Cell & Developmental Biology
Open Access
ISSN: 2168-9296
ISSN: 2168-9296
Research Article - (2013) Volume 2, Issue 1
The histological and histochemical studies were carried out on 60 camel embryos and foetuses eye balls of different Crown Vertebral Rump Length (CVRL). Paraffin sections were prepared and subjected to different stains. The present study revealed that, the optic pit was detected at 0.8 cm CVRL, as shallow groove on either side of the neural folds. The optic vesicle was observed at 1.5 cm CVRL, comprising from neuroectodermal cells, which became modified into a crescentic cup-like structure representing the optic cup at 2 cm CVRL. The optic cup was comprised from outer pigmented epithelium and the inner neuroblastic layer. The ganglion cells were distinctly differentiated at 6 cm CVRL, which separated from neuroblastic cells by the inner plexiform layer. The inner nuclear layer was firstly observed at 30 cm CVRL, which exhibited its full differentiation at 35 cm CVRL and became separated from the outer nuclear layer by the outer plexiform layer. First appearance of the photoreceptor cells was observed at 55 cm CVRL, which differentiated into outer and inner segments at 74 cm CVRL. All the layers of the retina were completely differentiated at 98 cm CVRL, comprising ten retinal layers.
<Keywords: Optic pit, Optic vesicle, Optic cup, Retinal cells, Photoreceptors
Knowledge of induction, neuron proliferation and differentiation of different cell types of retina is very important to explain the causes of some congenital anomalies that cause significant visual deficits as retinal separation and immaturation of photoreceptor cells [1,2]. Development of the mammalian retina is a highly organized process which originating from neuroectodermal cells through induction of the optic pit, optic vesicle and the optic cup [3-5]. A few available literatures described the histogenesis of camel retina as Ahmed et al. [6] and Abdel-Moniem [7].
The present work was undertaken with the aim of establishing the early prenatal histological and histochemical changes of camel’s retina associating with the induction, proliferation and differentiation of the retina.
The current study was carried out on the eye balls of 60 one humped camel embryos and foetuses, collected from El-Basateen slaughterhouse. Their Crown Vertebral Rump Lengths (CVRL) were ranged from 0.8 cm to 115 cm. The collected specimens were divided according to their CVRL into three groups: Group A, whole embryos from 0.8 to 5 cm CVRL were immersed directly into 10% Neutral Buffered Formalin (NBF). Group B, the head of embryos from 6 to 13 cm CVRL were directly immersed in the NBF. Group C, foetuses more than 15 cm CVRL, the eye ball of foetuses were gently enucleated and injected through sclera close to the limbus with NBF [8]. After collection of the samples, they were immersed directly into 10% NBF for at least 7 days. The preserved samples were dehydrated into a graded series of ethanol then absolute alcohol, then cleared in 3 changes of xylene, then embedded in paraffin wax. The blocks were serially sectioned at 4-5 μm thickness. The paraffin sections were subjected to the following stains; Harri’s Hematoxylin and Eosin (H&E) [9], Periodic-Acid Schiff (PAS) [10], Alcian blue technique (pH 2.5) [10,11], Heidenhain’s iron hematoxylin stain [12], and Mallory’s Phosphotungstic Acid- Hematoxylin method [13]. The afore-mentioned techniques were followed as outlined after [14].
At 0.8 cm CVRL stage
The optic pits were detected as a pair of shallow groove (stalk like) on either side of the neural folds (Figure 1). The wall of the neural fold and optic pit were formed from neuroectodermal cells (neuroepithelial) with condensed, moderately basophilic, oval nuclei and faintly eosinophilic cytoplasm (Figure 2). The uppermost cell layer facing the neural groove showed fine, eosinophilic, cilia-like processes (Figure 2).
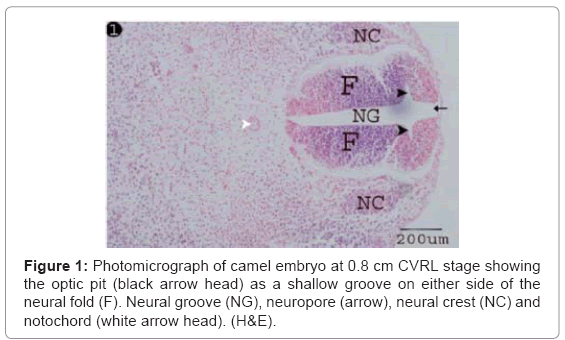
Figure 1: Photomicrograph of camel embryo at 0.8 cm CVRL stage showing the optic pit (black arrow head) as a shallow groove on either side of the neural fold (F). Neural groove (NG), neuropore (arrow), neural crest (NC) and notochord (white arrow head). (H&E).
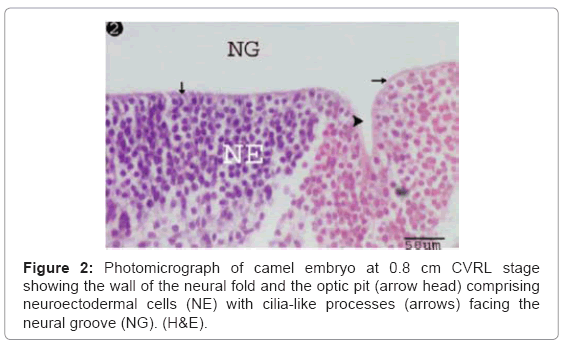
Figure 2: Photomicrograph of camel embryo at 0.8 cm CVRL stage showing the wall of the neural fold and the optic pit (arrow head) comprising neuroectodermal cells (NE) with cilia-like processes (arrows) facing the neural groove (NG). (H&E).
At 1.5 cm CVRL stage
The optic vesicle was formed as a result of aggregation of neuroectodermal cells at lateral sides of the primitive fore-brain, which made up of highly stratified undifferentiated neuroectodermal cells encircling an irregular lumen. The optic vesicle was closely subjacent to the surface epithelial ectodermal cells (Figure 3). The neuroectodermal cell layers were comprised closely packed cells with polymorphic, moderately basophilic nuclei and slightly acidophilic cytoplasm (Figure 4). The uppermost cell layer was bearing fine, cilia-like processes emerging into the vesicular lumen (Figure 4).
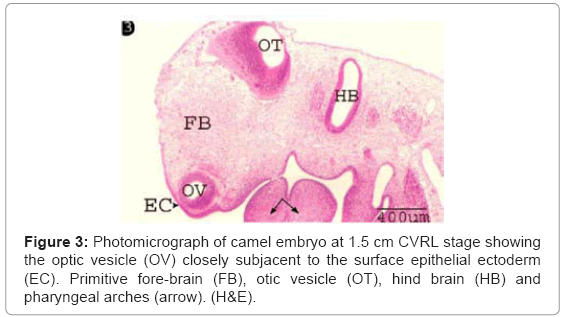
Figure 3: Photomicrograph of camel embryo at 1.5 cm CVRL stage showing the optic vesicle (OV) closely subjacent to the surface epithelial ectoderm (EC). Primitive fore-brain (FB), otic vesicle (OT), hind brain (HB) and pharyngeal arches (arrow). (H&E).
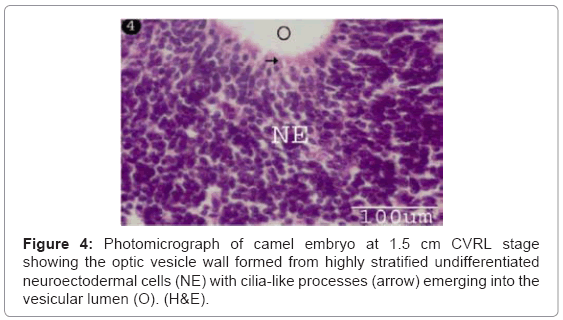
Figure 4: Photomicrograph of camel embryo at 1.5 cm CVRL stage showing the optic vesicle wall formed from highly stratified undifferentiated neuroectodermal cells (NE) with cilia-like processes (arrow) emerging into the vesicular lumen (O). (H&E).
At 2 cm CVRL stage
The optic cup was firstly observed as a crescentic cup-like structure comprising an outer, thinner pigmented cell layer, and an inner, much thicker one separated by a narrow intraretinal residual space (Figure 5). The outer thinner layer was formed from a single layer of pigmented cells “the pigmented retinal epithelium” (Figures 5 and 6). The pigmentation among the latter was increased at the rim of the optic cup (Figure 5). The inner thick layer “the presumptive neural retina” was representing the bulk of the cellular elements of the optic cup. Its thickness was thinnest at the rim, thicker at the sides and thickest at the bottom of the cup (Table 1) (Figure 5).
| CVRL (CM) |
RP | NF | OLM | ILM | GC | IP | GC SIZE | ON | IN | OP | P |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 2 | 8.31 | 20.5 | 1.5 | 0.9 | ------ | ------ | ------ | ------ | ------ | ------ | ------ |
| 6 | 9.03 | 25.14 | 1.92 | 1.4 | 40.05 | 3.78 | 3.12 | ------ | ------ | ------ | ------ |
| 9 | 9.01 | 30.44 | 1.95 | 1.62 | 30.41 | 6.5 | 3.8 | ------ | ------ | ------ | ------ |
| 16 | 8.88 | 32.2 | 2 | 1.69 | 24.14 | 8.29 | 4.24 | ------ | ------ | ------ | ------ |
| 22 | 8.78 | 35.12 | 2.04 | 1.83 | 13.18 | 8.92 | 4.5 | ------ | ------ | ------ | ------ |
| 27 | 8.7 | 35.53 | 2.51 | 1.87 | 12.01 | 9.35 | 4.7 | ------ | ------ | ------ | ------ |
| 30 | 8 | 35.54 | 2.72 | 1.92 | 11.9 | 18.9 | 4.8 | 77.03 | 13.7 | 2.2 | ------ |
| 35 | 6.02 | 35.6 | 2.82 | 1.94 | 11.5 | 20.73 | 4.88 | 55.77 | 50.77 | 7.15 | ------ |
| 50 | 5.51 | 37.18 | 3.01 | 2.01 | 10.01 | 26.03 | 5.18 | 58.61 | 39.57 | 10.04 | ------ |
| 55 | 5.5 | 37.19 | 3.04 | 2.24 | 9.58 | 26.41 | 5.22 | 58.82 | 38.21 | 10.26 | 4.35 |
| 60 | 5.1 | 38.23 | 3.21 | 1.52 | 9.01 | 26.5 | 5.46 | 60.34 | 38.01 | 13.62 | 10.50 |
| 74 | 4.9 | 38.5 | 2.25 | 1.32 | 8.09 | 30.4 | 6.2 | 49.75 | 32.09 | 15.03 | 20.98 |
| 98 | 4.21 | 40.57 | 0.95 | 0.83 | 7.9 | 35.92 | 6.96 | 49.03 | 32.01 | 15.04 | 22.13 |
| 105 | 3.5 | 40.59 | 0.7 | 0.75 | 7.3 | 33.16 | 7.01 | 44.45 | 28.32 | 14.2 | 22.5 |
| 115 | 3.2 | 42.12 | 0.65 | 0.7 | 7.01 | 31.02 | 7.01 | 30.11 | 14.26 | 12.42 | 24.01 |
Table 1: Showing the quantitative changes in the thickness (μm) of retinal pigmented (RP), nerve fiber layer (NF), outer limiting membrane (OLM), inner limiting membrane (ILM), ganglion cell layer (GC), inner plexiform layer (IP), ganglion cell size in posterior retina, outer nuclear layer (ON), inner nuclear layer (IN), outer plexiform layer (OP) and photoreceptors (P).
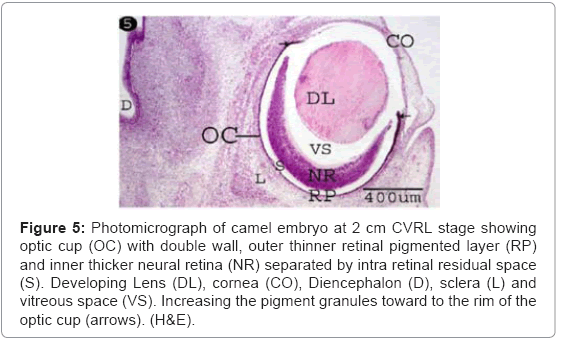
Figure 5: Photomicrograph of camel embryo at 2 cm CVRL stage showing optic cup (OC) with double wall, outer thinner retinal pigmented layer (RP) and inner thicker neural retina (NR) separated by intra retinal residual space (S). Developing Lens (DL), cornea (CO), Diencephalon (D), sclera (L) and vitreous space (VS). Increasing the pigment granules toward to the rim of the optic cup (arrows). (H&E).
The developing neural retina was differentiated into three distinct layers: a) an outer, thinner and esinophilic limiting membrane facing the residual space, b) an inner, thicker neurofilamentous layer representing the nerve fibers facing the inner vitrous space, and c) a middle, thickest, markedly stratified neuroblastic epithelial cell layer whose nuclear profiles were arranged into 15-25 layers (Figure 6). Depending upon the density of the nuclear elements, the middle layer was differentiated into three ill-distinct zones; zone I, was the thickest, condensed and showed numerous mitotic figures. Zone II, was the thinnest and showed less densely packed nuclei, and zone III, subjacent to the inner neurofilamentous layer (nerve fiber layer) (Figure 6).
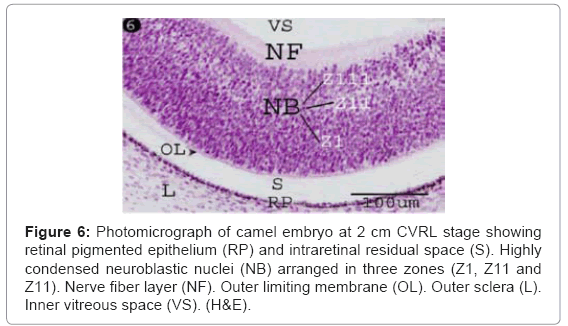
Figure 6: Photomicrograph of camel embryo at 2 cm CVRL stage showing retinal pigmented epithelium (RP) and intraretinal residual space (S). Highly condensed neuroblastic nuclei (NB) arranged in three zones (Z1, Z11 and Z11). Nerve fiber layer (NF). Outer limiting membrane (OL). Outer sclera (L). Inner vitreous space (VS). (H&E).
At 6-27 cm CVRL stage
The ganglion blastic cells were distinctly differentiated from the neuroblastic cells forming individual layer. The latter was arranged in about three to four nuclear layers with spherical, less condensed and polymorphic nuclei with thickness 40.05 μm, and with size 3.12 μm at 6 cm (Table 1) (Figure 7). The ganglion blastic cells were separated from the neuroblastic layer by well demarcated inner plexiform. The nerve fiber layer was appeared thicker than the prevoius stage, which limited by the inner limiting membrane facing the vitreous space while the neuroblastic cell layers were limited by the outer limiting membrane facing the pigmented epithelium (Figure 7). The nerve fiber layer and the inner plexiform layer were exhibited moderately alcianophilia (Figure 8). The outer limiting membrane showed strong positive PAS reaction while the inner plexiform layer showed slight reaction (Figure 9). High mitotic activity was observed in the neuroblastic cells nuclei as black dots representing the chromatin granules (Figure 10). Displaced ganglion cells were located in the inner plexiform layer representing the amacrine cells. The latter showed large basophilic nuclei, remnant eosinophilic cytoplasm and fine cytoplasmic processes connecting with the neuroblastic cells (Figure 11).
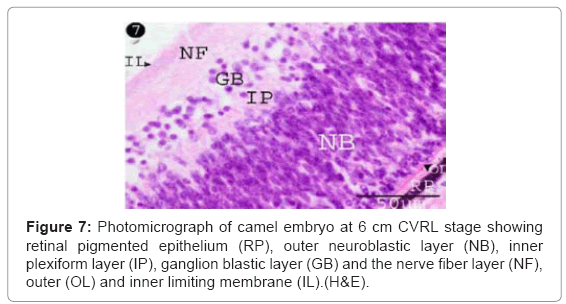
Figure 7: Photomicrograph of camel embryo at 6 cm CVRL stage showing retinal pigmented epithelium (RP), outer neuroblastic layer (NB), inner plexiform layer (IP), ganglion blastic layer (GB) and the nerve fiber layer (NF), outer (OL) and inner limiting membrane (IL).(H&E).
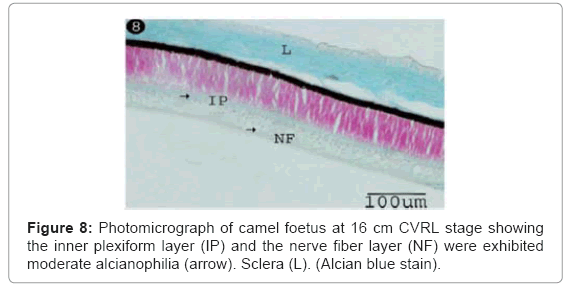
Figure 8: Photomicrograph of camel foetus at 16 cm CVRL stage showing the inner plexiform layer (IP) and the nerve fiber layer (NF) were exhibited moderate alcianophilia (arrow). Sclera (L). (Alcian blue stain).
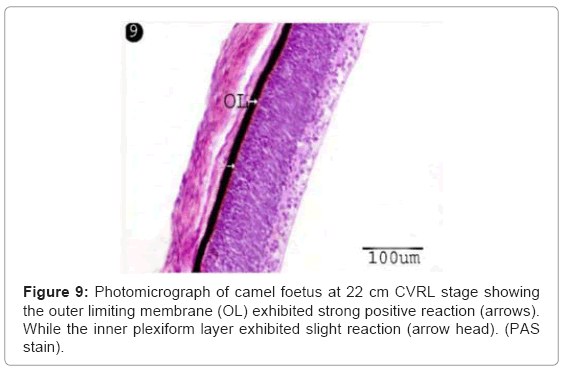
Figure 9: Photomicrograph of camel foetus at 22 cm CVRL stage showing the outer limiting membrane (OL) exhibited strong positive reaction (arrows). While the inner plexiform layer exhibited slight reaction (arrow head). (PAS stain).
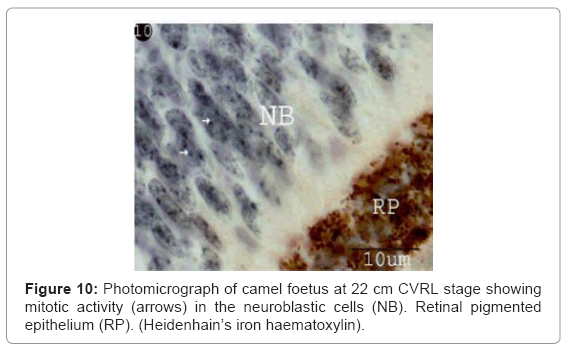
Figure 10: Photomicrograph of camel foetus at 22 cm CVRL stage showing mitotic activity (arrows) in the neuroblastic cells (NB). Retinal pigmented epithelium (RP). (Heidenhain’s iron haematoxylin).
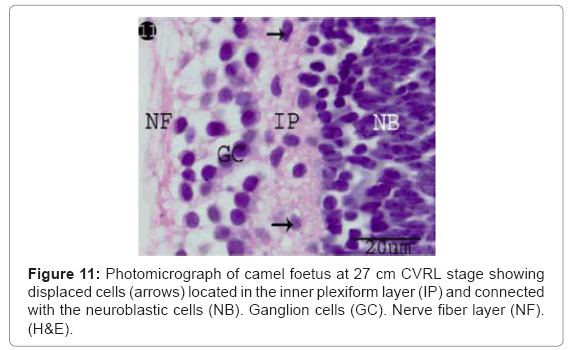
Figure 11: Photomicrograph of camel foetus at 27 cm CVRL stage showing displaced cells (arrows) located in the inner plexiform layer (IP) and connected with the neuroblastic cells (NB). Ganglion cells (GC). Nerve fiber layer (NF). (H&E).
At 27 cm CVRL stage, the thickness of the ganglion cell layers were decreased while the ganglion cell size was increased (Table 1) (Charts 1 and 2).
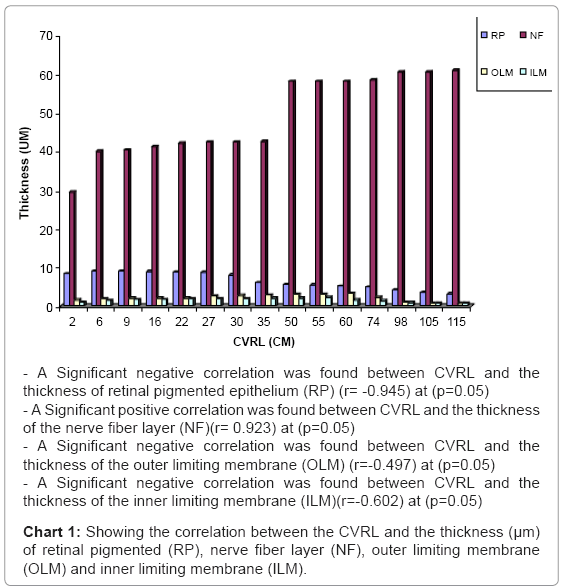
Chart 1: Showing the correlation between the CVRL and the thickness (μm) of retinal pigmented (RP), nerve fiber layer (NF), outer limiting membrane (OLM) and inner limiting membrane (ILM).
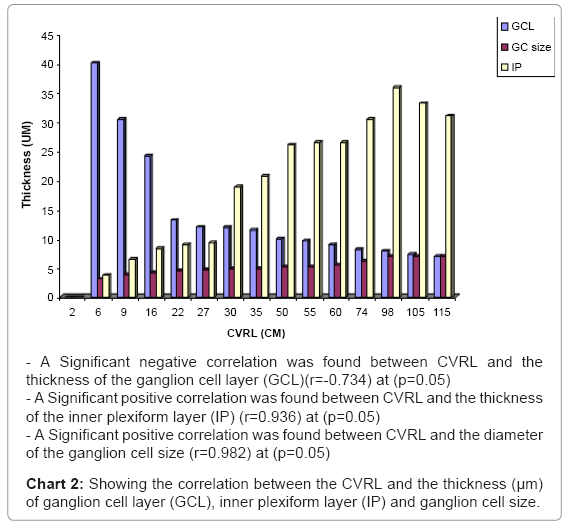
Chart 2: Showing the correlation between the CVRL and the thickness (μm) of ganglion cell layer (GCL), inner plexiform layer (IP) and ganglion cell size.
At 30 cm CVRL stage
The cells in the inner third of neuroblastic layer became slightly differentiated into ill-defined layer. The latter was comprised less condensed nuclei representing the inner nuclear layer (middle nuclear layer) with thickness of 13.7 μm (Table 1). Thin-walled blood vessels were firstly appeared, lining by flat nuclei, spreading in the nerve fiber layer and surrounded by dispersed oval nuclei (Figure 12).
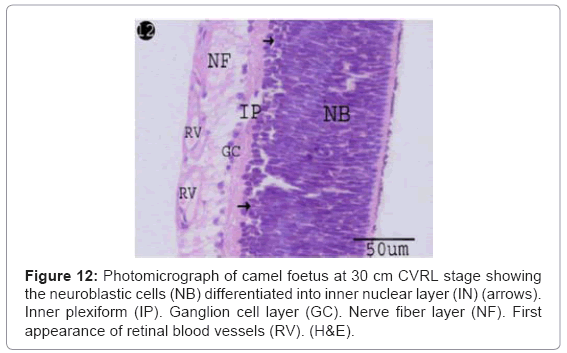
Figure 12: Photomicrograph of camel foetus at 30 cm CVRL stage showing the neuroblastic cells (NB) differentiated into inner nuclear layer (IN) (arrows). Inner plexiform (IP). Ganglion cell layer (GC). Nerve fiber layer (NF). First appearance of retinal blood vessels (RV). (H&E).
At 35-50 cm CVRL stage
The inner nuclear layers were distinctly differentiated and separated from the outer nuclear layer by newly formed outer plexiform layer (Figures 13 and 14), which contained displaced cells nuclei (horizontal cells) with round euchromatic nuclei and apical fine processes connecting with outer nuclear layer (Figures 13 and 14). The inner nuclear cells were arranged in 8-10 rows with polymorphic, light stained nuclei and low mitotic figure, while the outer nuclear cells were arranged in 12-14 rows with oval tapered, dark stained nuclei and high mitotic activity (Figures 13 and 14).
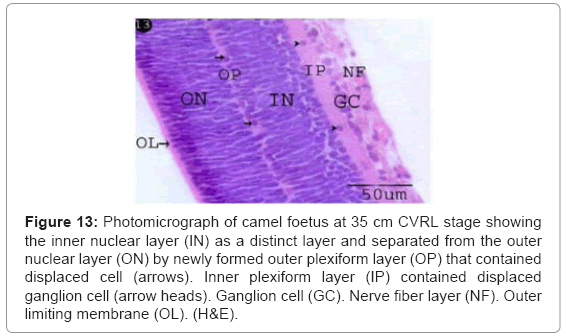
Figure 13: Photomicrograph of camel foetus at 35 cm CVRL stage showing the inner nuclear layer (IN) as a distinct layer and separated from the outer nuclear layer (ON) by newly formed outer plexiform layer (OP) that contained displaced cell (arrows). Inner plexiform layer (IP) contained displaced ganglion cell (arrow heads). Ganglion cell (GC). Nerve fiber layer (NF). Outer limiting membrane (OL). (H&E).
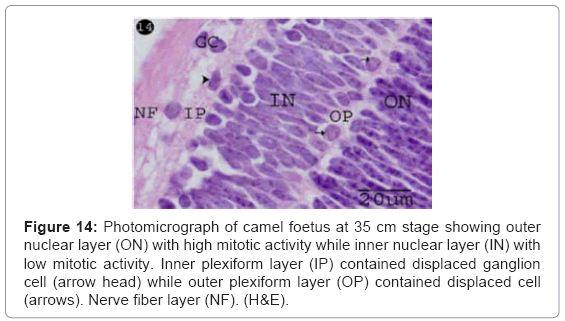
Figure 14: Photomicrograph of camel foetus at 35 cm stage showing outer nuclear layer (ON) with high mitotic activity while inner nuclear layer (IN) with low mitotic activity. Inner plexiform layer (IP) contained displaced ganglion cell (arrow head) while outer plexiform layer (OP) contained displaced cell (arrows). Nerve fiber layer (NF). (H&E).
In this stage, marked quantitative changes were observed within the developing retinal layers in the thickness (the pigmented epithelium, ganglion cell layer and the inner nuclear layer were decreased while the outer nuclear layer , the inner and outer plexiform layer and nerve fiber layer were increased) (Table 1) (Charts 1-3).
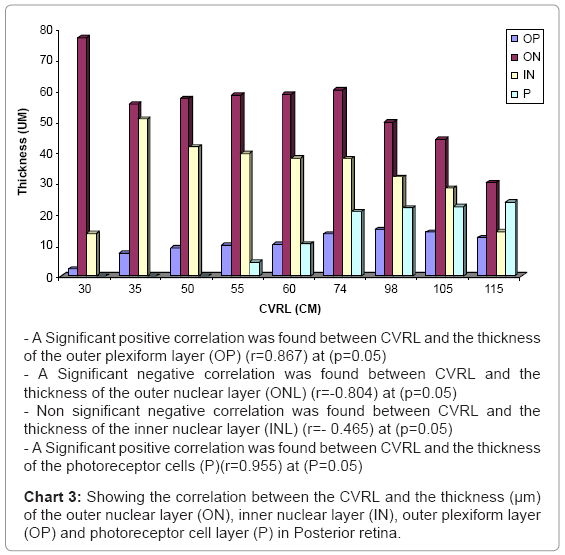
Chart 3: Showing the correlation between the CVRL and the thickness (μm) of the outer nuclear layer (ON), inner nuclear layer (IN), outer plexiform layer (OP) and photoreceptor cell layer (P) in Posterior retina.
At 55 cm CVRL stage
First appearance of the photoreceptor cells was observed with thickness 4.35 μm (Table 1) (Chart 3) as esinophilic film resting on thin outer limiting membrane (Figure 15). Spherical cells (horizontal cells) were located in the outer plexiform layer which connecting with the outer nuclear layer (Figure 15). Blue oval dots were observed in the photoreceptors layer indicating the presence of the mitochondria (Figure 16).
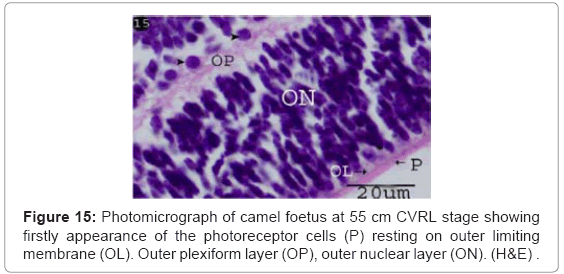
Figure 15: Photomicrograph of camel foetus at 55 cm CVRL stage showing firstly appearance of the photoreceptor cells (P) resting on outer limiting membrane (OL). Outer plexiform layer (OP), outer nuclear layer (ON). (H&E) .
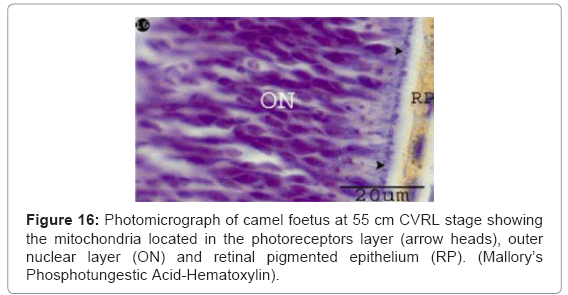
Figure 16: Photomicrograph of camel foetus at 55 cm CVRL stage showing the mitochondria located in the photoreceptors layer (arrow heads), outer nuclear layer (ON) and retinal pigmented epithelium (RP). (Mallory’s Phosphotungestic Acid-Hematoxylin).
At 60 cm CVRL stage
Photoreceptor cells were increased in the thickness 5.35 μm (Table 1) (Chart 3). These cells became differentiated into rod and cone cells. The rod cells were formed from long and thin processes. While the cone cells were formed from short and conical shaped processes (Figure 17).
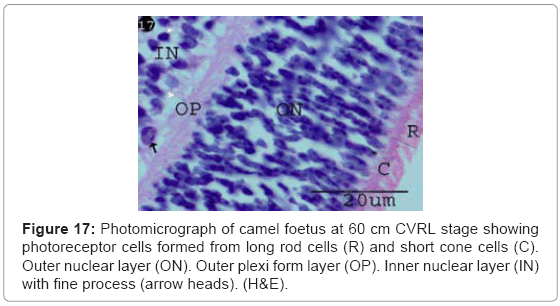
Figure 17: Photomicrograph of camel foetus at 60 cm CVRL stage showing photoreceptor cells formed from long rod cells (R) and short cone cells (C). Outer nuclear layer (ON). Outer plexi form layer (OP). Inner nuclear layer (IN) with fine process (arrow heads). (H&E).
The photoreceptor cell and inner limiting membrane showed strong alcianophilia indicated high amount of acidic mucopolysaccharides. While slight alcianophilia was observed in the outer, inner plexiform layer and the nerve fibre layer indicated fewer amounts of acidic mucopolysaccarides (Figure 18).
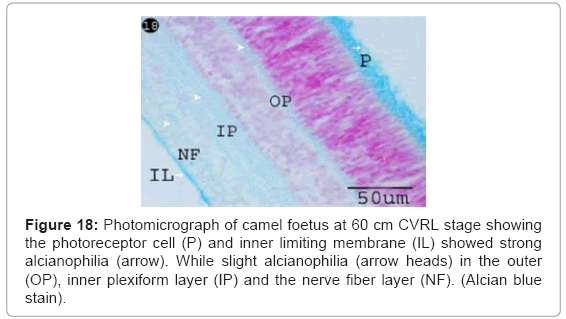
Figure 18: Photomicrograph of camel foetus at 60 cm CVRL stage showing the photoreceptor cell (P) and inner limiting membrane (IL) showed strong alcianophilia (arrow). While slight alcianophilia (arrow heads) in the outer (OP), inner plexiform layer (IP) and the nerve fiber layer (NF). (Alcian blue stain).
At 74 cm CVRL stage
The photoreceptor cells became more differentiated into rods (long process) and cones cells (short process). The latter cells were less in the number than the rods cells (Figure 19). The packed components of the photoreceptors (rod and cone cells) were differentiated into an outer and inner segment. The narrow basal slight basophilic zone that closed to the outer limiting membrane was represented the inner segment while the wide apical acidophilic zone was represented the outer segment (Figure 19).
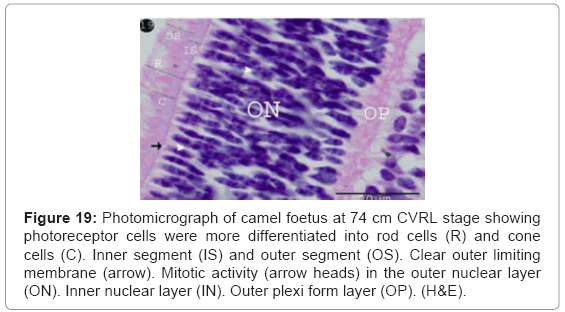
Figure 19: Photomicrograph of camel foetus at 74 cm CVRL stage showing photoreceptor cells were more differentiated into rod cells (R) and cone cells (C). Inner segment (IS) and outer segment (OS). Clear outer limiting membrane (arrow). Mitotic activity (arrow heads) in the outer nuclear layer (ON). Inner nuclear layer (IN). Outer plexi form layer (OP). (H&E).
At 98-115 cm CVRL stage
The full retinal layers were completely formed comprising ten retinal layers (retinal pigmented epithelium, photoreceptors, outer limiting membrane, outer nuclear layer, outer plexiform layer ,inner nuclear layer, inner plexiform layer, ganglion cells layer, nerve fibre layer and inner limiting membrane) with different thickness (Table 1) (Figure 20).
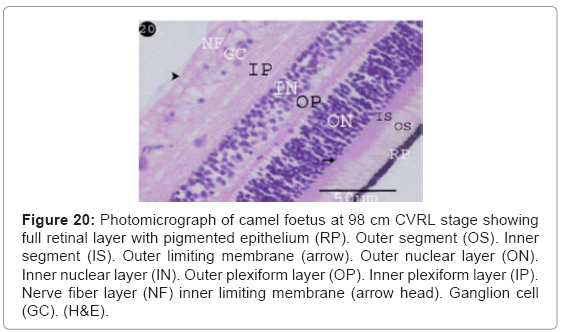
Figure 20: Photomicrograph of camel foetus at 98 cm CVRL stage showing full retinal layer with pigmented epithelium (RP). Outer segment (OS). Inner segment (IS). Outer limiting membrane (arrow). Outer nuclear layer (ON). Inner nuclear layer (IN). Outer plexiform layer (OP). Inner plexiform layer (IP). Nerve fiber layer (NF) inner limiting membrane (arrow head). Ganglion cell (GC). (H&E).
Strong alcianophilia was noticed in the outer segment of the photoreceptor cells and inner limiting membrane, while the outer and inner plexiform, the nerve fibre layer and outer limiting membrane was slight reaction. The inner segment of photoreceptor cells was negatively reacted (Figure 21).
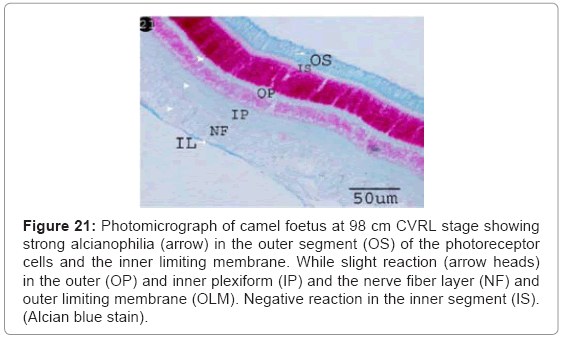
Figure 21: Photomicrograph of camel foetus at 98 cm CVRL stage showing strong alcianophilia (arrow) in the outer segment (OS) of the photoreceptor cells and the inner limiting membrane. While slight reaction (arrow heads) in the outer (OP) and inner plexiform (IP) and the nerve fiber layer (NF) and outer limiting membrane (OLM). Negative reaction in the inner segment (IS). (Alcian blue stain).
Strong PAS positive reaction was observed in the outer segment of the photoreceptors cells. While the inner segment, outer plexiform layer, inner plexiform layer and the nerve fibre layer showed slight reaction (Figure 22).
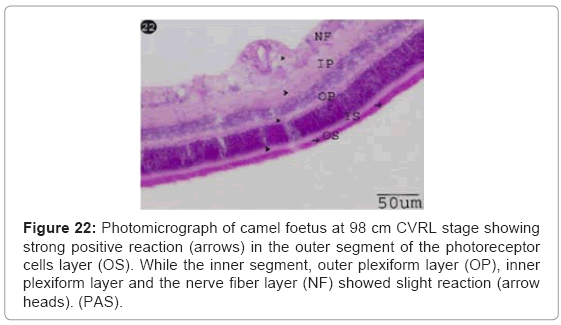
Figure 22: Photomicrograph of camel foetus at 98 cm CVRL stage showing strong positive reaction (arrows) in the outer segment of the photoreceptor cells layer (OS). While the inner segment, outer plexiform layer (OP), inner plexiform layer and the nerve fiber layer (NF) showed slight reaction (arrow heads). (PAS).
The small bodies of the mitochondria were increased and located in the inner segment of the photoreceptor cells. By Heidenhain’s iron hematoxylin stain, the mitochondria were appeared as black bodies (Figure 23). While by Mallory’s Phosphotungstic Acid-Hematoxylin method, they appeared as blue bodies (Figure 24). The outer segment of the photoreceptors sells was negatively reacted (Figures 23 and 24).
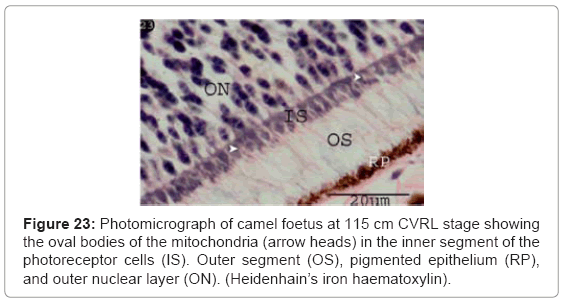
Figure 23: Photomicrograph of camel foetus at 115 cm CVRL stage showing the oval bodies of the mitochondria (arrow heads) in the inner segment of the photoreceptor cells (IS). Outer segment (OS), pigmented epithelium (RP), and outer nuclear layer (ON). (Heidenhain’s iron haematoxylin).
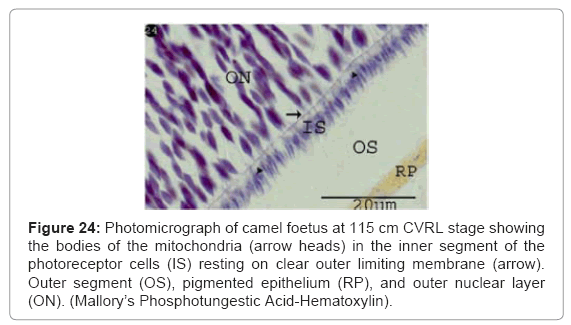
Figure 24: Photomicrograph of camel foetus at 115 cm CVRL stage showing the bodies of the mitochondria (arrow heads) in the inner segment of the photoreceptor cells (IS) resting on clear outer limiting membrane (arrow). Outer segment (OS), pigmented epithelium (RP), and outer nuclear layer (ON). (Mallory’s Phosphotungestic Acid-Hematoxylin).
At 0.8 cm CVRL, the present study detected the optic pits as a pair of shallow groove on either side of the neural folds. This finding was in agreement with Miyata, Samuel et al. in rat, Sharma and Ehinger and Fletcher and Weber in human [5,15-17].
At 1.5 cm CVRL, our results had revealed that the optic vesicle was formed as a result of aggregation of neuroectodermal cells at lateral sides of primitive fore-brain. Similar finding was also obtained by Kaufman et al. [3] and Moshiri et al. [4] in human. In contrast Abdel-Moniem stated that the optic vesicle is detected at 2 cm CVRL camel embryos [7].
The obtained results indicated that, the optic cup was firstly observed at 2cm CVRL, as a crescentic cup-like structure comprising an outer, thinner pigmented cell layer, and an inner, much thicker one forming the neural retina. This finding was similar to those of Ahmed et al. [6] in camel. Meanwhile Abdel-Moniem [7] mentioned that the optic cup is observed at 3.8 cm CVRL camel embryos. This different observation may occur as result of different sagittal histological sections and the stage between the 2 cm CVRL and 4 cm CVRL may called the optic cup stage with constant structure.
The current study observed numerous mitotic figures among the cells of the neuroblastic cells that located subjacent to the outer limiting membrane. This finding was in agreement with Rapaport and Stone [18] in developing cat retina.
The differentiation of the ganglion cells was at 6 cm CVRL stage. Differentiation of the inner nuclear layer was visible at 30 cm CVRL that became well defined at 35 cm CVRL. The photoreceptor cells were detected at 55 cm CVRL that became clearly defined at 60 cm CVRL that differentiated into rods and cones cells. The outer segment and inner segment were detected at 74 cm CVRL. This finding was in contrast to that obtained by Ahmed et al. [6], whom mentioned that the retina could be differentiated into a photoreceptor cell layer (cones and rods), a neuroblastic layer or bipolar cell, a ganglion cell, fibrous layer, inner limiting membrane at 20 cm in camel embryos. The finding was similar to that mentioned by Abdel-Moniem [7] in camel.
Our study observed that the ganglion cells were the first cells which developed and increased in size with direct proportion to the CVRL. This trend was found in cat, monkey and rat by Walsh et al., Walsh and Polley and Rapaport et al. and Rapaport et al. [19-22], whom mentioned that the small retinal ganglion cells were generated early and increase in size at later developmental stages.
The present work observed that the retina reached its maturity at 80-115 cm CVRL stage. Our finding was similar to that observed by Abdel-Moniem [7] in camel. This finding emphasized that the retina reached the maturity before birth that explained why the camel is able to see directly after birth. In contrast, the maturity was occurred in dog at 80 days post conception [23], and at 12 days old rat [24]. This result was also emphasized by Thomas and Alvin who stated that the ungulate retina is mature at birth, but the carnivore retina does not fully mature until about 5 weeks postnatal [17].
Our study emphasized that the retina at 20 cm CVRL, showed retinal pigmented epithelium, outer limiting membrane, inner limiting membrane, neuroblastic cells, inner plexiform layer, ganglion cell layer and nerve fiber layer (7 layers only). The result is not in agreement with that described by Ahmed et al. [6] who mentioned that the retinal showed nearly all its normally 10 layers at 20 cm CVRL camel embryos. Our investigation has clarified that the retinal 10 layers were appeared at 55-74 cm CVRL. This result was similar to that recorded by Abdel-Moniem [7] in camel.
showed nearly all its normally 10 layers at 20 cm CVRL camel embryos. Our investigation has clarified that the retinal 10 layers were appeared at 55-74 cm CVRL. This result was similar to that recorded by Abdel-Moniem [7] in camel.
By Heidenhain’s iron haematoxylin stain and Mallory’s Phosphotungstic Acid-Hematoxylin, the mitochondria were detected as small oval bodies in the inner segments of the photoreceptor cells, while the outer segment showed negative reaction. This observation might lead us to suggest that the photoreceptors mitochondria were located in the inner segments. This finding was in agreement to those of Stone et al. [25] whom stated the mitochondria are sequestered to the ends of the cell or to the inner segments and in some species extend to axon terminals in the outer plexiform layer. The later authors hypothesized that the mitochondria migrated to these locations towards sources of oxygen.
The reaction of the various layers of the retina to PAS and alcian blue stains were not described in any of the available literature. The present study detected positive reaction to PAS in the outer limiting membrane at 22 cm CVRL stage. At 98 cm CVRL, the outer segments of the photoreceptor cells showed strong positive reaction indicating their content of neutral mucopolysaccarides. AT 98 cm CVRL, the outer segments of the photoreceptor cells layer were reacted to alcian blue stain indicating their content of acidic mucopolysaccarides. The inner segments of the photoreceptor cells were negatively reacted with PAS and alcian blue stains. This observation might lead us to suggest that the photoreceptors contained large amount of protein in the outer segments.