Journal of Horticulture
Open Access
ISSN: 2376-0354
ISSN: 2376-0354
Research Article - (2015) Volume 2, Issue 4
Pot plant production in green house is most of the time under high relative humidity and frequent irrigation. While, during shipping and retailing plants may be exposed to high temperature and infrequent irrigation. These unfavorable conditions often cause water loss, desiccation of plants and short shelf life. To reduce the deteriorating effect of water loss, application of anti-transpirants is one of the integral options to implement. In this study we evaluated several physiologically active stomata closing compounds. Spathiphyllum sweet checo and Calathea rufibarba were sprayed with different active compound formulations. Weight loss of leaves as gram of water per gram of initial leaf weight and gram of water per centimeter square leaf area and stomatal conductance were measured. On both plant species most physiologically active compound treatments reduce weight loss and stomatal conductance significantly. 1.5 mM Abscisic acid (ABA), 5 mM Salicylic acid (SA) and 200 µM Sodium nitroprusside (SNP) on Spathiphyllum sweet checo and 1.5 mM ABA ,200 µM SNP and 200 µM Pyrabactin on Calathea rufibarba were the three most effective active compound treatments. Significant positive correlation were found between weight loss and stomatal conductance on both plant species (R=0.888, P<0.05), (R=0.811, P<0.05) for Spathiphyllum and Calathea respectively. The relative efficacy of different active compound treatments and the effects of leaf weight and leaf area on water loss were also inseparable in both plant species.
<Keywords: Leaf Shine; Abscisic Acid; Salicylic Acid; Sodium Nitroprusside; Pyrabactin.
Pot plant production in green house is most of the time under high relative humidity and frequent irrigation. While, during shipping and retailing plants may be exposed to high temperature and infrequent watering. These unfavorable post production environments can cause rapid substrate drying, desiccation of plants and acceleration of senescence and then short shelf life [1].Water loss is the main post production problem that deteriorate the quality of potted plants and make them unsalable [2]. For example, in co mMon potted foliage plants Monstera deliciosa and Philodendron sp the amount of water loss during shipping and retailing was much higher than their growing environment leading to premature leaf wilting and death [3].
The main cause of water loss from plants is because of transpiration through the stomatal opening. The opening of stomata allows gas exchange between the plant and its environment. In the course of stomatal opening CO2 can enter into the plant and water can be lost as a vapor. Hence, to avoid desiccation and eventual senescence, it is vital to have a right balance between carbon gain and water loss through stomatal movements [4]. A stoma (plural stomata) is a microscopic pore on the surface (epidermis) of land plants. It is surrounded by a pair of specialized epidermal cells called guard cells, which act as a turgor-driven valve that open and close its pores in response to a given environmental conditions. The opening and closing mechanism of stomata depends upon the turgor pressure in the guard cells. When the guard cells are turgid, the stoma opens and when the guard cells loose water, stoma closes. Stomatal closure can be induced by many factors, such as osmotic stress, darkness, high CO2 concentration and mechanical stresses [5]. Naturally, plants react to water deficit by closing stomata mainly mediated by Abscisic acid (ABA) During water stress ABA triggers the closure of stomata, consequently a reduction in water loss by transpiration [6]. In the course of ABA induced stomatal closure, first ABA binds to its receptors at the surface of the plasma membrane of the guard cells. The receptors in the guard cells can then activate several interconnecting pathways that converge to produce: Cytosolic PH rise and transfer of Ca2+ from the vacuole and intercellular spaces to the cytosols of the guard cells. The changes in cytosol pH and transfer of Ca2+ can stimulate the loss of negatively-charged ions (anions), especially NO3- and Cl- from the guard cells. The loss of these anions can lead to the depolarization of guard cells membrane. The depolarization of the plasma membrane can trigger the loss of K+ from guard cells. The loss of these solutes in the cytosol then reduces the osmotic pressure of the cell and thus turgor [7,8].
To reduce water loss and its subsequent negative effect on the qualities of pot plant, it is important to develop techniques that can be engaged to reduce transpiration and preserving postharvest quality for extended time [9]. The major techniques that can be used to reduce transpiration could be the use of anti-transpirant agents. Anti-transpirant agents are compounds or provisions capable of reducing water loss from leaves of plants. These agents can be grouped into three groups [10], i) Film-forming types: that can create a physical barrier between the leaf and the surrounding (e.g. glycerol) ii) Reflecting materials: that can reflect the radiation falling on the surface of the leaves to reduce leaf temperature and the light needed for signaling during stomatal opening (e.g. Kaolin), and iii) Physiologically active stomata closing types: such as ABA and NO, ethylene etc. capable of affecting metabolic processes in leaf tissues.
All things considered, the interest of this study was the evaluation of physiologically active stomata closing compounds for their anti-transpirant effect on two pot plant species and to compare the extent at which these active compounds control transpiration relative to the successful co mMercially available film forming anti-transpirant Leaf shine (LS) Demonstrated by many studies, physiologically active compounds instituted to be effective in reducing stomatal apertures in different plant species and different conditions are Abscisic acid (ABA) Aminoethoxyvinylglycine (AVG), Ethylene, 1-methylcyclopropene (1-MCP), Nitrous oxide (NO), Hydrogen peroxide (H2O2), Pyrabactin, Salicylic acid (SA) and Methyl Jasmonate (MJ) etc.
Excised leaves of Spathiphyllum sweet Checo and Calathea rufibarba were used. Leaves were cut from their mother plant using sharp scissor leaving a 10 cm petiole intact with the leaf and 30 ml of tap water filled in a plastic tube enclosed by a rubber cap was used as a vase solution. Matrix solution (producer unknown) as carrier solution for active compounds, film forming anti-transpirant: Chrysal Leaf shine concentrate solution diluted 1:20 with deionised water (Chrysal international B.V,Naarden ,the Netherlands), active compounds: ABA 10% w/v (Valent USA, Walnut Creek, CA, united states of America)Hydrogen peroxide 30.wt.sol.in water, Sodium nitroprusside dehydrate, Salicylic acid sodium salt (2 hydroxy benzoic acid), Methyl jasmonate 95% and (CaCl2) Calcium chloride dehydrate (all from Sigma-Aldrich Corporation, Steinheim Germany ) were used. Spraying was done under the flow hood using a very fine airbrush sprayer powered by pressurized gas. The abaxial and adaxial surfaces of excised leaves were sprayed homogenously up until the spraying solution started to flood off the leaf surface. After spraying was done leaves were left to dry outside the flow hood in the laboratory for about two hrs. After leaves were completely dried they were transferred to the flow hood under continuous light exposure. Temperature in the flow hood was revolving between 20 to 22°C and relative humidity was 45 to 50 throughout the 10 day experimental period.
At the start of each experiment initial leaf weight of each leaf was determined. Once leaves were sprayed and dried, the weight of the leaf+tube+vase solution were measured and recorded as a starting point. In the course of the experimental period weight loss from the system was measured and recorded every 24 hr. At the last day of each experiment leaf area was also measured using LI-3100C Area Meter; before the actual leaf area measurement, the area meter was turned on for 15 minutes and calibrated. Daily water loss from the leaf surface was calculated using a simple formula: Daily water loss=(Weight of the leaf+tube+vase solution at day 0, 1, 2…10–weight of leaf+tube+vase solution at day 1, 2, 3….10)The amount of water transpired from the leaf surface each day was then calculated both in terms of amount of water transpired per grams of initial leaf weight and per centimeter square leaf area using the formula:
Transpiration 1=((Weight of the leaf+tube+vase solution at day 0, 1, 2…(n)–weight of leaf+tube +vase solution at day 1, 2, 3….(n) / initial leaf weight)) and Transpiration 2=((Weight of the leaf+tube+vase solution at day 0, 1, 2…(n)–weight of leaf+tube+vase solution at day 1, 2, 3….(n)/ leaf area))
Cumulative water loss over time was also calculated by su mMing up weight loss at a given time points. Stomatal conductance was measured every morning 10:00 to 12:00 hr using SC-1 leaf Porometer for stomatal conductance measurements (Decagon Devices Inc., Pullman, USA)
The design of the experiment was CRD with three replications (leaves as a replication) and data analysis was done using SAS version 9.1. To test differences in weight loss and stomatal conductance one way ANOVA, followed by a Tukey mean separation test was carried out. Correlation analysis was also done to show relationships between different parameters.
For Spathiphyllum sweet checho, statistically, the amount of water transpired significantly differed between treatments (p<0.05) LS 1:20 was significantly different with all treatments except ABA 1.5 mM, ABA 3 mM, SA 5 mM, SNP 200 µM, SNP 400 µM, H2O2 2 mM, H2O2 4 mM and Pyrabactin 400 µM. Whereas, the best active compound treatment, ABA 1.5 mM was only significantly better than CaCl2 100 mM, MJ 100 µM, water, CaCl2 50 mM and matrix. The other best active compound treatments; SA 5 mM, SNP 200 µM, H2O2 4 mM and SNP 400 µM were also significantly different with MJ 100 µM, water, CaCl2 50 mM and matrix. Generally, most best performing active compound treatments were insignificantly different with each other but significantly different with the inferior treatments. Unlike Spathiphyllum Leaf shine was less effective than active compound treatments on Calathea. There were only significant differences for LS 1:20 with SA 10 mM and CaCl2 100 mM and ABA 1.5 mM with SA 10 mM and CaCl2 100 mM. On the other hand, similar to Spathiphyllum the difference in water loss between most active compound treatments was insignificant. Nevertheless, water loss in leaves treated with LS 1:20 remained lower than the other treatments and from active compound treatments ABA 1.5 mM and SA 10 mM were the most and least effective treatments respectively. None of the active compound treatments bested the efficiency of LS 1:20 and some active compound treatments even caused higher amount of water loss than the least effective control treatment in the trial (Table 1)
| No | Treatments | Weight loss (g/g) | |
|---|---|---|---|
| Spathiphyllum sweet checo | Calathearufibarba | ||
| 1 | Water | 1.742( e) | 0.937(ab) |
| 2 | Matrix | 1.759( e) | 0.909(ab) |
| 3 | Leaf shine 1:20 | 0.587(a) | 0.525(a) |
| 4 | Matrix+ABA 1.5mM | 0.981(ab) | 0.545(a) |
| 5 | Matrix+ABA 3mM | 1.092(abcd) | 0.719(ab) |
| 6 | Matrix+Pyrabactin 200mM | 1.224 (bcde) | 0.701(ab) |
| 7 | Matrix+Pyrabactin 400 μM | 1.101(abcd) | 0.839(ab) |
| 8 | Matrix+SNP 200 μM | 1.019(abc) | 0.679(ab) |
| 9 | Matrix+SNP 400 μM | 1.073(abc) | 0.703(ab) |
| 10 | Matrix+H2O2 2mM | 1.153(abcde) | 0.758(ab) |
| 11 | Matrix+H2O2 4mM | 1.028(abc) | 0.891(ab) |
| 12 | Matrix+CaCl2 50mM | 1.743( e) | 0.925(ab) |
| 13 | Matrix+CaCl2 100mM | 1.603(cde) | 1.258(b) |
| 14 | Matrix+SA 5mM | 1.006(abc) | 0.790(ab) |
| 15 | Matrix+SA 10mM | 1.320(bcde) | 1.315(b) |
| 16 | Matrix+H2O2 2mM+SA 10mM | 1.223(bcde) | 0.992(ab) |
| 17 | Matrix+H2O2 4mM+SA 10mM | 1.215( bcde) | 0.995(ab) |
| 18 | Matrix+MJ 100 μM | 1.700(de) | 1.024(ab) |
| 19 | Matrix+MJ 200 μM | 1.384(bcde) | 0.909(ab) |
| Mean | 1.261 | 0.864 | |
| Sig | * | * | |
| CV (%) | 15.7 | 24.8 | |
Table 1: Total weight loss at the 10th day on Spathiphyllum sweet checo and Calathea rufibarba expressed as (g/g) of initial leaf weight. Values are based on the three leaves per treatment. Significant differences are denoted by different lower case letters (a,b).
Statistically significant differences were also observed between treatments on cumulative stomatal conductance readings for both plant species (Table 2)
| No | Treatments | Stomatal conductance (mMol/m2/s) | |
|---|---|---|---|
| Spathiphyllum sweet checo | Calathearufibarba | ||
| 1 | Water | 13.900 (i) | 4.567( f) |
| 2 | Matrix | 20.600( j) | 3.033( cde) |
| 3 | Leaf shine 1:20 | 0.200(a) | 0.000(a) |
| 4 | Matrix+ABA 1.5mM | 1.067(ab) | 0.000( a) |
| 5 | Matrix+ABA 3mM | 2.933(abc) | 0.067(a) |
| 6 | Matrix+Pyrabactin 200mM | 4.433(cdef) | 0.700(a) |
| 7 | Matrix+Pyrabactin 400 μM | 3.867(bcde) | 0.300(a) |
| 8 | Matrix+SNP 200 μM | 1.667(abc) | 0.133(a) |
| 9 | Matrix+SNP 400 μM | 3.467(bcd) | 0.267(a) |
| 10 | Matrix+H2O2 2mM | 3.500(bcd) | 1.233(ab) |
| 11 | Matrix+H2O2 4mM | 6.333(defg) | 0.833(a) |
| 12 | Matrix+CaCl2 50mM | 13.633(i) | 3.833(cdef) |
| 13 | Matrix+CaCl2 100mM | 12.667(hi) | 2.900(cd) |
| 14 | Matrix+SA 5mM | 1.900(abc) | 0.500(a) |
| 15 | Matrix+SA 10mM | 7.533(fg) | 5.300(f) |
| 16 | Matrix+H2O2 2mM +SA 10mM | 6.733(efg) | 4.467(ef) |
| 17 | Matrix+H2O2 4mM+SA 10mM | 9.500(gh) | 2.533(bc) |
| 18 | Matrix+MJ 100 μM | 10.733(hi) | 4.300(def) |
| 19 | Matrix+MJ 200 μM | 4.667(cdef) | 2.500(bc) |
| Mean | 6.868 | 1.971 | |
| Sig | * | * | |
| CV (%) | 15.2 | 24.3 | |
Table 2: A 10 day cumulative stomatal conductance of Spathiphyllum sweet checo and Calathea rufibarba ( mMol/m2/s) Values are based on the three leaves per treatment. Significant differences are denoted by different lower case letters (a,b).
Analogous to weight loss stomatal conductance was higher for Spathiphyllum than Calathea. The correlation analysis showed a significant positive correlation between weight loss and stomatal conductance (R=0.888, P<0.05) for Spathiphyllum and (R=0.811, P<0.05) for Calathea. The relationship between weight loss and stomatal conductance was more potent on Spathiphyllum than Calathea (Figures 1 and 2)
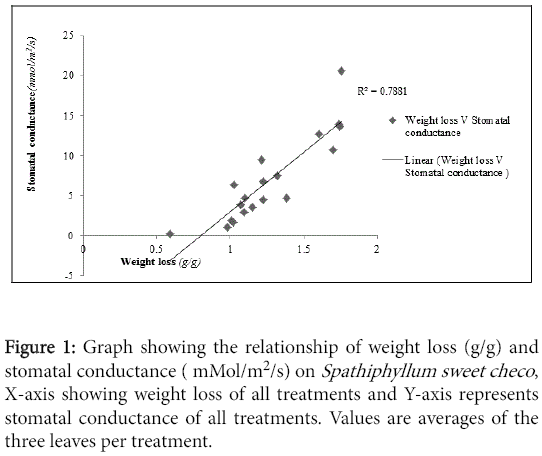
Figure 1: Graph showing the relationship of weight loss (g/g) and stomatal conductance ( mMol/m2/s) on Spathiphyllum sweet checo , X-axis showing weight loss of all treatments and Y-axis represents stomatal conductance of all treatments. Values are averages of the three leaves per treatment.
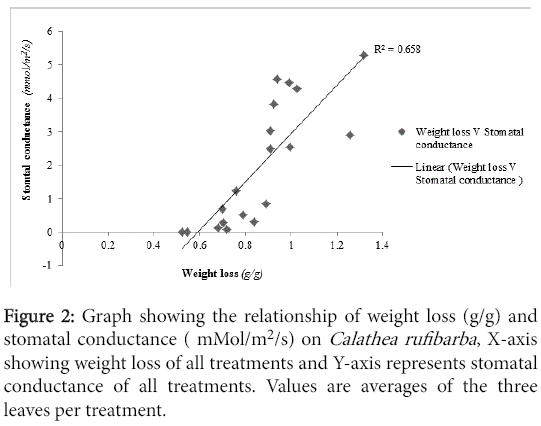
Figure 2: Graph showing the relationship of weight loss (g/g) and stomatal conductance ( mMol/m2/s) on Calathea rufibarba , X-axis showing weight loss of all treatments and Y-axis represents stomatal conductance of all treatments. Values are averages of the three leaves per treatment.
As evidenced by the correlation analysis (Figure 3) the effects of different treatments on either of the pot plants used were relatively similar (R=0.637, p<0.05) However, it might be because of the effect of the leaf surface on the performance Leaf shine and because Calathea leaves has low stomatal conductance, most treatments inflicted lower water loss on Calathea than Spathiphyllum. In addition, on Calathea, it might be because of the delicate nature of the leaves, some active compound treatments also triggered more water loss than the control treatments water and matrix.
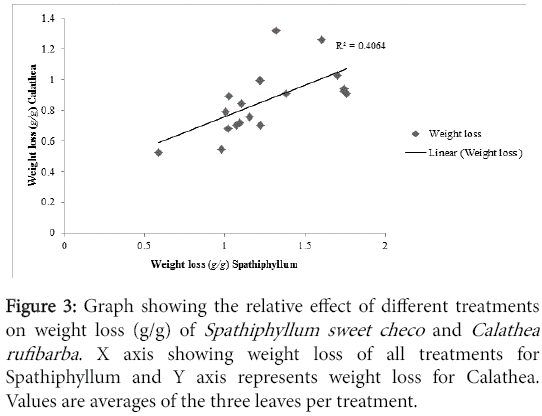
Figure 3: Graph showing the relative effect of different treatments on weight loss (g/g) of Spathiphyllum sweet checo and Calathea rufibarba . X axis showing weight loss of all treatments for Spathiphyllum and Y axis represents weight loss for Calathea. Values are averages of the three leaves per treatment.
The effect of leaf weight and leaf area on transpiration for Spathiphyllum sweet checo is shown (Figure 4) The plotted graph confirms their inseparable effect. The correlation analysis also shows a significant positive correlation between (g/g) and (g/cm2) of weight loss (R=0.985, P<0.05) The same is true for Calathea rufibarba and for the reason that their effect is similar, for future studies either of the measurements can be used.
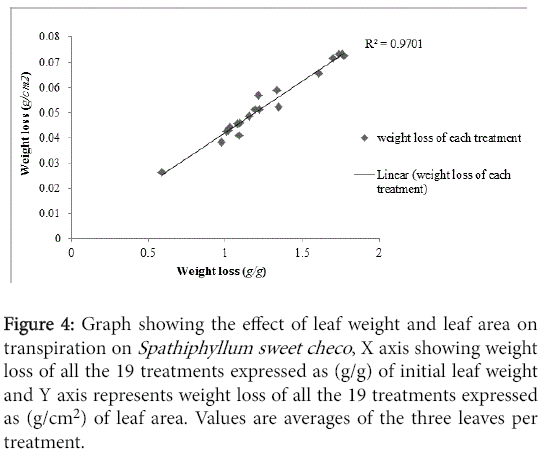
Figure 4: Graph showing the effect of leaf weight and leaf area on transpiration on Spathiphyllum sweet checo , X axis showing weight loss of all the 19 treatments expressed as (g/g) of initial leaf weight and Y axis represents weight loss of all the 19 treatments expressed as (g/cm2) of leaf area. Values are averages of the three leaves per treatment.
Although most of the previous studies done on physiologically active stomata closing compounds were on epidermal strips wherein the opening and closing of stomata can be monitored on a microscope, our experiment only focused on the practical implications by directly spraying the active compound with its carrier on the leaf surface and evaluating their performance on weight loss and stomatal conductance basis. As hypothesized, on both plant species most active compound treatments marked substantial reduction of water loss. However, it wasn’t possible to get an active compound treatment that can surpass the efficacy of leaf shine, only on Calathea rufibarba, an active compound treatment i.e. ABA 1.5 mM, as capable as Leaf shine has been discovered. While, on Spathiphyllum Leaf shine was superior to the best active compound treatment almost by 40%. We suggest the hairs on Calathea leaf surface made it harder for Leaf shine to create a continuous film so as to make a successful physical barrier to obstruct stomatal opening, consequently the best active compound treatment reduced water loss equivalently with leaf shine.
On the other hand, the smooth and glossy surface of Spathiphyllum might’ve been conducive to leaf shine. The manufacturer of Leaf shine also acknowledged the ineffectiveness of their product on hairy leaf surface. Nevertheless, on both plant species most active compound treatments with their effective concentration considerably reduced water loss.
As described as the most successful physiologically active anti-transpirant [11], ABA was the best active compound inducing stomatal closure on both pot plant species. Similar to our results Berkowitz and Rabin [12] and Smiljana et al. [13] found that foliarly ABA treated pepper seedling transplants had higher survival rates mainly attributed by a reduced stomatal conductance and increased leaf water potential. ABA’s agonist Pyrabactin was also effective. As reported by Puli and Raghavendra [14] the stomatal closure either by ABA or Pyrabactin might’ve been reached because of the rise in cytoplasmic PH, reactive oxygen species (ROS) and NO in the guard cells of stomata.
Nitric oxide (NO), as its donor compound SNP and Salicylic acid (SA) were the other effective compounds in either of the plant species. Similar results were also reported using 50μM SNP on excised leaves of wheat (Triticum aestivum), Tradescantia and Poncirus trifoliate [15,16]. Likewise, using SA 1 mM and 10 mM significant reduction of stomatal apertures on epidermal strips of vicia faba [17] and substantial reduction of water loss on excised leaves of co mMelina co mMunis [18] and Phaseolus vulgaris [19] were reported. To achieve their anti-transpirant effect, NO might induced the movement and release of inter-cellular and vacuolar Ca2+ into the cytoplasm of the guard cells [15]. While, SA might triggered the generation of H2O2 and inhibition of H2O2 scavenger catalase [20] and as it was reported recently SA might also induced the generation of extracellular super oxide O-2 and the subsequent increase of cytoplasmic Ca2+ [21,22]. In contrast, H2O2 might also induced stomatal closure by activating Ca2+ channels and by inhibiting the uptake of K+ through K+ channels on plasma membrane of the guard cells [23].
Although there is strong evidence methyl jasmonate can induce stomatal closure, in our experiments, it might have been because of the concentrations we used it couldn’t give us the desired result. Even so, there isn’t a study claiming Ca2+’s anti-transpirant effect, because Ca2+ is the major signaling molecule in the course of stomatal closure we thought it might have some effect and evaluated 50 and 100 mM CaCl2, but it also couldn’t show an anti-transpirant effect. We also thought because SA can inhibit catalase and catalase is H2O2 scavenger, the application of 10 mM SA together with 2 and 4 mM H2O2 might give us a better result, but it was not as good as anticipated. This might’ve been because of the concentration of SA we used, unlike 5 mM SA, 10 mM SA alone was relatively substandard.
Even though, it wasn’t possible to get an active compound superior to Leaf shine and the appearance of leaves sprayed with Leaf shine were better than the other treatments, given the result of our research it’s possible to use active compounds with their appropriate carrier solution as an anti-transpirant agent on pot plants.
I want to forward my sincere appreciation to Netherlands fellowship program (NFP) for funding my study and to my thesis supervisor Professor Dr. Ernst Woltering for his continuous guidance and support throughout the study period. I also want to acknowledge Mr. Sasan Ali for providing me the necessary information about the Porometre and stomatal conductance measurements. I also want to thank Mr. Arjen Van Peppel for his day to day assistance in in the course of the study period.