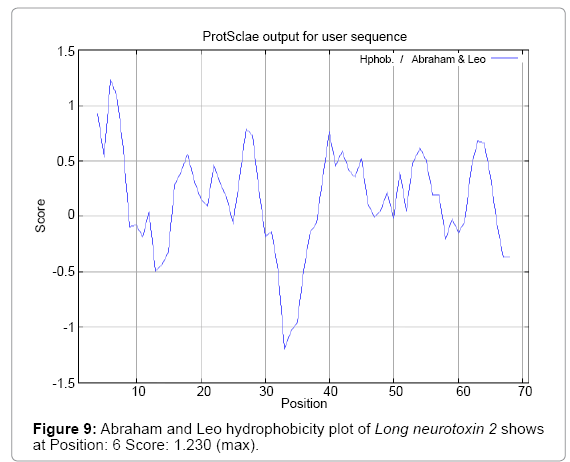
Figure 9: Abraham and Leo hydrophobicity plot of Long neurotoxin 2 shows at Position: 6 Score: 1.230 (max).
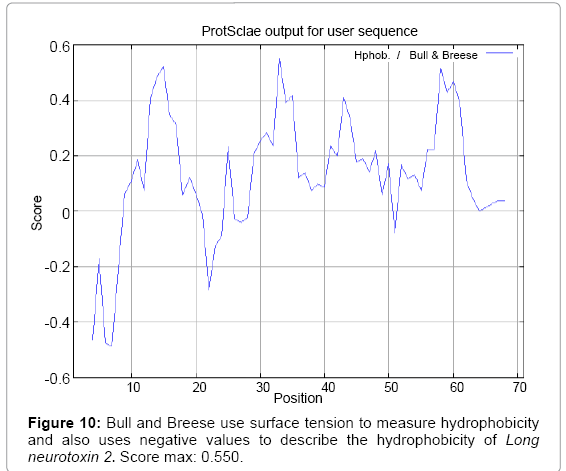
Figure 10: Bull and Breese use surface tension to measure hydrophobicity and also uses negative values to describe the hydrophobicity of Long neurotoxin 2. Score max: 0.550.
Figure 11: Hydrophobicity plot of Guy for Long neurotoxin 2.
Figure 12: Hydrophobicity plot of Miyazawa et al. [32] of Long neurotoxin 2, Score max: 6.737.
Figure 13: Hydrophobicity plot of Roseman MA [33] of Long neurotoxin 2. Score max: 0.334.
Figure 14: Hydrophobicity plot of Wolfenden et al. [34] of Long neurotoxin 2. Score max: 0.170.
Figure 15: Hydrophobicity/HPLC plot of Wilson et al. [35] of Long neurotoxin 2, Score max: 3.886.
Figure 16: Hydrophobicity/HPLC pH 3.4/plot of Cowan [36] of Long neurotoxin 2, Score max: 0.899.
Figure 17: Hydrophobicity plot of Chothia [37] of Long neurotoxin 2, Score max: 0.400.
We found the antigenic determinants by finding the area of greatest local hydrophilicity. The Hopp and Woods [19], Welling et al. [20], Parker et al. [21], Kolaskar and Tongaonkar [22] antigenicity scales (Figures 1-4) were designed to predict the locations of antigenic determinants in Long neurotoxin 2. Further this region form beta sheet which show high antigenic response than helical region of this peptide and shows highly antigenicity. We predict Solvent accessibility by using Emani et al. [26] that predicts the highest probability and Karplus and Schulz [27] predict backbone or chain flexibility (Figures 5 and 6). Solvent accessibility used to identify active site of functionally important residues in membrane proteins. Solvent-accessible surface areas and backbone angles are continuously varying because proteins can move freely in a three-dimensional space. The mobility of protein segments which are located on the surface of a protein due to an entropic energy potential and which seem to correlate well with known antigenic determinants. We also found the Sweet hydrophobicity, Kyte and Doolittle hydrophobicity; Abraham and Leo, Bull and Breese hydrophobicity, Guy, Miyazawa hydrophobicity, Roseman hydrophobicity, Wolfenden hydrophobicity, Wilson HPLC hydrophobicity, Cowan HPLC pH7.5 hydrophobicity, Chothia hydrophobicity scales (Figures 7-17). Theses scales are a hydrophilic with a polar residues assigned negative value. Because the N- and C-terminal regions of proteins are usually solvent accessible and unstructured, antibodies against those regions recognize the antigenic protein. In this study, we found predicted MHC-I peptide binders of toxin protein for 11mer_H2_Db, 10mer_H2_Db, 9mer_H2_Db, 8mer_H2_Db alleles (Table 1). We use a cascade SVM based TAPPred method which represents predicted TAP binders residues which occur at N and C termini from Long neurotoxin 2 (Table 2). T cell immune responses are derived by antigenic epitopes hence their identification is important for design synthetic peptide vaccine. T cell epitopes are recognized by MHCI molecules producing a strong defensive immune response against Long neurotoxin 2. Therefore, the prediction of peptide binding to MHCI molecules by appropriate processing of antigen peptides occurs by their binding to the relevant MHC molecules. Because, the C-terminus of MHCI-restricted epitopes results from cleavage by the proteasome and thus, proteasome specifity is important for determing T-cell epitopes. Consequently, RANKPEP server based prediction also focus on the prediction of conserved epitopes.
From the above result and discussion it is concluded that the ability of RANKPEP to predict MHC binding peptides, and thereby potential T-cell epitopes, Antigenic peptide that binds to MHC molecule are antigenic that means hydrophilic in nature. This means the increase in affinity of MHC binding peptides may result in enhancement of immunogenicity of Long neurotoxin 2, hence helpful in the designing of synthetic peptide vaccine.
This method will be useful in immunology, Vaccine design, immunodiagnostics, immunotherapeutic and molecular understanding of autoimmune susceptibility. Long neurotoxin 2 sequence involved multiple antigenic components to direct and empower the immune system to protect the host. MHC molecules which take active part in host immune response and involvement of MHC class in response to almost all antigens and it give effects on specific sites. Predicted MHC binding regions acts like red flags for antigen specific and generate immune response against the parent antigen. So a small fragment of antigen can induce immune response against whole antigen. The method integrates prediction of peptide MHC class binding; proteosomal C terminal cleavage and TAP transport efficiency. This theme is implemented in designing subunit and synthetic peptide vaccines.