Journal of Horticulture
Open Access
ISSN: 2376-0354
ISSN: 2376-0354
Short Communication - (2016) Volume 3, Issue 3
Florida hybrid (Vitis spp.) and muscadine (V. rotundifolia Michx.) grape acreage is increasing in the south-eastern United States for the fresh market, wine production, and other diversified products. There is little research on determining leaf area in Muscadine grapes that could be used to assess canopy balance. Previous research on nondestructive methods of assessing canopy leaf area in V. vinifera L. and V. labrusca L. varieties may not apply to V. rotundifolia and Florida hybrid varieties. Thus this research sought to verify the accuracy previous non-destructive methods for V. rotundifolia and Vitis spp. varieties grown in Florida. Linear measurements were taken on fully expanded leaf of ‘Blanc Du Bois’ (Vitis spp.), ‘Carlos’ (V. rotundifolia Michx.) and ‘Southern Home’ (interspecific V. rotundifolia × V. vinifera hybrid). Maximum length and width were used to predict leaf area using linear regressions. For each cultivar, 5 linear regression equations were developed to estimate leaf area using length and width. Length and width were log transformed and an additional 3 linear equations were developed. Equations with the product of length and width had a higher coefficient of determination for all three varieties. Therefore, these equations are recommended for use when conducting non-destructive leaf area measurements of muscadine and Florida hybrid grape varieties.
<Keywords: Canopy size; Leaf length; Leaf size; Leaf width; Models; Non-destructive leaf; Measurement
Muscadine varieties and hybrids such as ‘Blanc Du Bois’ are cultivated in the South-eastern United States for their resistance to Pierce’s Disease, caused by Xylella fastidiosa [1,2]. The fruit and extracted oils of muscadine grapes are high in vitamins, fiber, amino acids, and antioxidants that help to inhibit heart disease, prevent obesity-related health complications and certain cancers [3-5]. Therefore, muscadine production acreage is expected to expand as demand for these beneficial compounds increase.
Canopy size and volume have direct effects on the vine microclimate by influencing temperature and light interception within the canopy, with larger canopies reducing light penetration and air movement into and throughout the fruit zone. Therefore an adequate canopy size with sufficient light exposure is considered a balanced canopy. Moreover vines with equal vegetative and reproductive growth are considered a well-balanced vine. Light interception and the balance between vegetative growth and yield can affect accumulation of phenolic compounds such as flavonoids and anthocyanins [6,7], important antioxidants being sought for beneficial health attributes. Canopy parameters such as leaf density, distribution, and leaf area can be quantified to measure the light interception efficiency [8,9]. In addition, light penetration leads to thicker leaves and higher percentages of budbreak and bud fruitfulness, resulting in higher yield [10-12].
Previous research in V. vinifera suggests that 16 leaves/shoot are needed in order to adequately ripen a single cluster [13]; however, this differs with cultivar, training system and vineyard location. Shoot and leaf orientation directly affect bunch exposure with downward-facing shoots largely improving grape chemical components [11,14,15]. In the case of muscadine its procumbent shoot growth could make the leaves closer to the fruit a more important source of sugars, thus directly affecting fruit quality. This could be explained by the fact that the lights received by leaves located in specific parts of the plant contribute more to the photosynthetic rate of the plant than the total amount of light intercepted by the canopy [16]. Sun-exposed leaves have higher photosynthetic rates, producing more carbohydrates that are allocated to fruit with increased sugars and phenolic compounds [13,15,17,18]. Therefore, information on leaf area and distribution within the canopy can determine whether the vine is well-balanced. Cultural practice management can then be adjusted to include such practices as shoot thinning and hedging.
Direct methods of leaf area measurement may be destructive or non-destructive. Among the direct non-destructive methods are: automatic planimeters, leaf area from direct leaf dimension measurements, and leaf area to weight ratios [8,9,19-21]. Indirect methods using radiation techniques are expensive and require proper training; therefore, the use of linear measurements to nondestructively determine leaf area is desirable.
Non-destructive models have been developed for ‘Concord’ (V. labrusca L. ), ‘Chardonnay’ (V. vinifera L. ), ‘Chenin blanc’ (V. vinifera L. ), ‘Cencibel’ (V. vinifera L. ), ‘Thompson Seedless’ (V. vinifera L. ), ‘DeChaunac’ (Vitis spp. ), and ‘Niagara’ (V. labrusca L. ) [22-26]. Leaf shape and area for each of the V. vinifera cultivars are similar; however, there is no information on the applicability of these models for muscadine or Florida hybrid wine grapes, some of which have unique leaf shapes (Figure 1).

Figure 1: Differing leaf shapes of ‘Blanc Du Bois’ (Vitis spp.), ‘Carlos’ (V. rotundifolia Michx.) and ‘Southern Home’ (interspecific V. rotundifolia × V. vinifera hybrid).
Muscadine and Florida hybrid grape acreage is expected to expand because of interest in the production of healthy compounds and improved disease resistance. Thus, it will be beneficial to be able to determine leaf area by non-destructive methods to manage canopy size to obtain good fruit quality. In addition this technique could be used by grape physiologists studying the effect of salinity or drought on grape growth. Therefore, the objective of this work was to determine the ideal regression equation for non-destructive linear measurements to predict leaf area in southern U.S. grape varieties, particularly V. rotundifolia and Vitis spp.
Grape leaves were collected from five-year-old ‘Blanc Du Bois’ vines, located in Clermont, FL (28.5° lat., 81.7° long.), while ‘Carlos’ and ‘Southern Home’ vines were located in Gainesville, FL (29.6° lat., 82.3° long). ‘Blanc Du Bois’ vines were planted in rows oriented northsouth with 7 m between rows and 2 m between vines. ‘Blanc Du Bois’ vines were drip-irrigated and spur-pruned to 80 buds per vine and trained to a bilateral cordon 91 cm above the ground. ‘Carlos’ and ‘Southern Home’ vines were trained on a unilateral cordon 1.5 m above the ground. ‘Blanc Du Bois’ shoots were collected on 5 May 2013 and 21 May 2014, while ‘Carlos’ and ‘Southern Home’ were collected on 22 May 2014 (n=18). Shoots were collected from 10 plants at phenological stage 19 according to the modified Eichhorn-Lorenz scale [27]. For each sampling period, collected shoots were kept in a cooler with ice and transported to the laboratory in Gainesville, FL for leaf area measurements. The maximum width and the maximum length of each fully expanded leaf were measured and recorded to the nearest 0.5 cm [22]. Each leaf was then scanned using a leaf area meter (LI-3100C, LICOR, Lincoln, NE) and leaf area (cm2) recorded.
Maximum leaf length and width measurements were used to fit linear regression models using length, width and leaf area (Microsoft Excel 2013, Microsoft Corp., Redmond, WA). The data obtained for each measurement was transformed to the natural logarithm for some regression models when appropriate. Non-destructive leaf area equations were selected based on the best coefficient of determination (R2 >0.95). Thus, the higher the R2, the higher the accuracy of the equation. In each regression equation, p-values smaller than 0.05 indicated significant differences.
The products of length and width were better predictors of leaf area for all three varieties particularly when the data were transformed prior to analysis (Table 1). In this study, when only one of the linear measurements was used, width rather than length appeared to have a better fit for prediction leaf area for three varieties examined with exception of equation 5 for ‘Carlos’ (Table 1).
| Variety / Eq. number | Regression equations | R2 |
|---|---|---|
| ‘Blanc Du Bois’ | ||
| 1 | A=8.95L-24.472 | 0.847 |
| 2 | A=9.223805W-27.5463 | 0.865 |
| 3 | A=0.6296LW+1.6178 | 0.922 |
| 4 | A=0.605692L2+3.214566 | 0.884 |
| 5 | A=0.609868W2+2.089168 | 0.904 |
| 6 | A=0.87950(L)1.8563 | 0.916 |
| 7 | A=0.682085(W)1.965366 | 0.927 |
| 8 | A=0.732743(LW)0.970638 | 0.936 |
| ‘Carlos’ | ||
| 1 | A=6.4395L-15.098 | 0.918 |
| 2 | A=7.6343W-17.68 | 0.919 |
| 3 | A=0.6323LW+0.17525 | 0.970 |
| 4 | A=0.5287L2+1.719 | 0.937 |
| 5 | A=0.7009W2+0.4545 | 0.936 |
| 6 | A=0.641657(L)1.93319 | 0.955 |
| 7 | A=0.6109(W)2.0776 | 0.976 |
| 8 | A=0.59073(LW)1.0818 | 0.981 |
| ‘Southern Home’ | ||
| 1 | A=7.1365L-19.665 | 0.900 |
| 2 | A=7.3884W-19.868 | 0.915 |
| 3 | A=0.5542LW-0.4293 | 0.951 |
| 4 | A=0.5279L2+0.1335 | 0.925 |
| 5 | A=0.5585W2+0.2974 | 0.941 |
| 6 | A=0.40828(L)2.1138 | 0.957 |
| 7 | A=0.45018(W)2.0989 | 0.968 |
| 8 | A=0.4155(LW)1.0612 | 0.970 |
Table 1: Linear regression equations and coefficients of determination (R2) for ‘Blanc Du Bois’ (Vitis spp.), ‘Carlos’ (V. rotundifolia Michx.), and ‘Southern Home’ (interspecific V. rotundifolia× V. vinifera) hybrid leaf area (A) using two measurements, length (L) or width (W).
The coefficient of determination for leaf length and/or width as a predictor of leaf area was lowest for ‘Blanc Du Bois’ compared with ‘Southern Home’ and ‘Carlos’; however, data transformation improved the R2. In the case of ‘Blanc Du Bois’, the use of length as a predictor resulted in the lowest R2 value. For ‘Carlos’, an increase in coefficient of determination was observed when width and length were transformed to natural log (Table 1, Eq. 8). A similar result was obtained for ‘Southern Home’ with improvement in the R2 by 0.053 when data were log transformed (Table 1). Thus, transforming the data allows the variables (length and width) to be better predictors of leaf area, since R2 values were closer to one in equations where data were transformed. The increased accuracy when data were transformed is represented in Figures 2-4 where equations of the product of length and width were graphed and compared with the log transformation of the variables for each of the varieties sampled.
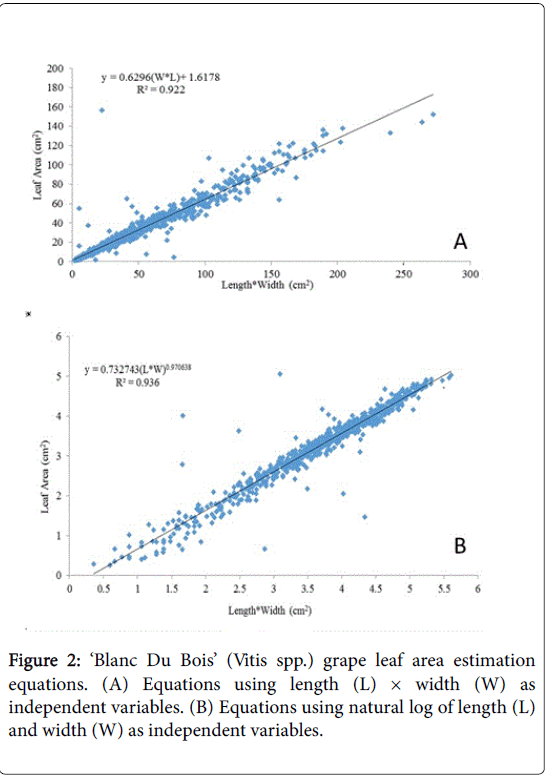
Figure 2: ‘Blanc Du Bois’ (Vitis spp.) grape leaf area estimation equations. (A) Equations using length (L) × width (W) as independent variables. (B) Equations using natural log of length (L) and width (W) as independent variables.
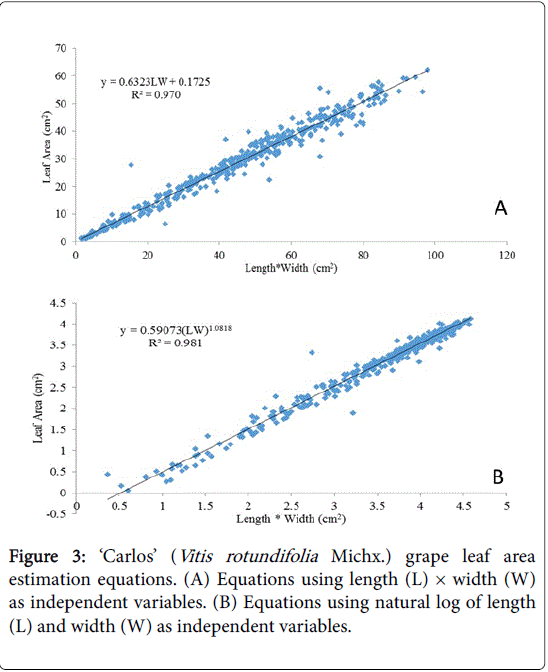
Figure 3: ‘Carlos’ (Vitis rotundifolia Michx.) grape leaf area estimation equations. (A) Equations using length (L) × width (W) as independent variables. (B) Equations using natural log of length (L) and width (W) as independent variables.
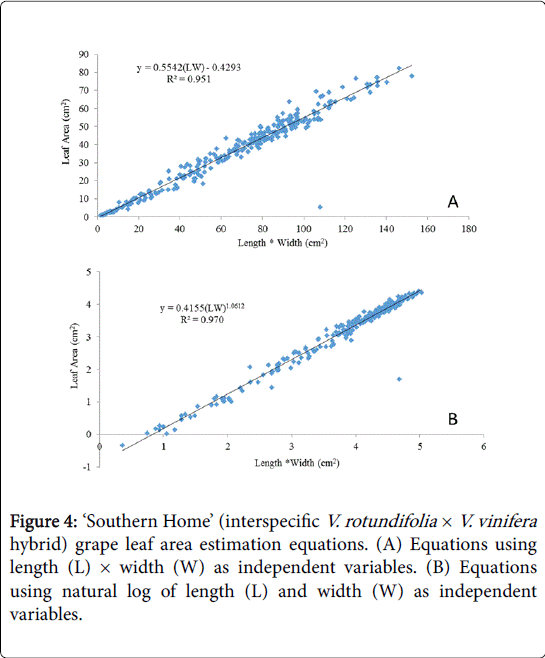
Figure 4 : ‘Southern Home’ (interspecific V. rotundifolia × V. vinifera hybrid) grape leaf area estimation equations. (A) Equations using length (L) × width (W) as independent variables. (B) Equations using natural log of length (L) and width (W) as independent variables.
Leaf area of ‘Blanc Du Bois’ and ‘Southern Home’ can be predicted by using a simple linear measurement such as width. Length itself was not a good predictor of leaf area in ‘Blanc Du Bois’. However, the product of length and width is a better predictor as demonstrated in other grape species such V. vinifera and V. labrusca [22,23,25,27] verifying the accuracy of these models for the grape species examined in this study. Similar results have been found for ‘DeChaunac’ and ‘Niagara’ grapes [26,28].
Conversely in ‘Carlos’, the R2 values of the prediction equations were similar when either length or width were used (Table 1). This may be because ‘Carlos’ has a minimally lobed, symmetrical leaf shape, as is the case with a majority of the muscadine grape varieties [3]. In ‘Southern Home’, which has a deeply lobed leaf (Figure 1), a single leaf parameter was less precise because of these lobes and deep dentate margins. This may explain the decrease in R2 value when length as a single measurement was used (Table 1).
However, when the data were displayed graphically, an increase in the coefficient of determination was not observed when data were transformed. In addition, a very strong relationship (R2>0.60) was observed without data being log transformed (Figures 2-4). Therefore, the necessity to transform the data to have a better predicting equation will depend on the accuracy required for its use. Moreover, it was observed in this experiment that width by itself could be used as the only predictor for the three varieties in order to save time and simplify data collection.
Non-destructive leaf area for ‘Blanc Du Bois’, ‘Carlos’ and ‘Southern Home’ could be predicted using length and width. However, the product of these variables best predicted the leaf area of ‘Blanc Du Bois’ (Florida hybrid; Vitis spp.) A=0.62956W*L+1.61775(R2=0.922), A=0.732743(LW) 0.970638 (R2=0.936), ‘Carlos’ (muscadine) A=0.6323LW+0.17525 (R2=0.970), A=0.59073(LW) 1.0818 (R2=0.981) and ‘Southern Home’ (interspecific V. rotundifolia × V. vinifera hybrid) A=0.5542(LW)-0.4293 (R2=0.951), A=0.4155(LW) 1.0612(R2=0.970). In this study, the product of width and length was found to be a better predictor of leaf area than either variable alone, consistent with previous reports.
Funding for this project by the Florida Viticulture Advisory Council is gratefully acknowledged. The authors wish to thank Matthew Ross, Dr. Richard Beeson, Jim Sievert and Javier Lopez for their technical assistance.