Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Research Article - (2015) Volume 4, Issue 3
Scorpion stings are life threatening in large parts of the world. Toxins from scorpion venom are responsible for severe metabolic and tissue disruption and immunotherapy is the only specific treatment able to neutralize the toxic effects of scorpion venom. The Androctonus mauretanicus (Am) is the most dangerous scorpion in Morocco, whereas in Tunisia Androctonus australis hector (Aah) is causing most casualties. In this work, we investigated the potential of NbF12-10, a new immunotherapeutic concept based on anti-toxin Nanobodies (Nbs) to neutralize Am scorpion venom. We first explored the immune cross-reactivity between Am and Aah scorpions venoms using anti-AahI and anti-AahII polyclonal, NbAahI’F12 (anti-AahI’), monospecific NbAahII10 (anti-AahII) and bispecific NbF12-10 (anti-AahI’/anti- AahII) monoclonal antibodies and subsequently we study the histological damages observed after envenomation with the F3 toxic fraction of Am scorpion venom by intra-cerebroventricular (i.c.v) injection and the capacity of NbF12-10 to reduce tissue damage induced by F3 fraction after i.c.v administration of F3:NbF12-10 mixture, in mice. Results showed significant para-specific activity of anti-Aah polyclonal and monoclonal antibodies towards Am venom fractions. Histological investigations revealed severe tissue damage in brain, lung and liver after i.c.v. administration of F3 fraction. The NbF12-10 pre-mixed with F3 fraction showed an efficient neutralizing capacity against lethal effect of this toxic fraction. Moreover, in vitro pre-incubation of F3 with NbF12-10 at 8-fold molar excess led to significantly reduced tissue damage. Further, NbF12-10 displays a noteworthy potential to neutralize Am toxins and to rescue 50% of envenomed mice from dying. This study provides first evidence that NbF12-10 nano-therapeutic has promising prospective to treat scorpion envenoming in the Maghreb area.
<Aah: Androctonus australis hector; AahI-AahI’-AahII: Toxins of Aah Venom; Am: Androctonus mauretanicus; BSA: Bovin Serum Albumin; ELISA: Enzyme-Linked Immuno Sorbent Assay; F3: Gel Filtration Toxic Fraction of Am Venom; HCAbs: Heavy-Chain Antibody; i.c.v: Intra-Cerebroventricular; MW: Molecular Weight; Nbs: Nanobody; PBS: Phosphate Buffer Saline; scFv: Single Chain Variable Fragment; SDS-PAGE: Sodium Dodecyl SulfatePolyacrylamide Gel
Envenoming by stings from dangerous scorpions constitute a frequent medical emergency and an important public health problem in many countries [1-8]. In Maghreb countries, this socalled scorpionism, is considered as a major public health problem (Direction du soin et de santé de base, DSSB, Ministère de la santé- Tunisie; Centre antipoison et de pharmacovigilance-Ministère de la santé-Morocco [9]). Various scorpion species are encountered in these regions. For the Androctonus genus, the Androctonus mauretanicus mauretanicus (Amm) renamed Androctonus mauretanicus (Am) is endemic exclusively in the arid region of Morocco (i.e. Marrakech- Tensit-Al Haouz) [6], whereas the natural biotope of Androctonus australis hector (Aah) is in south of Tunisia. The Buthus scorpion genus has the widest spreading throughout the Maghreb. Both Am and Aah sub-species are responsible for the majority of stings, especially in children [4,10]. It is well established that the toxicity of scorpion venom is mainly due to the fast diffusion of toxins throughout the body ([11,12]. These toxins affect ionic channels (i.e. Na+, K+ Ca2+ or Cl- channels) and modulate the transmission of nervous impulses by increasing the discharge of mediators (catecholamines, cytokines, neuropeptide…) causing the release of metabolic serum biomarkers and tissue damages [13-17]. Sodium channel specific toxins are the most represented venom proteins that have been structurally defined and classified in different antigenic groups as identified by polyclonal antibodies. Among these toxins, Amm V is one of the most abundant (46% of the whole Amm venom) and active toxin displaying a 75% sequence identity with AahII [18,19]. However, mass spectrometry analysis of Amm and Aah scorpion venoms revealed other compounds with molecular weight of about 7 kDa. Experimental data has well established that polyclonal antibodies exhibit a cross reactivity to toxins with more than 40% amino acid sequence identity (i.e., AahII/ AmmV/AmmVIII) [20]. However, the idea of the cross neutralizing capacity of polyclonal antibodies has been challenged [21].
To combat clinical symptoms of such hazardous disease, it is absolutely essential to administer the venom-specific antibody as soon as possible after an effective scorpion envenoming. In Tunisia, the use of F (ab)’2 based polyclonal obtained from hyper-immunized horses (with Aah and Bot venoms) is currently recommended as therapy for severe cases, in hospitals. In Morocco, a similar treatment was applied till 2000. Indeed, Fab fragments were also used in some clinical studies [22]. However, the neutralizing potency as well as the specificity of F (ab)’2 or Fab against scorpion toxins were limited due to their polyclonal origin and bio-distribution properties [10,12], [23-25]. Hence, the treatment could fail to rescue patients that are severely envenomed because of the very fast diffusion of scorpion toxins reaching their targets and causing major complications (i.e., pulmonary edema, and respiratory disorder) [26-28] and heart failures [29,30].
In order to improve the immunotherapy against scorpion envenoming, we explored the performance of a novel small antibody fragment that diffuses more rapidly in tissues. This product is derived from the unique Heavy-chain only antibodies (HCAbs) occurring in Camelidae and comprises a single-domain, antigen-binding fragment known as VHH, also referred to as Nbs [31]. Interestingly, the Nbs share a high sequence identity with human VH of family III [32,33]. From VHH library screenings, several Nbs have been retrieved using phage display technology, from which we further selected NbAahI’F12 and NbAahII10 against AahI’ and AahII toxins, respectively [34-36]. The corresponding bispecific NbF12-10 format of only 29 kDa yielded a therapeutic compound with a strong protective capacity against the whole Aah venom in mice [35].
However, the potential of NbF12-10 to neutralize venoms of other scorpions such as that from Androctonus mauretanicus scorpion, living in Morocco remains unexplored. Turning this challenge into practice, we first investigated the para-specificity by testing the capacity of polyclonal (anti-AahI and anti-AahII), monoclonal (NbAahI’F12 and NbAahII10) and bispecific antibodies (NbF12-10) to recognize and to bind Am venom compounds of black Morocco scorpion, using an ELISA assay. Thereafter, we assessed the potency of NbF12-10 to reduce histological lesions, observed in vital organs, after experimental pre-incubation with the most toxic fraction from Am venom. Our data demonstrate the capacity of NbF12-10 to reduce fatal disturbances provoked by a lethal dose of toxic fraction from Moroccan scorpion venom.
Animals
Male Swiss mice (20 ± 2 g) were housed under controlled temperature in the animal breeding unit at Pasteur Institute of Tunis. Animals used for toxicity assays and histological experiments were approved by the institution Research Board of the Pasteur Institute of Tunis and carried out according to the European Community Council Directive (86/609/EEC) for experimental animal care and procedures.
Scorpion venom
The venom of Androctonus mauretanicus individual scorpions was collected at the Pasteur institute of Morocco using electrical stimulation. The lyophilized venom was diluted in cold water (2 v/v) and centrifuged for 15 min at 4°C to remove lipid compounds and debris. The venom supernatant was stored at -20°C. The protein concentration was determined according to the Bradford method [37] using and BSA (Bovine serum albumin) as standard and the Bio-Rad reagent.
Production of polyclonal antibodies
Immune sera against AahII and AahI, the most active toxins from Aah scorpion venom, were expressed by immunizing New Zealand rabbits as described previously [38]. After collection of pre-immune sera, each animal was first injected intradermally at 10 μg of toxin (AahI or AahII) in Freund’s Adjuvant complete (Sigma) and then, rabbit received two injections of 10 μg and two injections of 15 μg of toxins in Freund’s Adjuvant incomplete (Sigma) through the subcutaneous route every 3 weeks.
Production of nanobodies
The production and purification of Nanobodies (i.e., NbAahI’F12, NbAahII10 and NbF12-10) was according to previously reported protocols [34,35]. Briefly, the clone harboring the gene for the AahI’, the AahII or the bispecific NbF12-10 protein was cultured separately in shaker flasks with Terrific Broth supplemented with 100 μg ampicillin/ ml and 0.1% glucose, until the absorbance at 600 nm reached a value between 0.6 and 0.9. At this time, the recombinant protein expression was induced with 1 mM isopropyl-thiogalactopyranoside and incubation for at least 16 h at 28°C. From the periplasmic compartment, proteins were extracted by osmotic shock [39] and loaded on a Ni-NTA super-flow Sepharose column (Qiagen). The purity of His-6 tagged NbF12-10 was checked by sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and the final yield was monitored from the UV absorption at 280 nm and the theoretical extinction coefficient of the Nb, calculated from its amino acid content, as previously described [35].
Gel-filtration chromatography of venom
The supernatant of the Am venom was loaded on a Sephadex G50 column (2.6 × 100 cm) (Sigma) for size exclusion chromatography using 1.6 M acetic acid as equilibration and running solution (28 ml/h) according to Miranda et al. [40] with minor modifications [41]. Fractions (2.5 ml/tube) were collected in a fraction collector (Frac-920, GE). Absorbance was assessed at 280 nm using a spectrophotometer (Beckman DU 640-UV/Visible). Pooled fractions were stored at -20°C till used for toxicity and histological assays.
In vivo assay
The toxicity of scorpion venom Am and derived fractions was measured in mice by i.c.v injection. The LD50 represents as amount of venom or toxic fraction leading to 50 % of mice surviving the injection [34,36]. Survivors were counted after 24h.
Polyacrylamide gel electrophoresis
Determination of molecular weight (MW) of Am venom and its toxic fraction F3 were performed in sodium dodecyl sulfate-containing polyacrylamide gels [42]. SDS-PAGE was performed under reducing conditions. Proteins were stained with Coomassie brilliant blue.
Immunological tests: indirect ELISA
ELISA were carried out using anti-AahI, anti-AahII specific polyclonal antibodies, NbAahI’F12 and NbAahII10 monospecific and NbF12-10 bispecific nanobodies, following standard protocols to explore their antigenic cross-reactivity against Am scorpion whole venom as well as F2 and F3 toxic fractions of Am venom. Micro titer 96 wells plates (Nunc Maxisorp Plate) were coated overnight at 4°C with 100 μl of 1 μg/ml of Am venom, F2 or F3 fractions in PBS buffer (Phosphate-Buffered-Saline). After washing with 0.1% Tween-PBS, the residual protein binding sites were blocked with 3 % milk in PBS, for 1 hr at 37°C. Subsequently, after washing steps as before, 100 μl of primary antibody (i.e. mouse anti-AahI or anti-AahII polyclonal antibodies, kind gift from Dr Christiane Devaux) or NbAahI’F12, NbAahII10 and NbF12-10 monoclonal antibodies) (at appropriate dilution or concentration as indicated below) were added. After a washing step, secondary peroxidase conjugated antibody (i.e. rabbit anti-mouse IgG (Sigma) or mouse anti-his-6 clone BMG-His-1, Roche laboratory, as appropriate) were added, After washing, ortho-phenylenediamine dihydrochloride-H202 (Sigma) was added as substrate and the reaction was stopped using 2N sulfuric acid. Absorbance was measured with a spectrophotometer (Multiskan EX, Thermo Electron Corporation) at 492 nm. All samples were applied in duplicate and ELISA procedures were repeated three times. All polyclonal antibodies concentrations have been adjusted to give similar optical density readout in the ELISA.
Pathophysiological study of F3 fraction from Am scorpion venom in mice
To highlight the tissue damage induced by F3 fraction, three groups of 4 mice were used. The first group as control received physiological saline. Second and third groups were challenged with 30 and 35 ng of F3 toxic fraction, an amount corresponding to 2 and 2.3 LD50 respectively. All injections were carried out intra-cerebroventricularly. The levies of organs: brain, lungs and liver were performed in postmortem animals.
In vivo neutralizing of the F3 toxic fraction with NbF12-10
To investigate the in vivo neutralizing capacity of NbF12-10 against toxic effect of F3 fraction, five groups of mice were injected with two lethal doses of F3 fraction: 30 and 35 ng (corresponding to 2 and 2.3 LD50) pre-incubated for 1 hour at 37°C with NbF12-10 at molar ratios in cascade from 1:1 to 1:8 molar excess of NbF12-10 dose (Table 1).
| Group of four mice | LD50 (F3 fraction) | Molar ratio F3:NbF12-10 |
|---|---|---|
| 1 | 2 | 1:1 |
| 2 | 2 | 1:2 |
| 3 | 2 | 1:4 |
| 4 | 2.3 | 1:4 |
| 5 | 2.3 | 1:8 |
Table 1: Different molar ratios of F3:NbF12-10 pre-incubated mixtures.
Organs: brain, lungs and liver were conducted in postmortem animals or in survivors after 24 hours of experimentation.
Experimental conditions for histological analysis
To analyze the histopathological effect on tissues induced by the F3 fraction of Am scorpion venom and the effectiveness of the NbF12-10 to neutralize the F3 that we checked for possible reduced tissue damage after i.c.v. injection of F3 toxins pre-incubated with the bispecific Nb construct. Brain, lung and liver were extracted from control animals, envenomed mice or treated with F3:NbF12-10 mixture. The organs were fixed in 10 % formalin solution, embedded in paraffin, sliced (2 to 4 μm) and stained with hematoxylin-eosin for microscopic analysis using a Carl Zeiss Axiskop 50 microscope (Germany).
Statistical analysis
Significance of the differences in the immune-reactivity towards Am venom and its fractions F2 and F3 of the polyclonal (anti-AahI, anti-AahII) and monoclonal antibodies (NbF12-10, NbAahI’F12 and NbAahII10) was assessed by Student’s t-test (the level of significance was p < 0.05).
Venom fractions
The fractionation of soluble Am venom by gel filtration on Sephadex G50 shows six distinct fractions: F1-F6 (Figure 1). Bradford protein quantification showed that F3 (15 μg/μl of protein) fraction represents 41.15% of the venom proteins (Figure 2).
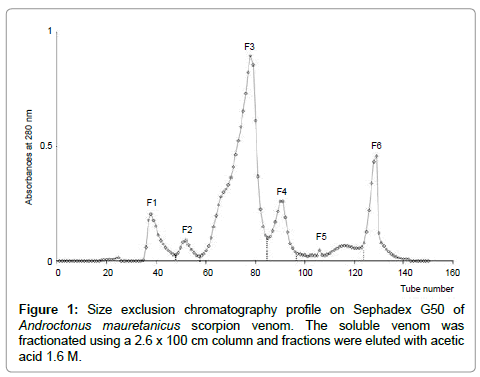
Figure 1: Size exclusion chromatography profile on Sephadex G50 of Androctonus mauretanicus scorpion venom. The soluble venom was fractionated using a 2.6 x 100 cm column and fractions were eluted with acetic acid 1.6 M.
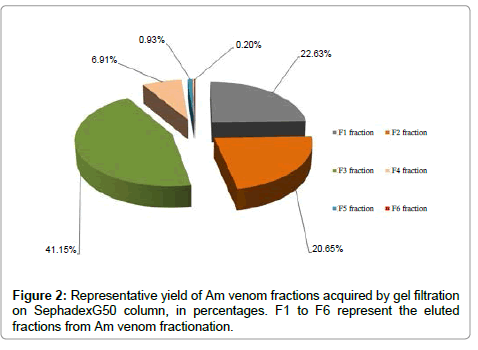
Figure 2: Representative yield of Am venom fractions acquired by gel filtration on SephadexG50 column, in percentages. F1 to F6 represent the eluted fractions from Am venom fractionation.
In vivo toxicity assay
The toxicity of F1, F2 or F3 fractions was assessed in mice by i.c.v injection of different amounts (between 5 ng to 2 μg) into Swiss mice (weighting 18-20 g). The number of surviving mice was monitored at 24 hr after toxin injection. As negative control, mice received only physiological solution (NaCl 0.9%). Results showed that F1 is not toxic below 2 μg injected per mouse while F2 causes envenoming symptoms, no mortality was recorded. In contrast, F3 is causing highly toxic symptoms and mortality. The LD50 of F3 fraction was estimated (using cohorts of 4 mice) to approximately 15 ng/mouse. F4 to F6 fractions showed small low molecular weight peptides displaying low toxicity. As expected, F3 fraction contains peptides with apparent molecular weight (MW) related to toxin’s MW (Figure 3).
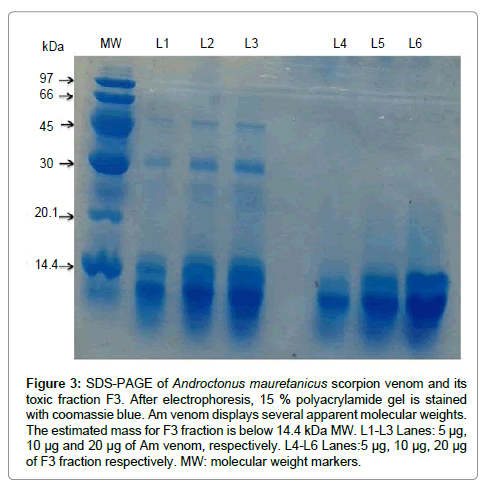
Figure 3: SDS-PAGE of Androctonus mauretanicus scorpion venom and its toxic fraction F3. After electrophoresis, 15 % polyacrylamide gel is stained with coomassie blue. Am venom displays several apparent molecular weights. The estimated mass for F3 fraction is below 14.4 kDa MW. L1-L3 Lanes: 5 μg, 10 μg and 20 μg of Am venom, respectively. L4-L6 Lanes:5 μg, 10 μg, 20 μg of F3 fraction respectively. MW: molecular weight markers.
Immune cross-reactivity between Am and Aah using AahI and AahII-specific polyclonal: Indirect ELISA
The cross-reactivity of polyclonal antibodies directed against the major toxins from Aah venom: anti-AahI, anti-AahII, towards Am venom or fractions F2 and F3, showed successful recognition of the F3 Am fraction by both anti-AahI and anti-AahII polyclonal antibodies with a very high significant p value (p = 0.0008 and p = 0.001, respectively). Also, anti-AahI and anti-AahII reacted immunologically with Am venom (p = 0.02; p = 0.004 respectively) (Figure 4A,4B) whereas F2 fraction revealed low reactivity to anti-AahII (p = 0.035) (Figure 4B).
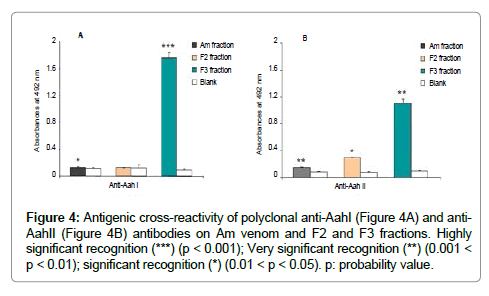
Figure 4: Antigenic cross-reactivity of polyclonal anti-AahI (Figure 4A) and anti- AahII (Figure 4B) antibodies on Am venom and F2 and F3 fractions. Highly significant recognition (***) (p < 0.001); Very significant recognition (**) (0.001 < p < 0.01); significant recognition (*) (0.01 < p < 0.05). p: probability value.
Immune cross-reactivity between Am and Aah scorpion venoms using NbF12-10, NbAahI’F12 and NbAahII10 specific monoclonal antibodies: Indirect ELISA
We evaluated immunological binding capacity of monoclonal Nbs: NbF12-10, NbAahI'F12 and NbAahII10 towards Am venom and its fractions F2 and F3. An antigenic cross-reactivity against F3 fraction is observed when NbF12-10 (Figure 5A) and NbAahI’F12 (Figure 5B) are used at concentration of 5μg/ml, and the p-values are highly significant (p = 0.0028 and 0.0022, respectively). A significant reactivity of NbAahI’F12 against Am venom is noted (p = 0.0025) (Figure 5B). Remarkably, only poor binding of NbAahII10 to Am venom as well as to F3 fraction is noticed (Figure 5C). No affinity has been noted between Nbs (NbF12-10, NbAah’F12 and NbAahII10) and F2 fraction
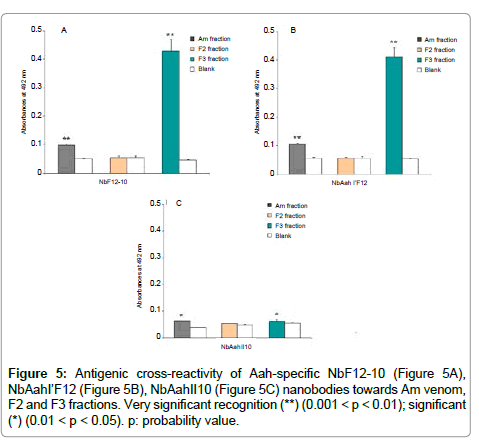
Figure 5: Antigenic cross-reactivity of Aah-specific NbF12-10 (Figure 5A), NbAahI’F12 (Figure 5B), NbAahII10 (Figure 5C) nanobodies towards Am venom, F2 and F3 fractions. Very significant recognition (**) (0.001 < p < 0.01); significant (*) (0.01 < p < 0.05). p: probability value.
Histology of brain, lung and liver tissues upon F3 injection with or without NbF12-10
Pathophysiological effects of F3 fraction have been investigated in mouse by i.c.v injection. Histological screening of organs sections from mice envenomed by F3 toxic fraction with two doses of 30 ng and 35 ng (corresponding to 2 LD50 and 2.3 LD50, respectively), showed 100% mortality of mice poisoned and structural changes in brain, lung and liver tissues. Indeed, in brain tissue, a marked congestion, vasodilatation, hemorrhage, edema and neuronal constriction were observed (Figure 6: panels B-B1 and K). In lung parenchyma, we observed many edemas in intra-alveolar and bronchiolar levels, and half of the lung tissue was invaded by edema and congestion. Moreover, several hemorrhagic foci, large fibrin deposition, infiltration of inflammatory cells and loss of alveolar walls were observed (Figure 6: panels E and M-M1). In liver tissue, F3 fraction caused congestion, sinusoidal dilatation, hepatocyte turgescence, hepatocyte with nuclear hypertrophy or bi-nucleating (Figure 6: panels H and O). Histological slices of control animals showed normal architecture structural (Figure 6: panels A, D-D1 and G).
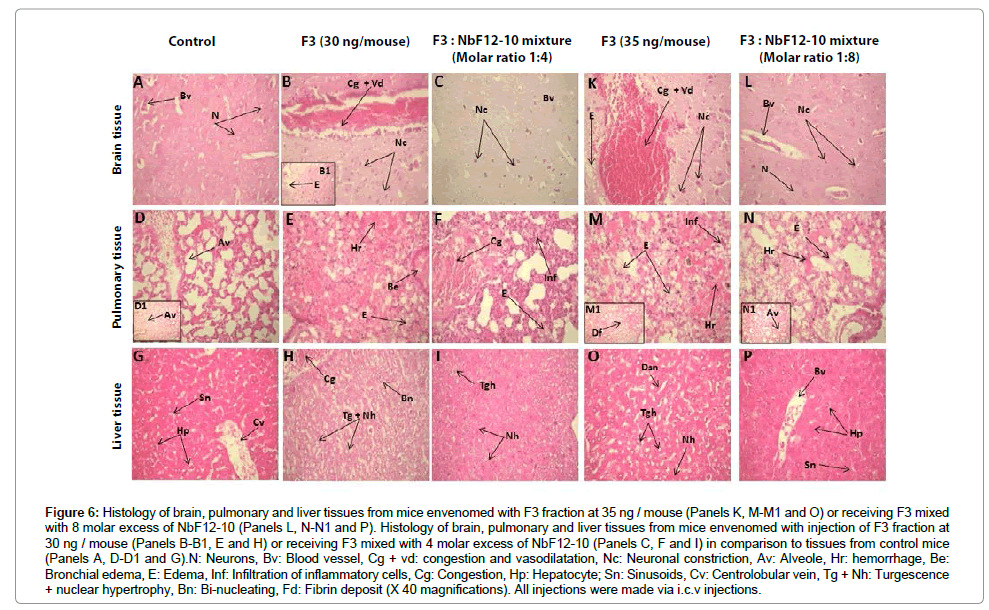
Figure 6: Histology of brain, pulmonary and liver tissues from mice envenomed with F3 fraction at 35 ng / mouse (Panels K, M-M1 and O) or receiving F3 mixed with 8 molar excess of NbF12-10 (Panels L, N-N1 and P). Histology of brain, pulmonary and liver tissues from mice envenomed with injection of F3 fraction at 30 ng / mouse (Panels B-B1, E and H) or receiving F3 mixed with 4 molar excess of NbF12-10 (Panels C, F and I) in comparison to tissues from control mice (Panels A, D-D1 and G).N: Neurons, Bv: Blood vessel, Cg + vd: congestion and vasodilatation, Nc: Neuronal constriction, Av: Alveole, Hr: hemorrhage, Be: Bronchial edema, E: Edema, Inf: Infiltration of inflammatory cells, Cg: Congestion, Hp: Hepatocyte; Sn: Sinusoids, Cv: Centrolobular vein, Tg + Nh: Turgescence + nuclear hypertrophy, Bn: Bi-nucleating, Fd: Fibrin deposit (X 40 magnifications). All injections were made via i.c.v injections.
To evaluate the capacity of NbF12-10 to neutralize the toxicity caused by 30 or 35 ng (i.e. 2 LD50 and 2.3 LD50) of the F3 toxins from Am scorpion venom per mice, we pre-incubated the toxins with NbF12-10 at various molar ratios before i.c.v. injection (Table 1). The NbF12-10 was added at an estimated 1 to 8 molar excess over the toxins in the F3 fraction, as indicated. The mice from the first, second and third group received 30 ng of F3 pre-incubated with either 120 ng, 240 ng or 480 ng of NbF12-10 (that correspond to a molar ratio of toxin: Nb of 1: 1, 1:2 and 1:4, respectively). The fourth and fifth groups of mice received 35 ng of F3 pre-incubated with 560 ng or 1120 ng of NbF12-10 (corresponding to 4 or 8-fold molar excess of Nb, respectively).
Our results revealed that NbF12-10 ensures the survival of 50 % of mice injected with 35 ng of the F3 fraction with an 8 fold molar excess of Nb (Table 2). Hence, a significant Am-neutralizing capacity of the NbF12-10 was noticed under these conditions whereas a reduced excess of NbF12-10 (i.e. equimolar to 1:4 ratio) in the mixtures failed to neutralize a 2 LD50 of F3-Am fraction as all mice died from F3 poisoning and important structural changes in tissues were observed experimentally.
| Table 2 Group of four mice | Molar ratio F3:NbF12-10 | Survivors mice |
|---|---|---|
| 1 | 1 : 1 | 0/4 |
| 2 | 1 : 2 | 0/4 |
| 3 | 1 : 4 | 0/4 |
| 4 | 1 : 4 | 0/4 |
| 5 | 1 : 8 | 2/4 |
Table 2: Efficiency of NbF12-10 against the toxic effects of fraction F3.
Interestingly, the survivals didn’t exhibit any severe symptoms of envenoming (e.g. neurological signs, agitation and lumps dorsal are missing) and damages were clearly less significant in the target tissue of envenomed mice that survived following injection. Indeed, the detailed histological study of tissue structure demonstrates a significant reduced proportion of tissue damage within the organ sections. In particular, brain parenchyma was completely devoid of edema and nerve cells display their common feature (Figure 6: panel L). Regression of lung edema was evident. The number of hemorrhagic areas was less important nearby minor congestions and no deposited fibrin was noticed (Figure 6: panel N-N1). Moreover, liver tissue revealed a similar architecture to control tissue without dilatation of sinusoids, neither cell turgescence (Figure 6: panel P). However, a lower molar excess of NbF12-10 (30 ng of F3 fraction mixed to 4-fold molar excess of NbF12-10) resulted in a severely reduced experimental neutralizing efficacy and mice from this cohort failed to survive. Hence, histological analysis of post-mortem animals showed tissue damage as well. In cerebral cortex, we noted extravasation of blood (data not shown), edema, less regression of neurons and congestion of a large vessel (Figure 6: panel C). Pulmonary tissue was invaded by edema (Figure 6: panel F). Sinusoidal dilatation and a minority of hepatocyte turgescence were observed (Figure 6: panel I).
In the present study, we have demonstrated that F3, a toxic fraction from Am scorpion venom has led to structural abnormalities. This fraction corresponds to most abundant toxic component, obtained after analytical filtration of Am venom (Figure 1). F3 fraction showed a similar retention time as that of AahG50 (derived from Aah venom) fraction whereas the eluted fractions on either side of this F3 peak display significant divergence in retention time [34,36]. The LD50 of F3 fraction, estimated as the amount of toxins whereby 50% of the mice (cohorts of 4 mice) survived the injection that corresponds to 15 ng/mouse, approximately. However, previous experiments were conducted using intra-peritoneal injection of the toxins or venoms. Here, we investigated for the first time the envenoming effect after injection of an Am toxic fraction (F3) via an i.c.v. route with 30 and 35 ng (equivalent to 2 LD50 and 2.3 LD50). Histological examination showed that both doses provoked considerable tissue damage leading to significant structural changes in brain, lung and liver parenchyma.
It has been demonstrated that toxins from scorpion venom target to their specific receptors in sodium, potassium channels of nerve and muscle cells which disrupt the propagation of nerve impulses (activation or inactivation of channels) and cause the release of neurotransmitters and cellular mediators. Such changes is causing disturbance of metabolic parameters (glycemia, urea, creatinine, …) and structural disorganization leading to dysfunction of vital organs have been described (i.e. brain, lungs, heart, liver, kidneys) [15,16], [43,44].
Indeed, cerebral alterations were assumed to be the consequence of neurotransmitter releases [43-51]. More recently, similar structural damage in brain tissue (i.e., edema, hemorrhage, neuronal darkness) were evoked by a small scorpion toxin called Kaliotoxin isolated from Aah [52].
Many studies showed similar results in pulmonary tissues [14-16], [53,54]. It was highlighted that scorpion envenomation is followed by an imbalance of pro-and anti-inflammatory response [55] and scorpion venom is able to affect parameters for pulmonary mechanisms [56]. The fibrin deposit was associated with disruption of blood coagulation process generated after scorpion envenomation [57]. The genesis of pulmonary edema can be attributed to a cardiogenic dysfunction [58]. Moreover, it may be non-cardiogenic origin [17,59] and consequently, pulmonary edema is in that case multifactorial.
In liver tissue, similar observations (i.e., hepatocytes with cytoplasm distended to nuclear hypertrophy and large vacuoles) were noticed after envenomation by Tityus genus [14]. Otherwise, hepatocyte cells may exhibit Mallory bodies resulting from hepatic steatosis [60].
Hence, to remove the circulating scorpion venom toxins, immunotherapy is the specific treatment to be applied. Currently, F (ab’)2 based therapy is the only anti-venom therapy available into market for clinical envenoming treatment. This so-called fabotherapy is based on administration of scorpion anti-venom serum produced in preimmunized horses. As polyclonal product, F (ab’)2-based antivenom has potency to prevent histopathological injuries in tissue and to return into normal, metabolic parameters and electrocardiogram changes caused in patients after scorpion envenoming [61,62]. However, the efficiency of intra-muscular route for antivenom treatment of envenomed patients was not reported systematically in all cases [63]. Hence, avian anti-Tityus cariptensis (Tc) venom antibodies (isolated from chicken egg yolks) was recently applied as anti-venom alternative and was effective in neutralizing only 2 LD50 doses of Tc venom [64]. A mixture of antibody fragments F (ab’)2/Fab is able to significantly reduce local leukocytosis, hemorrhage and inflammatory edema induced Aah venom and its toxins [25].
In this study, we first studied the para-specificity between Aah and Am venoms using polyclonal (anti-AahI and anti-AahII) and monoclonal (NbF12-10, NbAahI’F12 and NbAahII10) antivenom against Am venom and its fractions F2 and F3. Thereby, the immunological screening using polyclonal antibodies and Nbs favors the presence of several AahI, AahI’ and AahII related or crossreacting toxins within Am venom and F3 fraction. Indeed, Am venom molecules (F3 fraction) could be detected easily by both, monospecific NbAahI’F12 and the bispecific NbF12-10 construct, which is suggestive for the presence of AahI related toxins or epitopes within the F3 fraction of Am venom. Similar results reported that human single chain fragment (scFv) recognized toxins of Centruroides noxius Hoffmann and Centruroides suffuses scorpion venom and rescue mice from severe envenomation by whole venom of Centruroides suffuses and Centruroides noxius [65]. Also, polyclonal anti-venom against Hadruroides lunatus displayed cross reactivity with venom from Tityus serrulatus, Centruroides sculpturatus and Androctonus australis hector scorpions [66]. Other study has explained that anti-AmmVIII (Anatoxin from Am venom) antibodies were also found to cross-react towards several of the peptides designed from the AahII structure [67]. Devaux et al. (2001) [68] reported that 9C2 (scFv), a murine monoclonal IgG, was able to neutralize AahI toxin from Aah scorpion venom.
Herein we demonstrated in the first time, the effectiveness of Aahspecific NbF12-10 nanotherapeutic to bind and neutralize Am venom related toxins and to prevent severe structural abnormalities at cerebral, pulmonary and hepatic parenchyma, provoked by i.c.v injection of Am venom F3 fraction. More interestingly, pre-incubation with NbF12- 10 leads to less toxic mixture and therefore envenomed mice rescued from death. Similarly, F7Nb nanobody raised against Heminecrolysin (major known hemolytic and dermonecrotic fraction of Hemiscorpus lepturus scorpion) venom showed in vitro and in vivo neutralization of heminocrolysin activity [69]. Clearly, our data revealed that i.c.v route is very sensitive and associated with distant and significant structural lesions and confirms the importance of metabolic releases in severe and dramatic symptoms [70,71]. Further investigations are in progress to test additional nanobody candidates with sub-nanomolar affinity for the AahI’ and AahII toxins, separately or in combination and to study the effect of NbF12-10 against whole Am venom. Indeed, study on large cohorts of animals and using classical pharmacokinetics/ pharmacodynamics experiments is in progress.
In this study, we demonstrated a significant antigenic crossreactivity (p < 0.05) of polyclonal antibodies and Nbs between the Tunisia Aah and Morocco Am scorpion venom components. Especially, the result with NbF12-10 is important, as this is our lead product (produced against the major toxins from Aah scorpion venom of Tunisia) with which we provided recently the proof-ofconcept to neutralize the Aah envenoming. These promising data prompted us to further study it’s in vivo neutralizing capacity against experimental Am envenoming. Our results with this bispecific Nb on the Am venom highlights its effectiveness to reduce pathophysiological effects of toxicity caused by the major toxic fraction of this Moroccan Am scorpion. Evidence is provided that this Nb construct might be seriously considered as promising anti-venom therapeutic to treat severe scorpion envenoming cases in the large Maghreb area. Further preclinical studies to include larger cohorts of mice and other experimental animals should be considered in the future.
This work was partially supported by International Pasteur Institute Network (RIIP), Islamic Bank for Development (BID), Pasteur Institute of Morocco (IPM) and Pasteur Institute of Tunis (IPT) grants. We would like to thank Prof. Serge Muyldermans for support in identifying Nanobodies. Our gratitude’s are addressed to Professors Naima ELMdaghri Director of Pasteur Institute of Morocco, Hechmi Louzir General Director of Pasteur Institute of Tunis for encouraging this research project. Our thanks are extended to Dr. Zakaria Benlasfar (IPT) and Lotfi Boussada (IPM) for animal facilities; we also thanked Dr. Abderrahmane Belhouari, Faculty of Sciences, Ben M'sik, Casablanca, Morocco for assistance in statistical data analysis.
Fatima Chgoury conducted biochemical and experimental studies. Rahma Ben Abderrazek and Naoual Oukkache contributed to technical and experimental work. Issam Hmila contributed in part to Nanobody production and purification; Haifa Tounsi and Samir Boubaker for design and analysis of histological sections; Rachid Saïle, Noreddine Ghalim and Mohamed El Ayeb for analysis and discussing results. Fatima Chgoury and Balkiss Bouhaouala-Zahar are responsible for writing this manuscript. Balkiss Bouhaouala-Zahar was responsible for designing experiments, result interpreting and the editorial corrections.
The authors declare that there are no conflicts of interest.