Anatomy & Physiology: Current Research
Open Access
ISSN: 2161-0940
ISSN: 2161-0940
Research Article - (2017) Volume 7, Issue 6
Neural crest cells give rise to melanoblasts that invade the dermis and epidermis to differentiate into melanocytes that can develop melanoma, which is one of the most aggressive forms of human cancer. Identification of mechanisms important for the initial development of melanoma is hampered because of the difficulty in studying them in transgenic mice with melanoma. In addition, there is only fragmentary knowledge about the histochemical and biochemical changes that accompany the very early stages of neural crest/premelanoblast invasion into the skin, which may later develop melanoma, in the naturally occurring melanoma models such as Xiphophorus fish hybrids. Herein, we used Xiphophorus hybrid fish to investigate the distribution and extracellular matrix (ECM) environment of premelanoblasts during early stages of skin development, and to test the hypothesis that there are parallels between neural crest/premelanoblast invasion and ECM changes in the early developing skin. To test this hypothesis, we investigated neural crest/premelanoblast cell distribution through development and characterize their ECM environment. We found several histochemical evidences for the correlation between this pattern of developing skin invasion by neural crest cells/premelanoblasts and the spatiotemporal relationship of extracellular matrix components that stimulate cell migration/invasion such as collagen, and others such as non-sulfated glycosaminoglycan that may facilitate cell invasion by enhancing the intercellular spaces and inhibition of extensive intercellular interactions by physically separating cells. These findings help to uncover some histochemical and biochemical changes that accompany the early stages of skin formation in Xiphophorus fish hybrids, and can also provide a conceptual framework for future mechanistic studies in this area, and for studies that use this fish hybrid as a naturally occurring melanoma model.
Keywords: Skin; Neural crest cells; Premelanoblasts; Extracellular matrix
The skin is the largest organ in the body, and protects against environmental insults and dehydration. In mammals, skin contains three major cell types, which have diverse origins; epithelial cells forming a stratified epidermis that contains keratins; mesenchymal cells forming the underlying dermis and, together with epithelial cells, form hair follicles and other appendages; and melanocytes that are responsible for pigmentation. These cell types undergo intensive interactions, migration, proliferation, and differentiation, which are controlled by a small number of signaling pathways such as WNT, Hedgehog, BMP, FGF and TGF, during embryogenesis and morphogenesis [1]. During embryogenesis, the epidermis originates from the surface ectoderm that proliferates and migrates from the dorsal midline to cover the whole embryo, while the lateral plate mesoderm forms to ventral trunk dermis, and head dermis arises, at least partially, from neural crest cells [2,3].
In fish, the embryonic epidermis consists of two characteristic cell layers: the surface layer or enveloping layer (EVL), and the inner layer that is also called the epidermal basal layer (EBL). These two layers are formed at the blastula and gastrula periods, respectively, and they become the simple epithelium of the embryonic fish skin [4]. Neural crest cells (NCCs) give rise to premelanoblasts that differentiate into melanocytes, which manufacture melanin, providing pigmentation of the epidermis and hair follicles. After an initial period of proliferation, premelanoblasts start migration between the developing dermatome and overlying ectoderm, and then they migrate through the dermis. Then, melanoblasts move from the dermis to invade the epidermis during later stages of embryonic development [5,6]. The processes of neural crest migration and differentiation as well as melanoblast migration, proliferation, and/or survival are tightly controlled by several genes/factors, in which mutations can cause pigmentation and other neural crest-derived defects or diseases [7,8]. One of major skin diseases is melanoma, which is a tumor of transformed melanocytes that are originally derived from the embryonic neural crest. Melanoma is one of the most aggressive forms of human cancer, with the fastest increase in incidence of all tumors. The fish genus Xiphophorus has long been established as a model organism for study of melanoma formation. Wildtype Xiphophorine fish are insusceptible of developing neoplasia. However, certain backcrosses of swordtail (Xiphophorus helleri ) with platyfish (Xiphophorus maculatus) as the recurrent parent produce offspring that can develop spontaneous neoplasia/melanoma in a Mendelian fashion [9,10]. Currently, there is only fragmentary knowledge about the histochemical and biochemical changes that accompany the very early stages of neural crest cells/premelanoblasts invasion into the developing skin of Xiphophorus hybrid fish. Herein, we investigated the invasion, distribution and extracellular matrix environment of neural crest/premelanoblasts during early skin development in Xiphophorus hybrid fish. We found that there are parallels between neural crest/premelanoblast invasion and ECM changes in the early developing skin.
Xiphophorin fish embryos
We obtained Xiphophorin fishes; platyfishes (Xiphophorus maculatus ), and swordtails (Xiphophorus helleri ) from a tropical fish shop. Fishes were bred in the laboratory to produce Platyfish-Swordtail Hybrids, as previously described [11,12]. Briefly, interspecific hybrids are produced by crossing female platyfish with male swordtail fishes. The backcross hybrids are produced by backcrossing hybrid females of the first generation to the parent male swordtail fish. Staging of fish embryos was carried out using criteria described by Tavolga and Sadaghiani and Vielkind [13,14].
Fixation of xiphophorin embryos for light and scanning electron microscopy (SEM)
Embryos for SEM observation were prepared as described [15-17]. Briefly for light microscopy, Xiphophorus embryos were fixed in 4% paraformaldehyde in PBS (pH 7.2) for 2-6 hours at room temperature or overnight at 4°C and washed in three changes of PBS during which the yolk was removed. For SEM, embryos were fixed in 2% glutaraldehyde in saline G for 1-20 hours and post-fixed by treatment with 1% OsO4 in 0-1 M cacodylate buffer at pH 7.4 and thiocarbohydrazide as described by Kelley [18]. The fixed specimens were then dried by the critical point method, gold-coated and examined using SEM microscope, as described before [14]. Briefly, dried specimens were mounted on aluminium stubs by using doublestick tape. Then, we removed the ectoderm of embryos using a small piece of tape. We also aimed to separate the epidermis from the dermis to further analyze the underside of the epidermis. The removed/ separated tissue was mounted on the same stub for further and detailed examination of its inner surface [14]. The mounted specimens were coated with gold, and viewed at 25 kV in a Jeol JSM-25511 scanning electron microscope.
Identification of extracellular matrix (ECM) components
Specimens of Xiphophorin embryos of various ages were fixed in Carnoy’s solution or 10% formalin in phosphate-buffered saline (PBS) with 0.5% cetylpyridinium chloride (CPC; Polysciences) and 0.25% polyvinylpyrrolidone (PVP; CPC and PVP maximally preserve glycosaminoglycans/proteoglycans) overnight at room temperature. Embryos were subsequently dehydrated in ethanol, cleared in xylene, and embedded in Paraplast. Specimens were then sectioned and stained with 1% Alcian blue (pH 1.0; stains only sulfated glycosaminoglycan (GAGs)/mucopolysaccharides; Green and Pastewka [19], 1% Alcian blue (pH 2.6; stains both sulfated and nonsulfated GAGs; Green and Pastewka, 1974) [19], or Alcian blue (pH 2.6) periodic acid/Schiff (PAS; stains neutral polysaccharides) double staining method overnight, as described [20-22]. Some sections were stained for collagen using van Gieson’s stain, as described [17,22,23].
Generation of xiphophorus hybrid fish
In the Gordon-Kosswig genetic cross, the platyfish (X. maculatus ) and the swordtail fish (X. helleri ) are routinely used to create F1 hybrids; when these fish are mated to X. helleri to generate a firstgeneration backcross (BC1), progeny that may spontaneously develop invasive and/or exophytic melanomas [11,12]. We used this genetic cross to generate Xiphophorus hybrid fish that are also called platyfishswordtail hybrids.
To test the hypothesis that there are parallels between neural crest/ premelanoblast invasion and ECM changes in the early developing skin, we investigated neural crest/premelanoblast cell distribution through development and characterize their ECM environment. In this study, the last two stages of embryogenesis of platyfish-swordtail hybrids; stages 15 and 16 were used as the developmental stages of choice because we observed changes in the ectoderm, extracellular matrix (ECM) and neural crest/premelanoblast cells in the hybrid fish starting from stage 15 (25 somite stage), compared to control swordtail fish.
To determine the onset of these phenotypic changes, we first examined the development of platy fish-swordtail hybrids at one early developmental stage (stage 13; 23 somite stage; as classified by Tavolga [13]. Trunk neural crest cell migrate through two major migratory pathways, namely the dorso-lateral and ventral pathways in teleost’s, including the swordtail fishes, amniotes and amphibians [14,16,24-26]. Neural crest cells on the apex of somites may represent migrating, presumptive neural crest-derived premelanoblasts [16]. We found many of these neural crest/ presumptive premelanoblast cells at the apex of somites, and they start to invade the ectoderm of platy fishswordtail hybrids at stage 13 (Figure 1A).
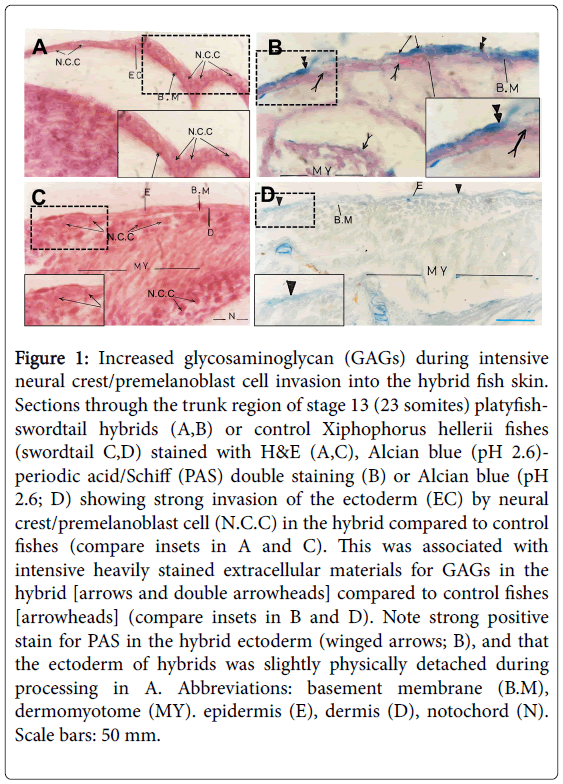
Figure 1: Increased glycosaminoglycan (GAGs) during intensive neural crest/premelanoblast cell invasion into the hybrid fish skin. Sections through the trunk region of stage 13 (23 somites) platyfishswordtail hybrids (A,B) or control Xiphophorus hellerii fishes (swordtail C,D) stained with H&E (A,C), Alcian blue (pH 2.6)- periodic acid/Schiff (PAS) double staining (B) or Alcian blue (pH 2.6; D) showing strong invasion of the ectoderm (EC) by neural crest/premelanoblast cell (N.C.C) in the hybrid compared to control fishes (compare insets in A and C). This was associated with intensive heavily stained extracellular materials for GAGs in the hybrid [arrows and double arrowheads] compared to control fishes [arrowheads] (compare insets in B and D). Note strong positive stain for PAS in the hybrid ectoderm (winged arrows; B), and that the ectoderm of hybrids was slightly physically detached during processing in A. Abbreviations: basement membrane (B.M), dermomyotome (MY). epidermis (E), dermis (D), notochord (N). Scale bars: 50 mm.
At pH 1.0, Alcian blue stains only sulfated glycosaminoglycan (GAGs)/ mucopolysaccharides, while it stains all GAGs (sulfated and non-sulfated) at pH 2.5 or 2.6 (Green and Pastewka). Trunk sections of stage 13 stained with Alcian blue (pH 2.6; stains sulfated and nonsulfated GAGs) showed extensive staining in the basement membrane and the entire ectoderm (Figure 1B). Neutral polysaccharides also existed in the ectoderm, as determined by Periodic acid-Schiff (PAS) staining (Figure 1B). Similarly, control swordtail fishes at stage 13 showed many presumptive neural crest/ presumptive premelanoblast cells at the apex of somites/dermo- myotomes, and some of them started to invade the ectoderm (Figure 1C). Alcian blue staining of these areas in control fish showed some stain in the basement membrane, dermis and epidermis, which was weak comparable to the same regions in platyfish-swordtail hybrids (Figure 1B and 1D).
During vertebrate embryogenesis, premelanoblasts undergo proliferation before migration through the somites and between the dermatome and overlying ectoderm. Then, they migrate through the developing dermis before invading the epidermis (Millar). The epidermis of teleost fish embryo consists of two characteristic cell layers: the surface layer or enveloping layer (EVL), and the inner or epidermal basal layer (EBL), which are formed at the blastula and gastrula periods, respectively, and they become the simple epithelium of the teleost fish skin [4].
Like stage 13, we observed intensive neural crest/premelanoblast cells on the apex of somites and inside the somites and their descendent dermomyotomes in the trunk region of stage 15 platyfishswordtail hybrids (25 somites; Figure 2A), as previously reported in swordtail fish [14,16].
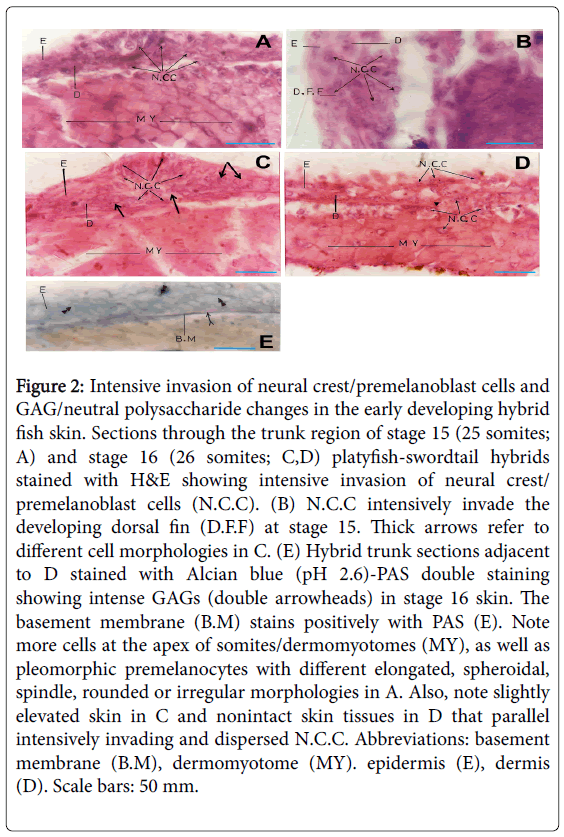
Figure 2: Intensive invasion of neural crest/premelanoblast cells and GAG/neutral polysaccharide changes in the early developing hybrid fish skin. Sections through the trunk region of stage 15 (25 somites; A) and stage 16 (26 somites; C,D) platyfish-swordtail hybrids stained with H&E showing intensive invasion of neural crest/ premelanoblast cells (N.C.C). (B) N.C.C intensively invade the developing dorsal fin (D.F.F) at stage 15. Thick arrows refer to different cell morphologies in C. (E) Hybrid trunk sections adjacent to D stained with Alcian blue (pH 2.6)-PAS double staining showing intense GAGs (double arrowheads) in stage 16 skin. The basement membrane (B.M) stains positively with PAS (E). Note more cells at the apex of somites/dermomyotomes (MY), as well as pleomorphic premelanocytes with different elongated, spheroidal, spindle, rounded or irregular morphologies in A. Also, note slightly elevated skin in C and nonintact skin tissues in D that parallel intensively invading and dispersed N.C.C. Abbreviations: basement membrane (B.M), dermomyotome (MY). epidermis (E), dermis (D). Scale bars: 50 mm.
In stage 15 platyfish-swordtail hybrids, neural crest/premelanoblast cells were intensively invading both the epidermis and dermis and were characteristically randomly distributed (Figure 2A). In addition, these neural crest cells were invading the somites/ dermomyotomes and were intensive on the apex of somites, as well as had a characteristic pleomorphic shape with different elongated, spheroidal, spindle-like, rounded or irregular morphologies (Figure 2A). Similar intensive invasion and morphological changes were observed in the epidermis and dermis of the developing dorsal fin (Figure 2B).
Similarly, the developing skin of platyfish-swordtail hybrid fish at stage 16 (26 somites; as classified by Tavolga [13] showed an intensive invasion of neural crest/premelanoblast cells (Figure 2C and 2D), with a characteristically slightly elevated skin (Figure 2C), or nonintact skin tissues (Figure 2D) at some areas of the trunk, which parallels intensively invading and dispersed neural crest/premelanoblast cells.
In adjacent stage 16 trunk sections processed for Alcian blue (pH 2.6) Periodic acid-Schiff (PAS) double staining, both sulfated and nonsulfated GAGs were intense in the dermis and epidermis, while the basement membrane appears to stain more intensely for neutral polysaccharides (Figure 2E). Staining of sections of the same region with Alcian blue at pH 1.0 (stains only sulfated GAGs) reveals a very weak staining in the basement membrane, dermis and epidermis (Figure 3A and 3B), suggesting that these hybrid skin regions contain non-sulfated rather than sulfated GAGs. Interestingly, intense collagen fiber stain was observed in these skin regions, as shown by van Gieson’s stain (Figure 3C). Notably, Alcian blue staining of control swordtail fishes show some weak stains in the basement membrane, dermis and epidermis, compared to platyfish-swordtail hybrids (Figure 3D).
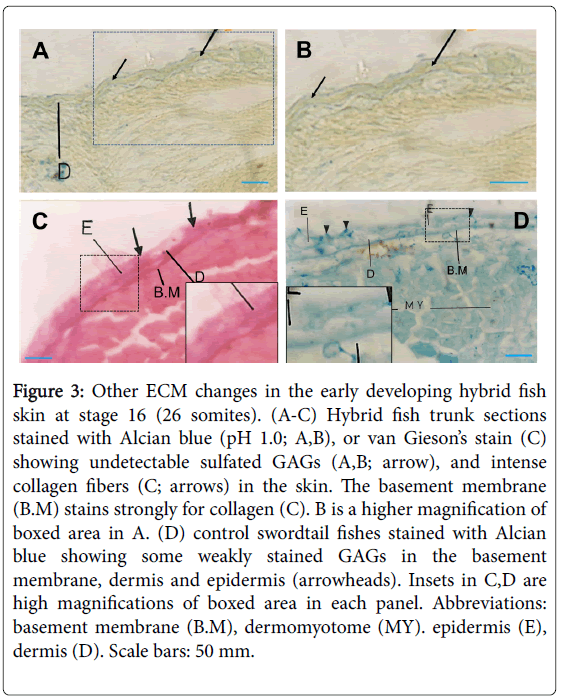
Figure 3: Other ECM changes in the early developing hybrid fish skin at stage 16 (26 somites). (A-C) Hybrid fish trunk sections stained with Alcian blue (pH 1.0; A,B), or van Gieson’s stain (C) showing undetectable sulfated GAGs (A,B; arrow), and intense collagen fibers (C; arrows) in the skin. The basement membrane (B.M) stains strongly for collagen (C). B is a higher magnification of boxed area in A. (D) control swordtail fishes stained with Alcian blue showing some weakly stained GAGs in the basement membrane, dermis and epidermis (arrowheads). Insets in C,D are high magnifications of boxed area in each panel. Abbreviations: basement membrane (B.M), dermomyotome (MY). epidermis (E), dermis (D). Scale bars: 50 mm.
Cryofractured specimens, which were prepared by removing separating the epidermis from the dermis to further analyze the underside of the epidermal tissue, were examined by SEM to confirm both the histological and histochemical data in platyfish-swordtail hybrid skin (Figure 4A-4C). They revealed that these neural crest/ premelanoblast cells exhibited their previously described characteristic morphology in Xiphophorus fish embryos [14] and other vertebrate embryos [27]. Thus they were pleomorphic with different elongated, spheroidal, spindle-like, flattened or irregular morphologies, and had intensive cell-ECM fibril contacts (Figure 4A and 4B), as previously described in swordtail/platyfish [14].
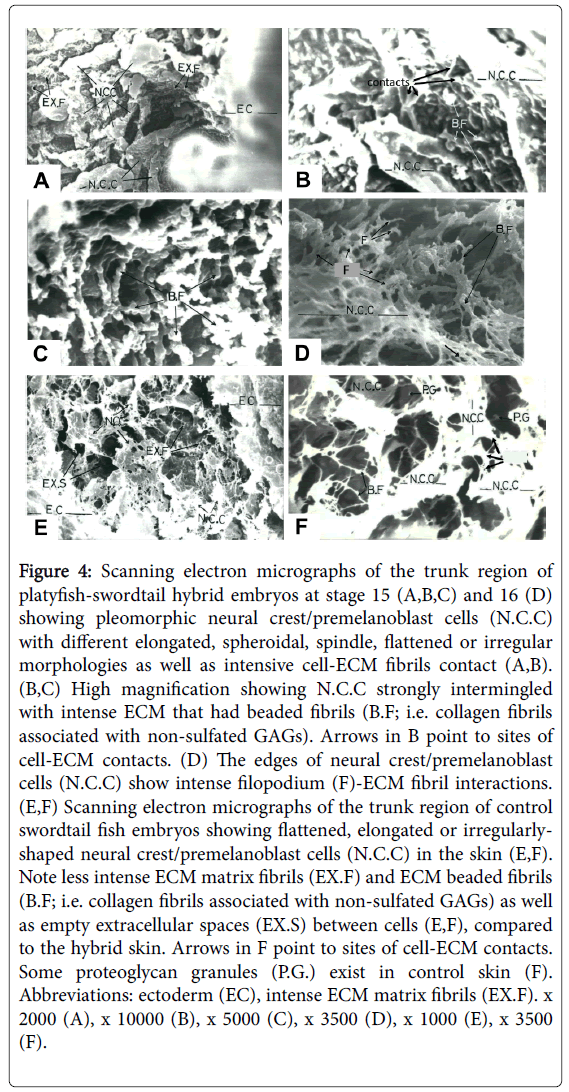
Figure 4: Scanning electron micrographs of the trunk region of platyfish-swordtail hybrid embryos at stage 15 (A,B,C) and 16 (D) showing pleomorphic neural crest/premelanoblast cells (N.C.C) with different elongated, spheroidal, spindle, flattened or irregular morphologies as well as intensive cell-ECM fibrils contact (A,B). (B,C) High magnification showing N.C.C strongly intermingled with intense ECM that had beaded fibrils (B.F; i.e. collagen fibrils associated with non-sulfated GAGs). Arrows in B point to sites of cell-ECM contacts. (D) The edges of neural crest/premelanoblast cells (N.C.C) show intense filopodium (F)-ECM fibril interactions. (E,F) Scanning electron micrographs of the trunk region of control swordtail fish embryos showing flattened, elongated or irregularlyshaped neural crest/premelanoblast cells (N.C.C) in the skin (E,F). Note less intense ECM matrix fibrils (EX.F) and ECM beaded fibrils (B.F; i.e. collagen fibrils associated with non-sulfated GAGs) as well as empty extracellular spaces (EX.S) between cells (E,F), compared to the hybrid skin. Arrows in F point to sites of cell-ECM contacts. Some proteoglycan granules (P.G.) exist in control skin (F). Abbreviations: ectoderm (EC), intense ECM matrix fibrils (EX.F). x 2000 (A), x 10000 (B), x 5000 (C), x 3500 (D), x 1000 (E), x 3500 (F).
Furthermore, these neural crest premelanoblast cells were strongly intermingled with intense ECM (Figure 4A,4B,4D) that mostly had beaded fibrils (Figure 4B and 4C). ECM beaded fibrils have been previously identified as collagen fibrils associated with non-sulfated GAGs [15,17,22]. In addition, we observed few proteoglycans granules (Figure 4B), which have been previously identified as sulfated GAGs [15,17,22]. Interestingly, control swordtail fishes at the same stage showed many flattened, elongated or irregularly-shaped neural crest/ premelanoblast cells in the skin (Figure 4E and 4F). However, ECM beaded fibrils (collagen fibrils associated with non-sulfated GAGs) were apparently less intense in control fish skin (Figure 4E and 4F), compared to the same region in platyfish-swordtail hybrid skin (compare Figure 4E with 4A, and Figure 4F with 4B and 4C). Remarkable intense filopodium-ECM fibril interactions at the edges of neural crest/premelanoblast cells were observed at stage 16 (Figure 4D). Taken together, these data suggest a correlation between the intensive invasion of neural crest/premelanoblast cells in the dermis and epidermis with the increase of both collagen fibrils and nonsulfated GAGs in platyfish-swordtail fish hybrids.
Neural crest origin of melanoblasts, premelanocytes and melanocytes
Vertebrate skin melanocytes are derived from the neural crest, reside in epidermal and dermal layers and synthesize melanins in the skin. During embryogenesis, neural crest cells undergo an epithelio mesenchymal transition to yield migratory cells dispersing along specific pathways, then they differentiate into various cell types, including neurons, glial cells of the nervous system, endocrine cells and melanocytes [28,29]. Several studies have shown that murine melanoblasts undergo proliferation before migration through the somites and between the dermatome and overlying ectoderm. Then, they migrate through the developing dermis before invading the epidermis during embryogenesis [1,30]. Vertebrate neural cell markers studies have helped to identify and trace the various NC-derived cell types, including melanocytes and their melanoblast progenitors which are unpigmented, but have the potential to produce melanin. For instance, at the trunk level of the quill, NC cell migration proceeds in two definitive pathways. NC cells migrate dorsoventrally, near the neural tube within the anterior part of the somites and differentiate to peripheral ganglia and nerves. NC cells that migrate mediolaterally between the superficial ectoderm and the dermomyotome give rise to melanocyte precursors, which invade the subectodermal mesenchyme [30-33].
The role of ECM in NCC and premelanocytes, melanoblast migration and invasion
Two types of molecules could function as positive regulators of NCC and premelanocyte/melanoblast migration: permissive factors such as ECM molecules that promote migration and motility, chemoattractants that drive NCCs to specific locations. The multivalent interactions of NCCs and premelanocyte/melanoblasts with their surrounding ECM molecules such as sulfated and nonsulfated GAGs, collagen, laminin and fibronectin are essential for the guidance of their migration and development [26.27]. In Xiphophorus and other vertebrate embryos, the level of expression of sulfated GAGs (e.g.; chondroitin sulfate proteoglycans), fibronectin and hyaluronic acid (a non-sulfated GAG) increases dramatically in NC streams to facilitate their migration and, then, decreases after migration was nearly completed [25,27,34].
Several In vivo and in vitro studies on the potential function of ECM molecules as migration-promoting factors have helped to classify ECM into three distinct categories: permissive category, including ECM molecules supporting extensive NCC attachment and migration such as sulfated GAGs (chondroitin- sulfate proteoglycans), some types of collagens, fibronectin and laminin; non-permissive category that promotes a weak cell adhesion, without sustaining a significant cell locomotion such as non-sulfated GAGs (hyaluronic acid); and inhibitory category that directly impedes NCC movement such as Aggrecan [27]. Indeed, experiments involving dermomyotomal ablations and injection of target complexes of GAG-degrading enzymes into the subectodermal region that inhibits sulfated GAGs have shown great defects in NCC migration [27]. Yet, little is known about the histochemical and biochemical changes that accompany the very early stages of the invasion of neural crest cells/premelanoblasts into the developing skin of Xiphophorus hybrid (platyfish-swordtail) fish. Here, we investigated the distribution and invasion of neural crest/premelanoblast cells and characterize their ECM environment in the developing skin at chosen embryonic stages of platyfish-swordtail fish hybrids. We have also tested the hypothesis that there are parallels between neural crest/premelanoblast invasion and ECM changes in the early developing skin in platyfish-swordtail fish hybrids.
The results reported here provide several histochemical evidences for the correlation between the spatiotemporal relationship of ECM components and both the migration and invasion of neural crest/ premelanoblast cells into the developing skin of platyfish-swordtail hybrids, suggesting that these ECM molecules are involved in the intensive invasion of these cells into the skin. One possible mechanism is that hyaluronic acid (a non-sulfated GAG) may facilitate the invasion of neural crest/premelanoblast cells into the dermis and epidermis. As a non-permissive ECM, the hyaluronic acid may promote cell locomotion by enhancing the intercellular spaces and by the inhibition of extensive intercellular interactions by physically separating cells [27,34,35]. This is supported with early studies showing that injection of early chick embryos with hyaluronidase before a short in ovo incubation results in a marked loss of intercellular space, which indicates a role for hyaluronic acid in establishing and maintaining spaces during embryonic development [36]. Indeed, we found that the hyaluronic acid remarkably increases in the developing platyfish-swordtail hybrid skin, compared to controls during the intensive invasion of neural crest/premelanoblast cells (compare Figure 2E and 3A, 3B with Figure 3D). This was correlated with apparent cell separations and the creation of wide intercellular spaces between the invading neural crest/premelanoblast cells in the skin (Figure 2D). In addition, the strong increase of collagen fibrils (Figure 3C and Figure 4), which are known permissive ECM components, may provoke the invasion and interspersion of neural crest/premelanoblast cells in the developing platyfish-swordtail hybrid skin, as in other organs [37-39] as well as in other systems in which collagen directly stimulates cell migration and invasion [40,41]. Taken together, our data suggest that there are parallels between neural crest/premelanoblast invasion and ECM changes in the early developing skin in platyfish-swordtail fish hybrids.
Xiphophorus interspecies hybrids as a model for spontaneous melanoma
As a well-established model organism, the fish genus Xiphophorus has been extensively used to investigate melanoma formation [11,12,42,43]. Wild-type Xiphophorine fish are insusceptible of developing neoplasia. However, melanomas could occur in certain backcross fish as the result of specific hybrid matings that minimize the influence of environmental factors in genetic studies. Xiphophorus interspecies hybrids have provided well-developed genetically controlled models of melanoma formation. A `two-gene' spontaneous melanoma model called ‘Gordon-Kosswig' has been established to explain melanoma formation in hybrid fish. These hybrid fish are created by backcrossing swordtail (Xiphophorus helleri ) with platyfish (Xiphophorus maculatus ) as the recurrent parent, producing offspring that develop melanoma in a Mendelian fashion (Anders, 1991; Schartl, 1995; Vielkind et al., 1989). Thus, spontaneous melanomas could form in first-generation backcross hybrids [BC(1)] that are produced from backcrossing F(1) hybrids derived from the swordtail X. helleri and the platyfish X . maculatus to the X. helleri parental strain (the Gordon- Kosswig hybrid cross). These hybrids develop melanosis at different ages, and thus spontaneous melanoma are found in the hybrid progeny of BC1 mated to F1 hybrids at early (newly hatched or fry stage) and old ages, which is called the fry and adult melanomas, respectively [43-45]. Our current data suggesting that there are parallels between neural crest/premelanoblast invasion and ECM changes in the early developing skin in platyfish-swordtail fish hybrids, may therefore help in understanding the mechanisms that regulate early stages of melanoma development in this fish, in addition to early skin development. They also provide a conceptual framework for future mechanistic studies on the biochemical, cellular and molecular changes that accompany the early stages of melanoma formation in platyfish-swordtail hybrids [46].