Journal of Cell Science & Therapy
Open Access
ISSN: 2157-7013
ISSN: 2157-7013
Research Article - (2018) Volume 9, Issue 3
Keywords: LncRNA; ZFAS1; Cervical cancer; Prognosis; Chemosensitivity; Cisplatin
Cervical cancer, one of the most common female malignant tumors, accounts for over half of the reproductive system cancers in women, constituting a great threat to the health of women. [1] According to statistics, there are approximately 500 thousand new cases diagnosed and about 230 thousand women die of cervical cancer annually around the world. [2] The occurrence and development of cervical cancer has been considered as a complicated and multi-stage process resulted from a wide range of factors [3]. Although great progress has been made in the screening and vaccines for the prevention of cervical cancer in recent decades, and most women with early cervical cancers could be cured after surgery or radiotherapy, about 20% to 25% patients still suffered from treatment failure because of recurrence and distant metastasis [4,5]. Besides, a significant proportion of patients with cervical cancer patients are in an advanced stage at diagnosis, and chemotherapy becomes the major therapy for the treatment, but the drug resistance developed during cancer treatment would directly affect the therapeutic efficacy and prognosis [6]. Therefore, it is of great importance to improve the chemosensitivity of cervical cancer patients and to find out the appropriate predicators and therapeutic targets.
Long non-coding RNA (lncRNA) is a class of RNA molecules with the transcript length of more than 200 nucleotides and without the protein-coding function [7]. Recently, lncRNAs have been identified to play a vital role in the occurrence and progression of tumors and become a hot spot in the study of tumor diagnosis and treatment [8,9]. LncRNA Zinc finger antisense 1 (ZFAS1), a newly found lncRNA located at 20q13 and the antisense RNA of the gene Zinc finger NFX1- type containing 1 (ZNFX1), is stably expressed in many tissues and organs [10]. Recently, more and more studies conformably reported that ZFAS1 was aberrantly expressed in the progression of different types of tumors [11,12], which was found to be overexpressed in gastric, colonic, glioma, and ovarian cancers, functioning as a potential oncogene through promoting tumor cell growth and metastasis [13- 15]. Originally, ZFAS1 was found to be abnormally expressed in breast cancer tissues by Askarian-Amiri et al. [16]. Later, Fan et al. [13] also revealed the down-regulation of ZFAS1 in breast cancer tissues and cell lines, and over-expressed ZFAS1 can suppress cell proliferation and induce cell apoptosis via blocking EMT process [13], showing a potential role of ZFAS1 in gynecology oncology. Additionally, ZFAS1 is also dysregulated in other types of cancer. For example, Li et al. found ZFAS1 expression was significantly increased in hepatocellular carcinoma and closely related to intrahepatic and extrahepatic metastasis and poor prognosis of patients [17]. The up-regulated ZFAS1 can also act as an oncogene in colorectal cancer through destabilization of p53 or interaction with CDKl/cyclin B1 to promote cell cycle and inhibit cell apoptosis [18]. More importantly, the expression of ZFAS1 was pointed out to be up-regulated in ovarian cancer cells, which was closely associated with the cisplatin chemosensitivity [6]. Thus, we hypothesized that ZFAS1 may play a significant part in the growth and the chemosensitivity of cervical cancer. The purpose of the study was to investigate the expression of ZFAS1 in cervical cancer and to analyze its relationship with the prognosis and chemosensitivity to cervical cancer cells.
Ethics statement
This study was approved by the Ethics Committee of the First Affiliated Hospital of Soochow University and conformed to the protocols in the Helsinki declaration [19] and all patients signed the informed consent form prior to the study.
Study subjects
From December 2010 to December 2012, cervical cancer tissues and adjacent normal tissues were collected from 85 cervical cancer patients who underwent resection in our hospital and all cancer tissue samples were confirmed as cervical cancer by pathological examination. The subjects in this study were 24-60 years old, with the median age of 44.5 ± 1.24 years. According to the clinical staging criteria for cervical cancer put forward by the International Federation of Gynecology and Obstetrics (FIGO) [20], there were 58 cases at the I-II stage and 27 cases at the III-IV stage. Through histological grading, 19 cases were in G1 (well-differentiated), 35 cases in G2 (moderately-differentiated), and 31 cases in G3 (poorly-differentiated). As for the tumor diameter, there were 55 cases ≤ 4 cm and 30 cases >4 cm. Also, 33 patients had nymph node metastasis and 52 cases had no nymph node metastasis. In terms of the depth of myometrial invasion, 59 cases were <50% myometrial invasion and 26 cases were ≥ 50% myometrial invasion.
Cell selection and culture
Normal cervical epithelial cell line HaCaT and cervical cancer cell lines (CaSki, HeLa and C33A) were all purchased from the American Type Culture Collection (ATCC). Cell lines were cultured in DMEM medium (Gibco, USA) supplemented with 10% fetal bovine serum in an incubator with 5% CO2 at 37°C.
ZFAS1 expression detected by qRT-PCR
Total RNA was extracted from tissues and cells by using the Trizol method (Invitrogen, Carlsbad, CA) and determined for OD260/280 ratio (with a UV spectrophotometer) and RNA concentration, which was cryopreserved at -80°C for later use. The primer sequences were designed by the Primer 5.0 software as follows: ZFAS1 (forward 5’-AAGCCACGTGCAGACATCTA-3’ and reverse 5’-CTACTTCCAACACCCGCATT-3’), and β-actin (forward 5’-TGACGGGGTCACCCACACTGTGCCCATCTA-3’ and reverse 5’-CTAGAAGCATTTGCGGTGGACGATGGAGGG-3’). Those primers were synthesized by Sangon Biotech (Shanghai) Co., Ltd. The PCR amplification was performed in the ABI 7300 PCR instrument (ABI Company, USA) with β-actin as the internal reference gene. The relative expression level of target genes was calculated by using the 2-△△Ct method.
Prognosis and follow-up
The start of follow-up was the discharge date after systemic treatment and the end of follow-up was in December 2017. The follow-up was conducted by outpatient service, telephone interview or medical record review. The survival time was counted by month and the patient’s death was credited as the end of observation. The overall survival (OS) was defined as the duration from the end of systemic surgery treatment to the time when patients died of any causes.
Cell transfection and grouping
CaSki and HeLa cells were inoculated onto 6-well plate by 1 × 106 cells/well for 24 h of growth. Next, cells were transfected according to the instructions on the LipofectamineTM3000 Liposomal Transfection Reagent (Invitrogen, USA). After 48 h, cells were collected for later experiments and divided into three groups: Blank group (cells without any treatment), siR-Control group (cells transfected with negative control siRNA sequence), and siR-ZFAS1 group (cells transfected with ZFAS1 siRNA sequence). Both the ZFAS1 siRNA sequence and negative control siRNA sequence were provided by Guangzhou RiboBio Co., Ltd.
Wound-healing assay
The horizontal lines were drawn appropriately on the back of the 6-well plate with a marker pen and a ruler. Then, about 5 × 105 cells were added into the plate. Next day, a pipette tip was used to draw vertical lines along the ruler vertical to the lines on the back of the plate. Subsequently, cells were washed three times with PBS buffer to remove the detached cells, followed by the addition of serum-free medium for incubation at 37°C in 5% CO2. Samples and picture were taken at the 0 h, 24h and 48 h for analysis. Triplicate experiments were performed.
Transwell assay
The Transwell chamber was pre-coated by 100 μL Matrigel for incubation at 37°C. After the Matrigel was solidified, transfected cervical cancer cells were seeded into the chamber by 4 × 104 cells/well and 600 μL of usual medium was added for 48 h of incubation. Next, the chamber was taken out, then cells were fixed with paraformaldehyde and stained with crystal violet, and cells without migration were wiped off from the chamber with cotton swabs. After that, cells left in the chamber were observed and photographed under an optical microscope. The number of invasive cells were counted after randomly selecting 10 high power fields. Triplicate experiments were performed.
MTT assay
CaSki and HeLa cells of different transfection groups were seeded onto 96-well plates and incubated in 200 μl of medium with cisplatin (Sigma Aldrich, USA) (including 0, 0.5, 1, 2, 5, and 10 μmol/L). After 24 h, 48 h, 72 h and 96 h of culture, 20 μl of MTT solution (5 g/L) was added into each well for 4 h of incubation. Next, the cells were centrifuged and 150 μl of DMSO (dimethyl sulfoxide) was added for oscillation. The absorbance value (A value) at the wavelength of 490 nm was measured with a microplate reader. Cell inhibition rate (%) = (1- OD treatment group/ OD control group) × 100%. The drug concentration which led to 50% inhibition concentration (IC50) data was calculated using the weighted linear regression method with GraphPad 6.0 software. The experiment was repeated for three times.
Cell cycle assay
To evaluate cell cycle distributions, a total number of 5 × 106 cells were harvested after transfection for 48 h. Transfected cells were resuspended using PBS containing 70% ethanol for 48 h at 4°C and stained with 10 μg/ml propidium iodide and 0.5 mg/ml RNase A for 15 min at 37°C. Cell cycle was analyzed by FACS caliber flow cytometry (BD Biosciences, US.). Each experiment was done in triplicate.
Cell apoptosis
After the treatment with cisplatin for 48 h, CaSki and HeLa cells of different transfection groups were collected, washed once with PBS buffer, and suspended in 0.5 ml binding buffer. Next, 6 μl of FITCannexin V and 20 μl of propidium iodide (PI) (BD Company, USA) were added for 15 min of incubation at room temperature with the avoidance of light. Then, centrifugation was performed, the supernatant was removed, and 300 μl of binding buffer was added. Subsequently, cells were observed under the Epics XL flow cytometer (Beckman- Coulter Company) to detect the cell apoptosis rate (1 × 104 cells). Cell apoptosis index = number of apoptotic cells/number of total cells × 100%. The experiment was repeated three times.
Western blotting
Cells were harvested and the whole-cell lysates were prepared. The total protein concentrations were quantified by BCA protein assay kit (Thermo Scientific, USA). Proteins were resolved on a 10% sodium dodecyl sulfate-polyacrylamide gel (SDS-PAGE) and blocked with 5% non-fat milk and incubated with primary antibodies against PARP and cleaved caspase-3 (Cell Signaling Technology, USA) or β-actin (Abcam, USA). After rinsing with TBS, the membrane was incubated with secondary antibodies at room temperature for 30 min. Finally, the immunoreactivity was visualized with an enhanced chemiluminescence substrate (Thermo Scientific, USA).
Chemosensitivity experiments in nude mice with transplanted model of cervical cancer
Female BALB/c nude mice (4-6 weeks old) were bought from the Laboratory Animal Center of the First Affiliated Hospital of Soochow University. Animal care procedures and animal experiments in this study were approved by the Laboratory Animal Ethics Committee of the First Affiliated Hospital of Soochow University. HeLa cells, in the logarithmic growth phase from Blank group, siR-Control group and siR-ZFAS1 group, were collected, digested routinely by trypsin, as well as centrifuged and counted. Next, PBS buffer was used to wash cells, which were re-suspended in normal saline and adjusted for concentration to 5 × 106 cells/ml. Next, the single cell suspension was injected subcutaneously into the back of 14 nude mice by 200 μl/mouse, to establish transplanted tumor model of human cervical tumor in nude mice. When the tumor volume reached 0.1 cm3, 7 nude mice were randomly selected from each group with the intraperitoneal injection of 1 ml normal saline, while the other 7 mice were injected with cisplatin (3 mg/kg), once a week for four consecutive weeks. During the treatment, a Vernier calliper was used to measure the long and short diameters. The tumor volume was calculated according to the following formula: Tumor volume = tumor long diameter × tumor short diameter 2/2. Tumor growth curve was draw based on the tumor volume changes. After treatment, nude mice were sacrificed to take out their tumor mass for weight measuring.
Statistical methods
All data was statistically analyzed with the software SPSS 21.0 (SPSS, Inc, Chicago, IL, USA). The data in this study were presented by mean ± standard deviation. Comparison among multiple groups was performed by using One-Way ANOVA, while between two groups was analyzed by Student’s t-test. Kaplan-Meier methods with the log-rank test were performed to calculate overall survival. p<0.05 indicated the statistical significance.
Expression of ZFAS1 in cervical cancer tissues and cell lines
The expression levels of ZFAS1 in cervical cancer tissues and cells were detected by qRT-PCR. The results showed the ZFAS1 expression was apparently up-regulated in cervical cancer tissues as demonstrated in Figure 1A, and meanwhile, it was obviously increased in CaSki, HeLa and C33A cells when compared to HaCaT cells, which was significantly higher in HPV-positive cells (CaSki and HeLa) than the HPV-negative cell line C33A, and also higher in CaSki cells than HeLa cells (all p<0.05, Figure 1B). Thus, MTT assay was applied to observe the effects of different concentrations of cisplatin on the IC50 values of HeLa and CaSki cells Figure 1C. In comparison with HeLa cells, CaSki cells presented higher IC50 values, suggesting that CaSki cells had the significantly stronger drug resistance to cisplatin and ZFAS1 might be associated with cell sensitivity to cisplatin. Moreover, 21 cases out of the 85 patients died of cervical cancer according to the follow-up. Then, based on the median expression level of ZFAS1, cervical cancer patients were classified into high-expression group and low-expression group, and we found that cervical cancer patients with low-ZFAS1-expression had significantly higher survival rate than high-ZFAS1-expression ones (all p<0.05, Figure 1D).
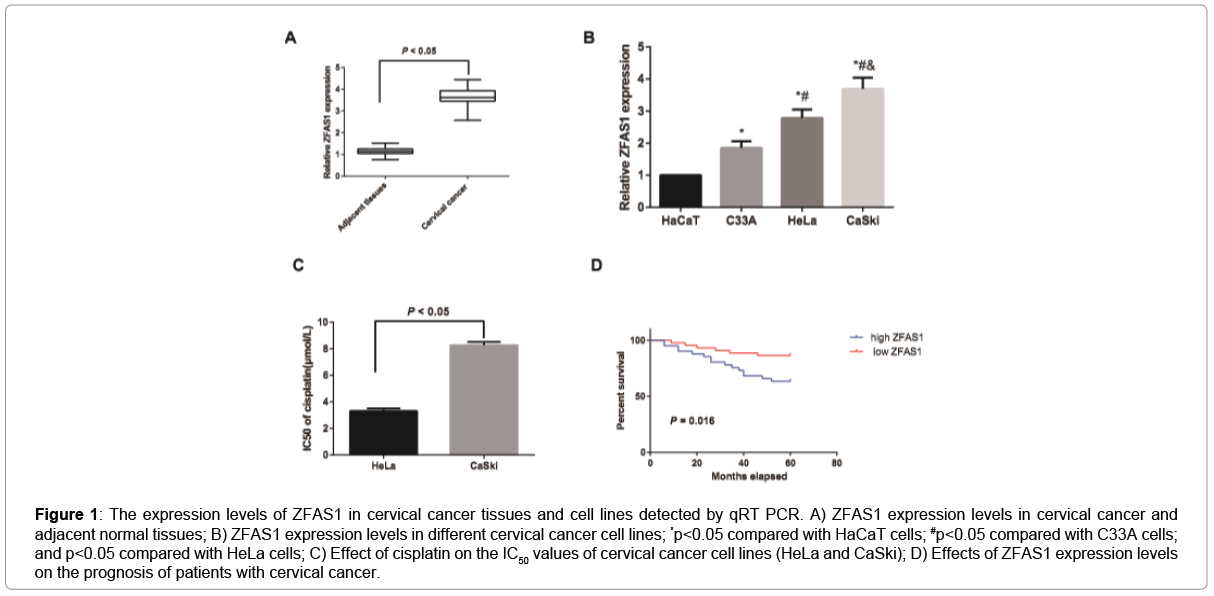
Figure 1: The expression levels of ZFAS1 in cervical cancer tissues and cell lines detected by qRT PCR. A) ZFAS1 expression levels in cervical cancer and adjacent normal tissues; B) ZFAS1 expression levels in different cervical cancer cell lines; *p<0.05 compared with HaCaT cells; #p<0.05 compared with C33A cells; and p<0.05 compared with HeLa cells; C) Effect of cisplatin on the IC50 values of cervical cancer cell lines (HeLa and CaSki); D) Effects of ZFAS1 expression levels on the prognosis of patients with cervical cancer.
Association of ZFAS1 with the clinicopathological characteristics of cervical cancer patients
As displayed in Table 1, ZFAS1 expression was not significantly related to the age and tumor diameter of cervical cancer patients (all p>0.05), but statistically associated with the FIGO staging, histological grading, lymph node metastasis, and depth of myometrial invasion in patients with cervical cancer (all p<0.05). In other words, patients with advanced FIGO stage, high histological grade, lymph node metastasis, and deep myometrial invasion had higher expression levels of ZFAS1 (all p<0.05).
| Clinicopathological characteristics | n | ZFAS1 expression | p-value |
|---|---|---|---|
| Age (Years) | |||
| < 45 | 48 | 3.65 ± 0.35 | 0.792 |
| ≥ 45 | 37 | 3.67 ± 0.34 | |
| Histological grading | |||
| G1 | 19 | 3.19 ± 0.18 | <0.001 |
| G2 | 35 | 3.57 ± 0.27 | |
| G3 | 31 | 4.02 ± 0.19 | |
| FIGO staging | |||
| I-II | 58 | 3.46 ± 0.24 | <0.001 |
| III-IV | 27 | 4.07 ± 0.18 | |
| Tumor diameter | |||
| ≤ 4 cm | 55 | 3.65 ± 0.36 | 0.609 |
| > 4 cm | 30 | 3.69 ± 0.31 | |
| Lymph node metastasis | |||
| Positive | 33 | 4.01 ± 0.20 | <0.001 |
| Negative | 52 | 3.42 ± 0.22 | |
| Depth of myometrial invasion | |||
| < 1/2 | 59 | 3.46 ± 0.23 | <0.001 |
| ≥ 1/2 | 26 | 4.08 ± 0.18 | |
Table 1: Association of ZFAS1 expression with clinico-pathological characteristics of patients with cervical cancer.
siR-ZFAS1 inhibits the proliferation of cervical cancer cells
According to the qRT-PCR detection Figure 2A, CaSki and HeLa cells in the siR-ZFAS1 group declined dramatically in the expression of ZFAS1 as compared with those in the Blank group and siR-Control group (all p<0.05). Cell proliferation was detected by MTT assay and the results showed that the proliferation of CaSki and HeLa cells in the siR-ZFAS1 group were significantly poorer than those in the Blank and siR-Control groups (all p<0.05, Figure 2B). However, there was no observable difference between the Blank group and the siR-Control group in ZFAS1 expression levels and cell proliferation (all p>0.05). Flow cytometric cell cycle assays in these cell lines demonstrated that ZFAS1 knockdown led to a significant cell cycle arrest at G0/G1-phase and a significant decrease in cells in S-phase (all p<0.05, Figure 2C).
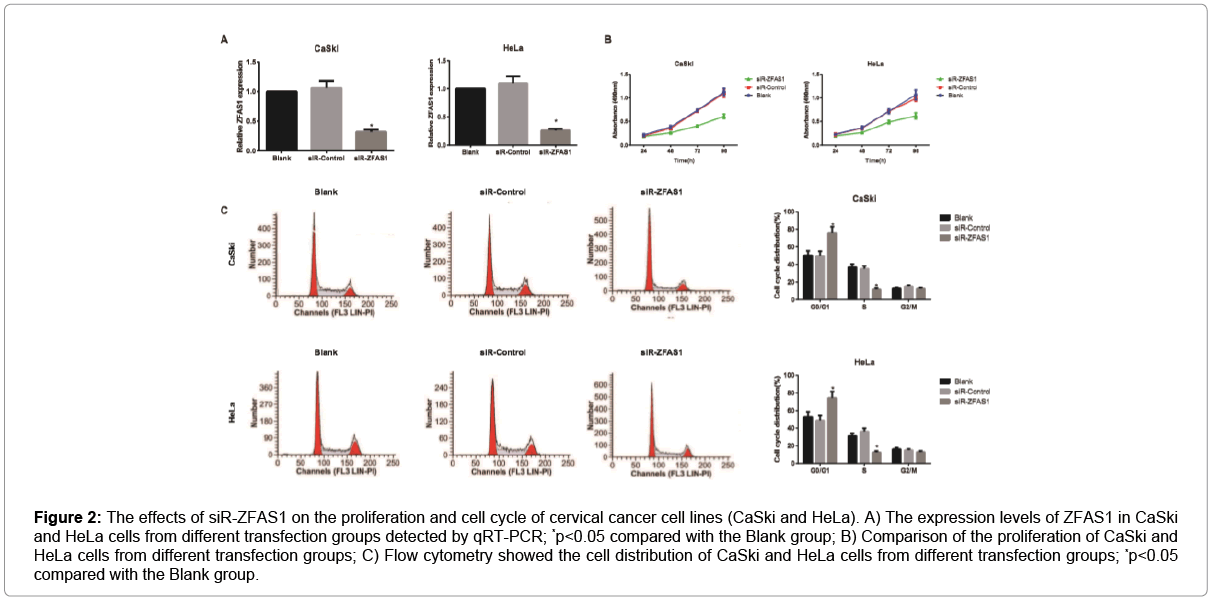
Figure 2: The effects of siR-ZFAS1 on the proliferation and cell cycle of cervical cancer cell lines (CaSki and HeLa). A) The expression levels of ZFAS1 in CaSki and HeLa cells from different transfection groups detected by qRT-PCR; *p<0.05 compared with the Blank group; B) Comparison of the proliferation of CaSki and HeLa cells from different transfection groups; C) Flow cytometry showed the cell distribution of CaSki and HeLa cells from different transfection groups; *p<0.05 compared with the Blank group.
siR-ZFAS1 inhibits the invasion and migration of cervical cancer cells
As shown in Figure 3, cell migration and invasion abilities were evaluated by wound-healing and Transwell assays. Obviously, CaSki and HeLa cells in the siR-ZFAS1 group decreased dramatically in the migration rate and the number of invasive cells, as compared with those in the Blank group and the siR-Control group (all p<0.05). However, Blank group and siR-Control group had no significant changes regarding the above factors (all p>0.05).
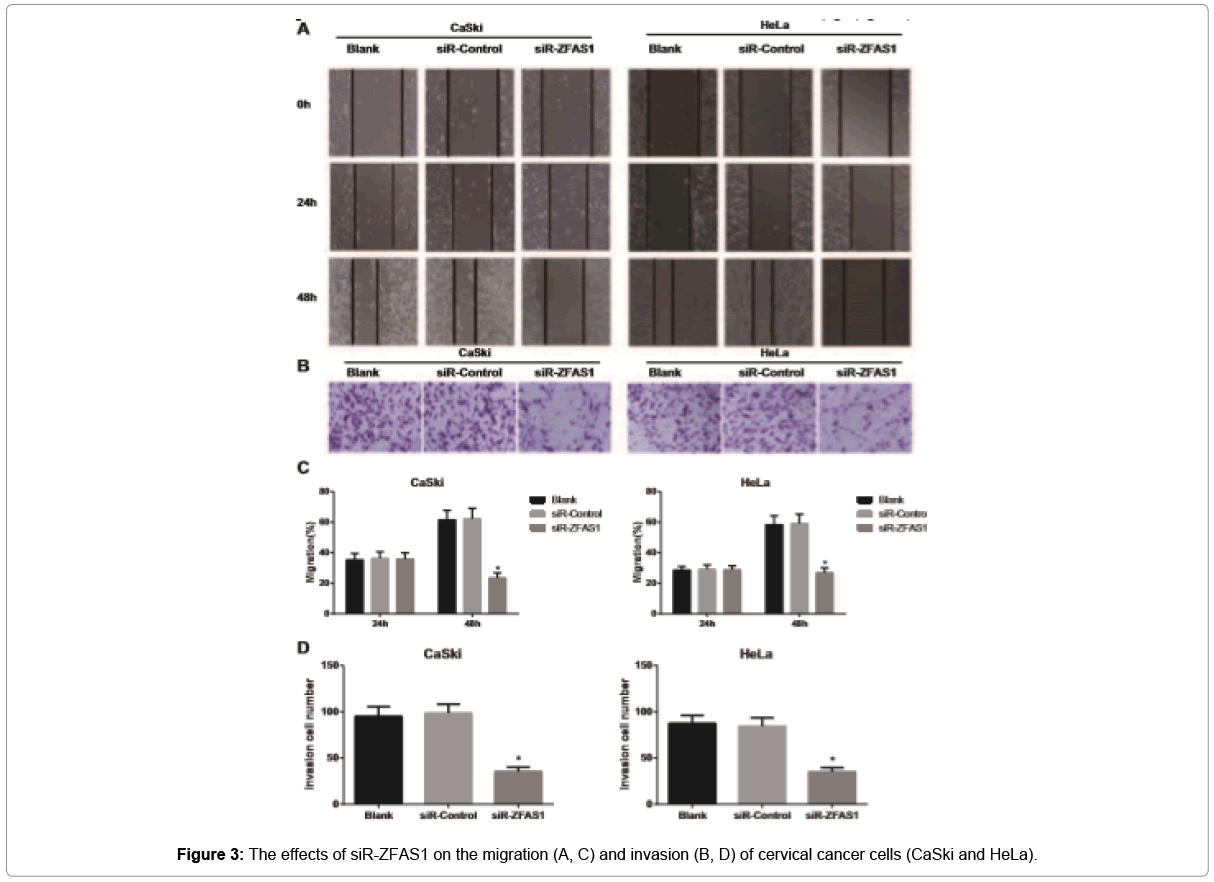
Figure 3: The effects of siR-ZFAS1 on the migration (A, C) and invasion (B, D) of cervical cancer cells (CaSki and HeLa).
siR-ZFAS1 enhances chemosensitivity to cisplatin in cervical cancer cells
The growth inhibition rate of CaSki and HeLa cells in the siRZFAS1 group was apparently elevated, as compared with Blank group and siR-Control group (both p<0.05, Figure 4), but no statistical difference was observed between the Blank group and the siR-Control group (p>0.05). Besides, the IC50 values of cisplatin in CaSki and HeLa cells from the siR-ZFAS1 group were appreciably lower than that from the Blank group and siR-Control group (all p<0.05), which had no obvious difference between Blank group and siR-Control group (p>0.05), suggesting that siR-ZFAS1 may effectively improve the chemosensitivity of CaSki and HeLa cells to cisplatin.
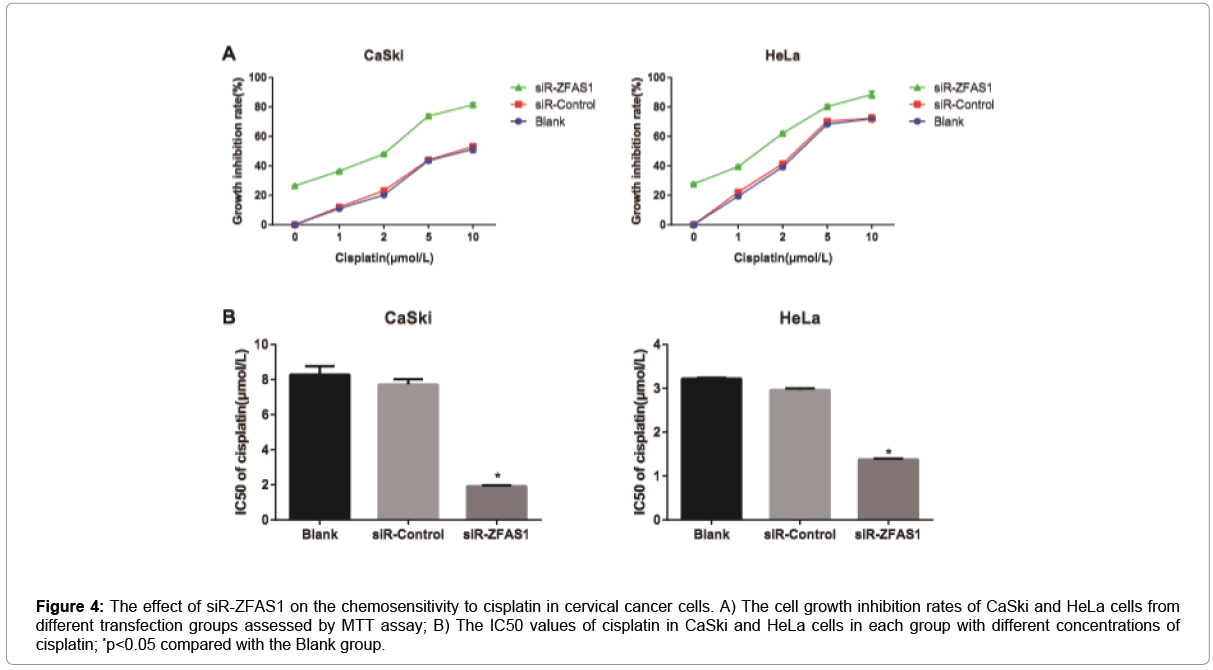
Figure 4: The effect of siR-ZFAS1 on the chemosensitivity to cisplatin in cervical cancer cells. A) The cell growth inhibition rates of CaSki and HeLa cells from different transfection groups assessed by MTT assay; B) The IC50 values of cisplatin in CaSki and HeLa cells in each group with different concentrations of cisplatin; *p<0.05 compared with the Blank group.
siR-ZFAS1 increases the apoptosis rate of cisplatin-induced cervical cancer cells
The apoptosis rate of cells in different groups with different concentrations of cisplatin was determined by flow cytometry. According to the results, the apoptosis rate of CaSki and HeLa cells in each group was elevated accordingly with the increase of cisplatin concentration. Besides, siR-ZFAS1 group had the increased apoptosis rate of CaSki and HeLa cells when compared to the Blank group and the siR-Control group (all p<0.05), but the Blank group and the siRControl group didn’t have significant difference from each other concerning the cell apoptosis rate (p>0.05, Figure 5). Furthermore, we treated CaSki and HeLa cells in different transfection groups with 10μmol/L of cisplatin and then followed by western blotting analysis. The results showed that expression levels of cleaved PARP and cleaved caspase-3 were increased in both cell lines after ZFAS1 silencing, indicating higher apoptosis as a consequence of ZFAS1 silencing (all p<0.05, Figure 6).
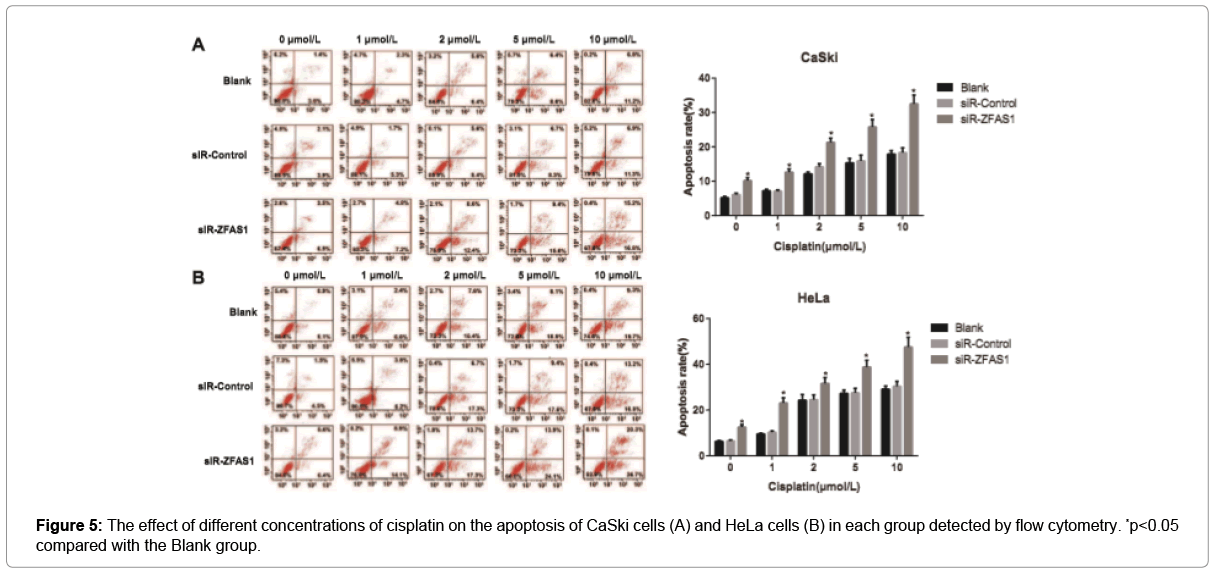
Figure 5: The effect of different concentrations of cisplatin on the apoptosis of CaSki cells (A) and HeLa cells (B) in each group detected by flow cytometry. *p<0.05 compared with the Blank group.
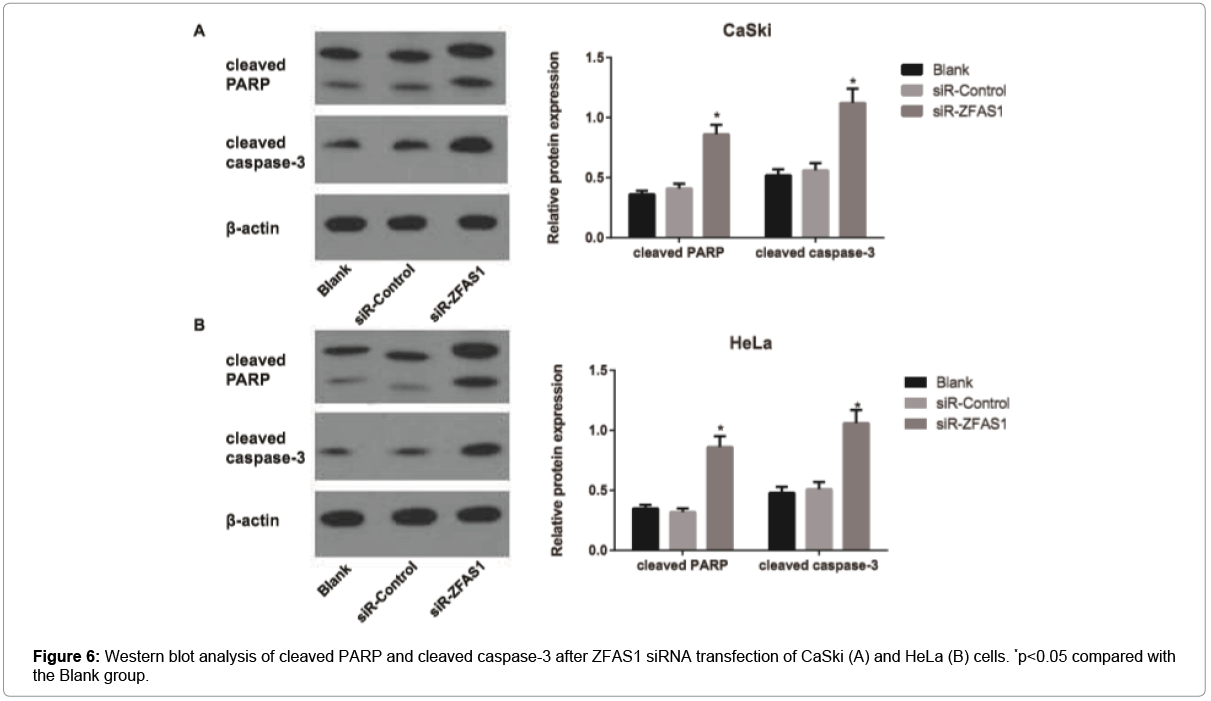
Figure 6: Western blot analysis of cleaved PARP and cleaved caspase-3 after ZFAS1 siRNA transfection of CaSki (A) and HeLa (B) cells. *p<0.05 compared with the Blank group.
siR-ZFAS1 suppressed subcutaneous tumor growth in response to cisplatin treatment in vivo
The tumor volume and weight were reduced in nude mice from the siR-ZFAS1 group, as compared with those from the Blank and siRControl groups (all p<0.05, Figure 7), but the tumor volume and weight in nude mice from Blank group was not differ from the siR-Control group (both p>0.05). After treated with cisplatin, nude mice in the siRZFAS1 + cisplatin group exhibited lower tumor volume and weight than those in the Blank + cisplatin group and the siR-Control + cisplatin group (all p<0.05), and there was no apparent difference between the Blank + cisplatin group and the siR-Control + cisplatin group (all p>0.05), indicating that siR-ZFAS1 can improve the chemosensitivity of tumor cells to cisplatin in nude mice with transplanted model of human cervical cancer cells.
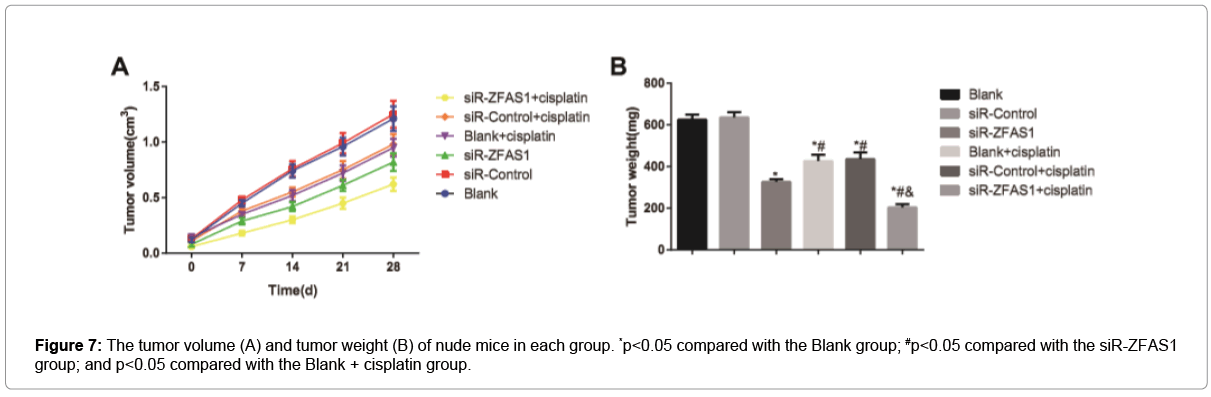
Figure 7: The tumor volume (A) and tumor weight (B) of nude mice in each group. *p<0.05 compared with the Blank group; #p<0.05 compared with the siR-ZFAS1 group; and p<0.05 compared with the Blank + cisplatin group.
In our study, the expression levels of ZFAS1 were found to be significantly up-regulated in both cervical cancer tissues and cell lines. Meanwhile, its expression was closely associated with the FIGO staging, histological grading, lymph node metastasis, as well as depth of myometrial invasive in cervical cancer, and patients with high-ZFAS1- expression predicted a poor prognosis, showing that ZFAS1 may act as an oncogene to affect the occurrence, development and prognosis of cervical cancer. In agreement with our findings, Bairong Xia et al. [10] identified ZFAS1 was also increased in ovarian cancer tissues and had a close relation with the major clinicopathological characteristics, like clinical stage and lymph node metastasis, as well as prognosis of patients [10]. Also, the dysregulated ZFAS1 was found to have an oncogenic role in other various types of human malignant tumors in numerous previous studies, such as bladder cancer and gastric cancer [12,21]. Besides, patients with higher expression levels of ZFAS1 predicted an unfavorable overall survival in glioma and non-small cell lung cancer (NSCLC), suggesting that ZFAS1 could function as a potential prognostic indicator and promising therapeutic target for certain cancers [22,23]. Worth mentioning, SP1 (specificity protein 1) was found to be an upstream activated factor of ZFAS1, which could bind to the promoter of ZFAS1 to activate its transcription in the study of Liu et al. [24]. There was evidence pointed out that SP1, as a sequencespecific DNA-binding protein, could initiate the transcription of many cellular genes, participating in a wide range of biological processes, like cell proliferation, differentiation and tumor formation [25]. More importantly, SP1 was discovered to be overexpressed in cervical cancer and involved in the development and progression of cervical cancer [26,27]. Thus, we hypothesized that the high expression of ZFAS1 in cervical cancer might be induced owing to the activation of its upstream transcription factor SP1.
To further investigate the role of ZFAS1 in cervical cancer, the ZFAS1 siRNA were transfected in CaSki and HeLa cells, and we found the proliferation, invasion and migration of cervical cancer cells were inhibited after silencing ZFAS1. Consistently, knockdown of ZFAS1 could reduce the expression of ZEB1 in colon cancer to inhibit cell proliferation and invasion and promote cell apoptosis, as indicated by Fang et al. [14]. Besides, ZFAS1 has been thought to play as a competing endogenous RNA (ceRNA) to regulate miRNA activity and modulate the expression of target genes, thereby playing an essential part in the occurrence and progression of tumors [28-30]. For example, the in vitro and in vivo experiments confirmed that ZFAS1 knockdown could suppress osteosarcoma growth and progression via sponging miR-486 [31], but on the contrary, ZFAS1 could increase the growth and metastasis of osteosarcoma cells via competitively binding to miR- 200b and miR-200c to up-regulate BMI1 expression and modulate ZEB2 protein stability [24] To our knowledge, a variety of lncRNAs, like HOTAIR [32] H19 [33] and PVT1 [34], have been reported to participate in the occurrence and development of cervical cancer by regulating the expression of downstream miRNAs. Therefore, we hypothesized that ZFAS1 may act as a ceRNA to regulate the expression of downstream pathway-related proteins, and thereby affecting the proliferation, invasion and migration of cells, but further study is needed to elucidate the specific mechanism.
In addition, we also found ZFAS1 expression was relatively higher in CaSki cells which had relatively low cisplatin sensitivity. Coincidentally, Liu and the colleague also reported that highly-expressed ZFAS1 in ovarian cancer cells indicated the poor chemosensitivity to cisplatin [35], suggesting that ZFAS1 may play a vital role in affecting the chemosensitivity of cervical cancer cells to cisplatin. In our study, after the cervical cancer cells treated with different concentrations of cisplatin, we observed that the knockdown of ZFAS1 can obviously enhance the cell growth inhibition rate and decrease the IC50 values, and elevate cell apoptosis, implying that inhibiting ZFAS1 can improve the chemosensitivity of cervical cancer cells to cisplatin. In the study of Bairong Xia et al. [10], inhibiting ZFAS1 expression can down-regulate SP1 expression by modulating miR-150-5p, thereby inhibiting the proliferation and migration of ovarian cancer cells and improving cells’ chemosensitivity to cisplatin [10]. While Vizcaíno et al. [25] also proved that SP1 can become the potential target of cancer chemotherapy [25]. Actually, it has been reported that cisplatin can bind to DNA to form DNA complex and activate the DNA damage response (DDR), and eventually killing the tumor cells [36,37]. However, Olofsson et al. [38] found that SP1 can promote the phosphorylation of SQ/TQ cluster domains in DNA damage regions to alleviate DNA damage and reduce tumor cell apoptosis. Furthermore, SP1 can directly regulate the promoter of cell apoptosis-related proteins to influence the apoptosis of tumor cells [39]. Taken together, down-regulation of ZFAS1 in cervical cancer cells can inhibit SP1 levels by regulating the downstream miRNAs, to increase cisplatin-induced DNA damage and promote cell apoptosis, ultimately improving the chemosensitivity to cisplatin.
Finally, through experiments in nude mice subcutaneously transplanted with human cervical cancer cells, we found that siRZFAS1 can improve the sensitivity of transplanted tumor cells to cisplatin chemotherapy, which further confirmed the above results in cell experiments. Collectively, ZFAS1 was found highly expressed in cervical cancer in this study, and patients with high ZFAS1 expression predicted a poorer prognosis. Besides, silencing ZFAS1 may inhibit the proliferation, migration and invasion of cervical cancer cells, and improve the chemosensitivity to cisplatin, which could offer a new perspective for the clinical chemotherapy of cervical cancer.
The authors would like to thank the reviewers for their constructive comments.
There were no competing interests.