Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Research Article - (2012) Volume 1, Issue 7
Diallyl disulfide (DADS) exerts numerous anticancer effects, involving multiple molecular mechanisms. In particular, DADS has been revealed as a potential inhibitor attenuating histone deacetylase (HDAC) activity, which could aid cancer prevention and therapy by inducing histone hyperacetylation and consequently reactivating epigenetically silenced tumor suppressor genes involved in cancer initiation and progression. In an in vitro study, we demonstrated that DADS increased histones H3 and H4 acetylation in human gastric cancer MGC803 cells in a time-dependent fashion, accompanied by increased p21WAF1 protein levels, which was consistent with G2/M phase cell cycle arrest. DADS
also demonstrated dose-dependent antitumor effects in MGC803-xenografted nude mice in vivo, resulting in tumor cells growth inhibition and G2/M phase arrest. Acetylated histones H3 and H4 were up-regulated by administration of DADS in association with elevated p21WAF1 protein expression. There was no evidence of gross toxicity associated with DADS treatment. DADS also preferentially enhanced acetylation of histone H3 relative to histone H4 both in vivo and in vitro. In addition, characteristic features of cell differentiation were observed in xenografted tumor cells. These results suggest that DADS can cause cell cycle arrest and inhibit cell proliferation by inducing hyperacetylation of histones
H3 and H4 and increasing p21WAF1 expression in MGC803 cells, which may partly account for its antitumor effects in gastric cancer. These results indicate that DADS may be useful for gastric cancer prevention, and may have important implications for epigenetic therapy by virtue of its ability to modulate histone acetylation.
Keywords: Diallyl disulfide; Histone acetylation; P21WAF1; Gastric cancer
DADS: Diallyl Disulfide; HDAC: Histone Deacetylase; HAT: Histone Acetylase; SB: Sodium Butyrate; NS: Normal Saline; ALT: Alanine Aminotransferase; AST: Aspartate Aminotransferase; BUN: Blood Urea Nitrogen; SCR: Serum Creatinine; PBS: Phosphate Buffered Saline; SDS: Sodium Dodecyl Sulfate; PCNA: Proliferating Cell Nuclear Antigen
Gastric cancer is a frequently encountered tumor and the second most common cause of cancer deaths worldwide, accounting for almost two-thirds of the cases occurring in developing countries and 42% in China alone. In 2008, over 70% of new cases and deaths were in developing countries. The 5-year survival rate for stomach cancer is still relatively low compared with most other cancers [1,2]. Although the total mortality has declined remarkably in China during the past decades, it remains high in rural areas [3]. The global burden of gastric cancer thus remains one of the key issues in cancer prevention worldwide.
Diallyl disulfide (DADS) is a major organosulfur constituent derived from garlic and a few other vegetables of the Allium family [4,5]. Epidemiological and laboratory investigations have shown that garlic and its constituents have anticancer effects manifesting as reduced cancer risk, protection against chemically-induced cancer in animal models, and altered biological behavior of tumors. Notably, convincing data have revealed that DADS can exhibit a variety of anticancer effects in diverse cancers by modulating carcinogen metabolism, retarding cell cycle progression, inhibiting cell proliferation, and inducing cell differentiation or apoptosis [6,7].
Studies of its molecular mechanisms have documented that DADS produces its anticancer effects by targeting multiple signaling pathways [7]. For example, DADS can directly or indirectly modify the metabolism of reactive oxygen species through the redox-based mechanisms, consequently inducing apoptosis or cell cycle arrest mediated by JNK or other specific signaling pathways, depending on the treatment concentration or tumor cell type [8]. In particular, DADS can induce cancer cell differentiation or inhibit proliferation by increasing histone acetylation through inhibition of histone deacetylase (HDAC) activity, and DADS has been suggested as a potential agent for modulating the dysregulated status of histone modification in cancer cells [9]. This has increased interest in further elucidating the role of DADS in epigenetic modulation.
Deregulation of histone deacetylation is one of the main events in the epigenetic alterations observed in cancer development and progression, and may result in aberrant gene silencing associated with uncontrolled proliferation, dedifferentiation, escape from apoptosis, and promotion of carcinogenesis. The balance between acetylation and deacetylation of histones is maintained by the opposing activities of HDACs and histone acetylases (HATs). HDAC overactivation and/ or HAT inactivation results in hypoacetylation or loss of histone acetylation, especially of specific residues in the core histones H3 and H4, which has been identified as a hallmark of cancer cells [10]. Recent evidence has shown that inhibition of HDAC activity can increase acetylated histones H3 and H4 levels on the P21WAF1 promoter region and increase P21WAF1 transcription, resulting in growth suppression in human skin cancer and non-small cell lung cancer [11,12]. P21WAF1 is a major cyclin-dependent kinase inhibitor involved in cell cycle control. Up-regulation of P21WAF1 expression can induce cell cycle arrest, which facilitates tumor growth arrest [13-17].
Our previous studies showed that DADS inhibited the proliferation of gastric cancer cells by causing G2/M phase cell cycle arrest through activation of p38 [18,19] or by interference with the ATR/Chk1/ Cdc25C/cyclinB1 signaling pathway [20,21]. DADS also induced differentiation by down-regulating the ERK signaling pathway [22]. It has been proposed that the differentiation of human leukemia cells and the growth inhibition of human breast and colon cancer cells induced by DADS were caused by increased histone acetylation [9,23]. Moreover, DADS-induced G2/M phase cell cycle arrest was associated with hyperacetylation of histones H3 and H4 in human colon tumor cells, concomitant with the increased expression of P21WAF1 [23,24]. We therefore speculated that DADS might exert similar effects in gastric cancer, which may represent an important epigenetic mechanism of DADS action, which could contribute to gastric tumor prevention and therapy.
We therefore examined the effects of DADS on the acetylation of histone H3 and H4, and on P21WAF1 expression in the human gastric cancer MGC803 cell line both In vitro and In vivo. In addition, we studied the effects of DADS on cell cycle progression and growth of xenografts In vivo and systematically explored DADS toxicity in xenograft models.
Chemicals and reagents
DADS (purity 80%, the remaining 20% being diallyl trisulfide and diallyl sulfide), purchased from Fluka Co., (Milwaukee, Wisconsin, USA), was dissolved in Tween 80 at 8 g/L and stored at -20°C. Sodium butyrate (SB), purchased from Sigma Co., (St Louis, Montana, USA), was dissolved in PBS at 11 g/L and stored at -20°C, and it was used as the positive control. Normal saline (NS) was used as the negative control. Anti-acetylated histone H3 and H4 rabbit polyclonal primary antibodies and horseradish peroxidase (HRP) conjugated anti-rabbit secondary antibody was purchased from Upstate Biotechnology, Inc. (Lake Placid, NY, USA). Mouse monoclonal antibodies against P21WAF1, proliferating cell nuclear antigen (PCNA) and HRPconjugated anti-mouse secondary antibody were obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, California, USA). The mouse monoclonal against human β-actin antibody was purchased Sigma Biotechnology, Inc. (Sigma-Aldrich, St Louis, Montana, USA).
Cell culture
Human gastric cancer cell line MGC803 was obtained from the Cancer Research Institute, Xiangya Medical College, and Central South University in China. Cells were cultured in RPMI-1640 medium (Gibco, Life Technologies, Vienna, Austria) containing 10% fetal bovine serum (Sijiqing Biological Engineering Material Co., Ltd., Hangzhou, China) with addition of 100 U/mL penicillin, 100 U/mL streptomycin, and maintained at 37°C in a humidified atmosphere of containing 5% CO2.
Animals
Male athymic BALB/c nude mice (3-4 weeks old) were purchased from the Experimental Center of the Chinese Academy of Science in Shanghai. The mice were housed in an environment controlled for temperature (22 ± 2°C), light (12 h light/dark cycles) and humidity (60 ± 10%). The animals were maintained under specific pathogenfree conditions in accordance with NIH Guide for the Care and Use of laboratory Animals.
Gastric tumor growth in nude mice
2×106 MGC803 cells in the logarithmic growth phase were injected into the subcutis of the right thigh per mouse. Fourteen days after inoculation, the human gastric cancer models were established when the transplanted solid tumor volumes of BALB/c nude mice reached around 100 mm3. Tumor volume was calculated using the standard formula (width2 ×length×0.5) as described previously [25]. The animals were randomly divided into five groups, and each group consisted of four mice. The mice were treated with normal saline, DADS, or sodium butyrate via intraperitoneal injection for three weeks. Thereafter, injections, three times a week, were continued until the termination of the experiment. The mice were weighed thrice weekly. At the termination of the experiment, all mice were sacrificed by cervical dislocation. The sizes of the locally growing tumors were determined. Tumor tissues were then flash frozen and stored in liquid nitrogen until applied to protein extraction or fixed in formalin and embedded in paraffin. The experiments were carried out according to the guidelines for animal use, the Ethics Committee of University of South China.
Serum biochemical analysis and histopathological examination
For analysis of DADS toxicity In vivo, the mice (n=4 per group) were treated with normal saline, DADS (50, 100, and 200 mg/kg) via intraperitoneal injection. At the termination of the experiment, blood was collected from the abdominal caval vein after anesthesia and an abdominal incision. After centrifugation, serum samples were collected and stored in -80°C until analysis. Blood chemistry levels of alanine aminotransferase (ALT), aspartate aminotransferase (AST), blood urea nitrogen (BUN), and serum creatinine (SCR) were determined using Sigma Diagnostic Kits (Sigma Chemical Co., St. Louis, MO). For histopathological examinations, selected organs (heart, liver, spleen, kidney, testes, and femoral bone marrow) were removed from the mice that received normal saline or DADS (100 mg/kg), and fixed in 10% formalin. The organs were prepared as paraffin-embedded glass slides stained with hematoxylin and eosin. Morphological changes were observed under microscope.
Cycle distribution analysis
Cells cultured In vitro untreated or treated with DADS (15 and 30 mg/L) were harvested and resuspended in 1 mL of ice-cold 75% ethanol and fixed for 24 h at 4°C. For the xenograft tumor cells, the mice were treated and tumor tissues were collected as described above. 0.1 g tumor tissues were cut into mono-cell suspension in ice-cold PBS. After centrifugation, cells were resuspended in 1 mL of ice-cold 75% ethanol and fixed for 24 h at 4°C. For subsequent flow cytometry analysis, fixed cells were washed with PBS once and resuspended in 1mL of PI staining reagents (20 mg/L ribonuclease and 50 mg/L propidium iodide). After the samples were incubated in the dark for 30 min, the data were collected using a FACS can flow cytometer (Becton Dickinson, Franklin Lakes, USA) and analyzed with Verity Winlist Software (Verity Software House, Topsham, ME, USA).
Protein extraction
For total protein extraction, cells cultured In vitro were washed with ice-cold PBS and then lysed directly on ice for 30 min in 0.5 mL lysis buffer [10 mmol/L Tris-HCl (pH 7.6), 100 mmol/L NaCl, 1 mmol/L EDTA (pH 8.0), 100 μg/mL PMSF and 1 μg/mL aprotinin]. Frozen tumor tissues (100 mg) from each group were ground under liquid nitrogen and washed in ice-cold PBS. After centrifugation, the tissue cells were centrifugated and resuspended in 0.5 mL lysis buffer as described above. The cell lysates were centrifugated at 12,000 rpm for 10 min and the supernatants were collected. Then protein contents were determined using a BCA protein assay kit (Pierce, Rockford, IL, USA).
Western blot analysis
Total proteins (30 μg) were loaded on a 12% SDS-polyacrylamide gel for electrophoresis and transferred onto Polyvinylidene Fluoride (PVDF) membrane. The blots were blocked in 5% skim milk in Tris buffered saline (TBS) containing 0.1% Tween 20 for 2 h at room temperature, and then incubated for 2 h at room temperature with the 1:1000 dilution of the acetylated histones H3 and H4 rabbit polyclonal primary antibodies and with the 1:400 dilution of mouse monoclonal anti-P21WAF1 primary antibody. The membranes were washed in TBS-T and then incubated with a 1:1000 dilution of HRP-conjugated secondary antibody for 1 h at room temperature. After washed with TBS-T, the membranes were developed by an enhanced chemiluminescence plus (ECL Plus) kit (Amersham Biosciences, Buckinghamshire, England) and bands were visualized on X-ray film (Kodak). To verify equal protein-sample loading, the membranes were re-incubated with anti- β-actin mouse monoclonal antibody. The target protein amounts were normalized towards β-actin quantity using densitometry and relative fold changes in protein levels were then calculated as ratios between treated versus control group values.
Immunohistochemistry
A representative 5-μm tissue section was cut from paraffinembedded sample from xenograft mice of human gastric cancer that received treatment with normal saline or 100 mg/kg DADS. The slides were H&E stained to observe morphology changes of gastric tumor cells. For analysis of P21WAF1 and PCNA expression, immunohistochemistry was performed as follows. Sections were deparaffinized in xylene and hydrated in graded alcohols, followed by antigen retrieval by heat treatment in 10 mM sodium citrate buffer (pH 8.0). Slides were incubated in 3% H2O2 in ethanol to quench the endogenous peroxidase, and then incubated with normal goat serum for 20 min. Sections were incubated with primary antibodies for P21WAF1 and PCNA (dilution 1:100) at 4°C overnight. Positive signals were developed with peroxidase-conjugated secondary antibody using 0.5% diaminobenzidine/H2O2 followed by counter staining with Mayer's hematoxylin, dehydration, clearing, and mounting. The slides treated with normal goat serum were evaluated as negative controls.
Statistical analysis
All results are presented as mean ± SD for three independent experiments. Statistical significance (P<0.05) was determined using Student’s t test.
DADS increases the acetylation levels of histones H3 and H4 in MGC803 cells In vitro
DADS has been shown to induce hyperacetylation of histones H3 and H4 in diverse tumors [9,23]. We performed In vitro experiments to determine if DADS also exerted this effect in human gastric cancer. Human gastric cancer MGC803 cells were incubated in medium alone, in medium containing 15, 30 and 60 mg/L DADS, or in DADS (30 mg/ L)+sodium butyrate (550 mg/L) for 12 h. Sodium butyrate is a known dietary HDAC inhibitor [23,26] and was used as a positive control in this study. The effects of DADS on the acetylation statuses of histones H3 and H4 were analyzed by western blotting. As shown in Figure 1A and 1B, DADS induced hyperacetylation of histone H3 after 12 h incubation. The greatest effect was achieved with 30 mg/L DADS, which produced a 1.7-fold increase compared with the other concentrations. DADS induced concentration-dependent hyperacetylation of histone H4, with the greatest effect produced by 60 mg/L DADS with a 1.5-fold increase (Figure 1C and 1D). Moreover, DADS had a greater influence on the acetylation of histone H3 compared with histone H4. We then treated cells with 30 mg/L DADS for different times. As shown in Figure 1E and 1F, histone H3 acetylation increased significantly after 6 h and was maintained up to 24 h, but then decreased slightly at 24 h. Compared with histone H3, histone H4 acetylation increased slightly at 6 h, and was increased evidently after 12 h. These data suggest that DADS-induced histone acetylation may be more effective after longer incubation times or at higher concentrations.
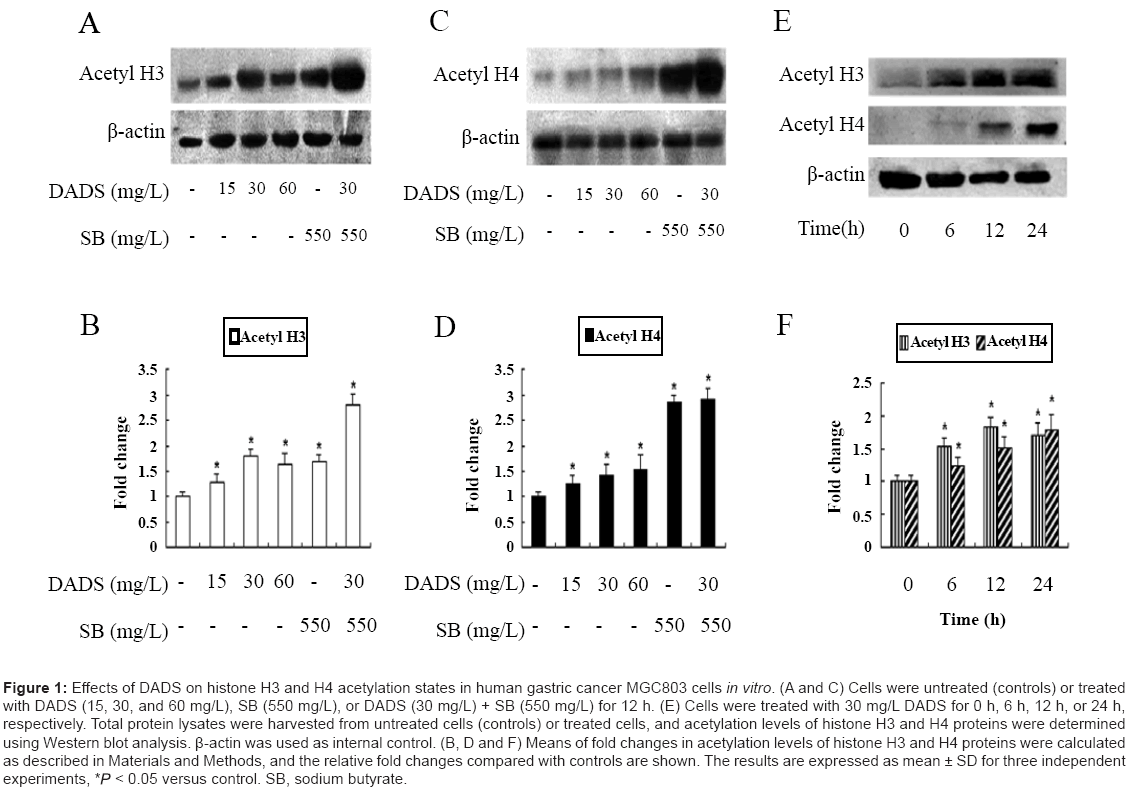
Figure 1: Effects of DADS on histone H3 and H4 acetylation states in human gastric cancer MGC803 cells in vitro. (A and C) Cells were untreated (controls) or treated with DADS (15, 30, and 60 mg/L), SB (550 mg/L), or DADS (30 mg/L) + SB (550 mg/L) for 12 h. (E) Cells were treated with 30 mg/L DADS for 0 h, 6 h, 12 h, or 24 h, respectively. Total protein lysates were harvested from untreated cells (controls) or treated cells, and acetylation levels of histone H3 and H4 proteins were determined using Western blot analysis. β-actin was used as internal control. (B, D and F) Means of fold changes in acetylation levels of histone H3 and H4 proteins were calculated as described in Materials and Methods, and the relative fold changes compared with controls are shown. The results are expressed as mean ± SD for three independent experiments, * < 0.05 versus control. SB, sodium butyrate.
DADS increases P21WAF1 protein expression in MGC803 cells In vitro
Hyperacetylation of histones H3 and H4 induced by DADS has been suggested to cause up-regulation of P21WAF1 mRNA and protein levels as one of its mechanisms, accounting for G2/M phase cell cycle arrest in human colon tumor cell lines [23]. We therefore detected P21WAF1 protein levels by western blot analysis after treating MGC803 cells with different concentrations of DADS for 12 or 24 h. P21WAF1 protein levels were up-regulated in MGC803 cells after incubation with 15, 30, and 60 mg/L DADS, in a time-dependent manner. The greatest increase occurred after incubation with 30 mg/L for 24 h, which resulted in a 2.6-fold increase above the control level (Figure 2A and 2B). These data suggest that MGC803 cells In vitro were more sensitive to 30 mg/L DADS than to other concentrations, and that induction of P21WAF1 expression by DADS may depend on incubation time.
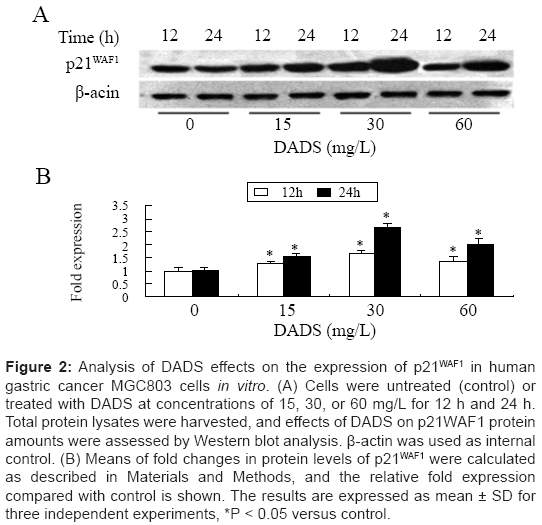
Figure 2: Analysis of DADS effects on the expression of p21WAF1 in human gastric cancer MGC803 cells in vitro. (A) Cells were untreated (control) or treated with DADS at concentrations of 15, 30, or 60 mg/L for 12 h and 24 h. Total protein lysates were harvested, and effects of DADS on p21WAF1 protein amounts were assessed by Western blot analysis. β-actin was used as internal control. (B) Means of fold changes in protein levels of p21WAF1 were calculated as described in Materials and Methods, and the relative fold expression compared with control is shown. The results are expressed as mean ± SD for three independent experiments, *P < 0.05 versus control.
DADS induces G2/M phase cell cycle arrest in MGC803 cells In vitro
To explore the correlation between DADS-induced P21WAF1 expression and cell cycle progression, we analyzed the cell cycle distribution of MGC803 cells by flow cytometry after incubation with 15 and 30 mg/L DADS for 12 and 24 h. As shown in Figure 3, DADS caused G2 phase arrest in concentration- and time-dependent manners. The percentages of G2 phase cells were 13% and 25%, respectively, after treatment with 15 and 30 mg/L DADS for 12 h. The equivalent percentages were 25.8% and 51.5%, respectively, after treatment with the same concentrations for 24 h. These effects were consistent with increased acetylated histone and P21WAF1 expression (Figure 1 and Figure 2) and suggest that DADS-induced G2/M phase cell cycle arrest may result from up-regulation of P21WAF1 expression via hyperacetylation of histones H3 and H4. The results of these In vitro experiments imply that the anti-gastric-tumor effects of DADS may involve modulation of histone acetylation leading to cell cycle arrest.
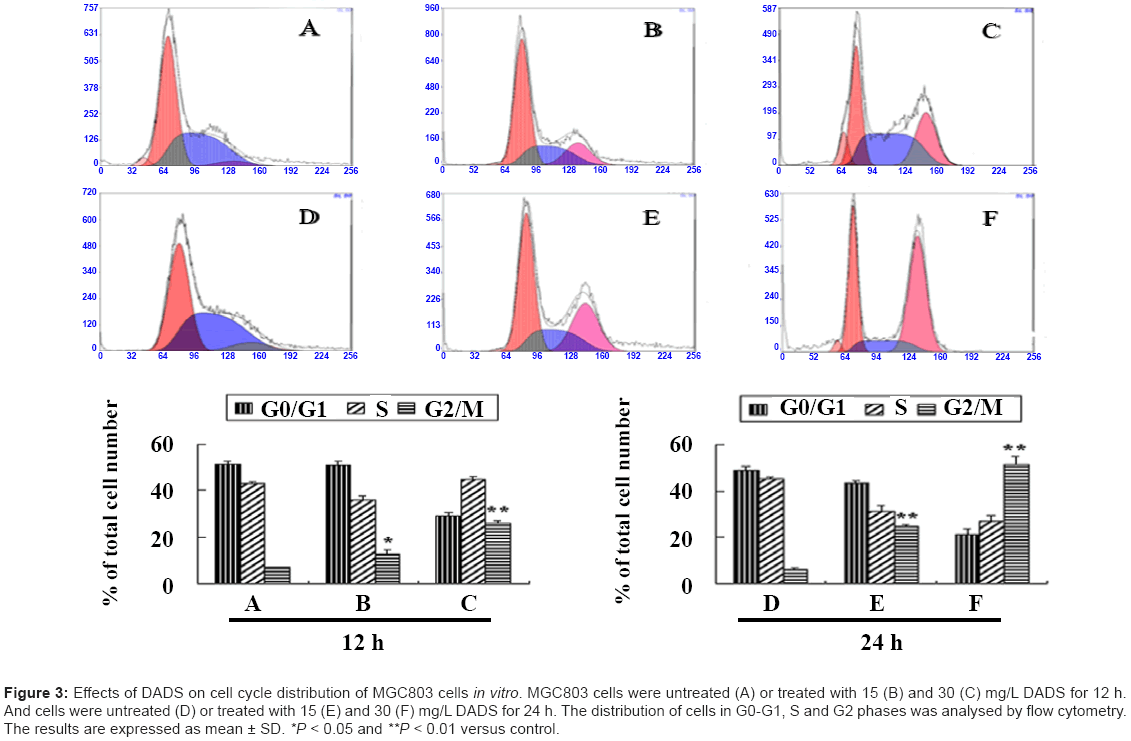
Figure 3: Effects of DADS on cell cycle distribution of MGC803 cells in vitro. MGC803 cells were untreated (A) or treated with 15 (B) and 30 (C) mg/L DADS for 12 h. And cells were untreated (D) or treated with 15 (E) and 30 (F) mg/L DADS for 24 h. The distribution of cells in G0-G1, S and G2 phases was analysed by flow cytometry. The results are expressed as mean ± SD. *P < 0.05 and **P < 0.01 versus control.
Antitumor effects of DADS in an animal model of human gastric cancer
We performed In vivo experiments to determine the ability of DADS to inhibit gastric tumor growth In vivo, and the possible association of this anticancer effect with up-regulation of acetylated histone and P21WAF1 expression. MGC803 cells were injected into nude mice. We investigated the dose-response effects by administering DADS at 50, 100, and 200 mg/kg via intraperitoneal injection three times a week for three consecutive weeks. Normal saline and sodium butyrate (66 mg/kg) were injected as negative and positive controls, respectively. As shown in Figure 4A and 4B, DADS administration exerted a significant antitumor effect, as did sodium butyrate. A detectable reduction in tumor size was observed in mice treated with DADS or sodium butyrate. Treatment with DADS demonstrated dose-dependent antitumor activity, with gradual decrease of tumor size associated with increasing DADS dose (Figure 4C). The inhibition rates in terms of tumor volume were 17.4%, 57.4% and 71.6% at DADS doses of 50, 100 and 200 mg/kg, respectively, compared with normal saline-treated control groups. No significant differences in body weights were observed in the mice treated with DADS compared with negative control mice (Figure 4D). These results suggest that DADS had no gross toxic effect on xenografted mice at the doses given.
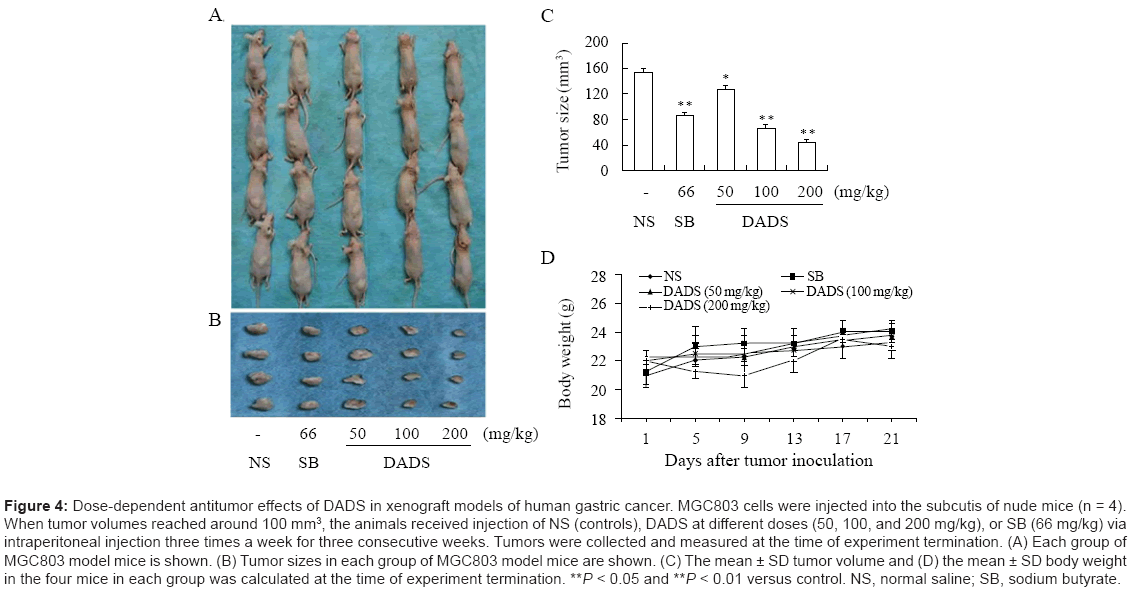
Figure 4: Dose-dependent antitumor effects of DADS in xenograft models of human gastric cancer. MGC803 cells were injected into the subcutis of nude mice (n = 4). When tumor volumes reached around 100 mm3, the animals received injection of NS (controls), DADS at different doses (50, 100, and 200 mg/kg), or SB (66 mg/kg) via intraperitoneal injection three times a week for three consecutive weeks. Tumors were collected and measured at the time of experiment termination. (A) Each group of MGC803 model mice is shown. (B) Tumor sizes in each group of MGC803 model mice are shown. (C) The mean ± SD tumor volume and (D) the mean ± SD body weight in the four mice in each group was calculated at the time of experiment termination. **P < 0.05 and **P < 0.01 versus control. NS, normal saline; SB, sodium butyrate.
Serum biochemical analysis and histopathological examination
To further determine if DADS exerted toxic effects in xenografted mice, we performed morphological examinations of selected organs including heart, liver, spleen, kidney, testes, and femoral bone marrow in mice treated with DADS. As shown in Figure 5, there was no evidence of kidney damage in DADS-treated mice, including tubule eosinophilia, degeneration, or necrosis. Likewise, no liver lesions, such as fatty change, infiltration of neutrophils, or hepatocellular necrosis were observed by microscopy. The remaining tissues (heart, spleen, testes, and femoral bone marrow) also appeared normal. Serum biochemical data indicated no effects of DADS on the levels of ALT, AST, BUN and SCR, compared with those in normal controls (Table 1). Treatment with DADS at low or high dose thus resulted in growth inhibition of gastric tumors In vivo with no obvious side effects.
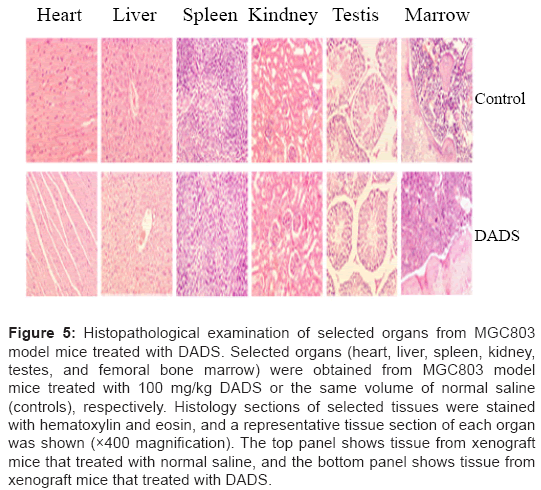
Figure 5: Histopathological examination of selected organs from MGC803 model mice treated with DADS. Selected organs (heart, liver, spleen, kidney, testes, and femoral bone marrow) were obtained from MGC803 model mice treated with 100 mg/kg DADS or the same volume of normal saline (controls), respectively. Histology sections of selected tissues were stained with hematoxylin and eosin, and a representative tissue section of each organ was shown (×400 magnification). The top panel shows tissue from xenograft mice that treated with normal saline, and the bottom panel shows tissue from xenograft mice that treated with DADS.
| Group | ALT (μ/l) | AST (μ/l) | BUN (mg/dl) | SCR (mg/dl) |
|---|---|---|---|---|
| NS | 23.2 ± 1.2 | 38.5 ± 1.5 | 29.2 ± 3.2 | 0.37 ± 0.07 |
| DADS (50 mg/kg) | 23.6 ± 1.6 | 38.8 ± 2.8 | 28.6 ± 2.6 | 0.39 ± 0.06 |
| DADS (100 mg/kg) | 23.9± 1.7 | 39.5 ± 2.5 | 30.0 ± 2.0 | 0.43 ± 0.04 |
| DADS (200 mg/kg) | 24.1 ± 1.9 | 39.1 ± 3.1 | 28.9 ± 2.1 | 0.42 ± 0.03 |
Table 1: Biochemical analysis of serum after treatment of MGC 803 cell xenograft mice with DADS. Mice (n = 4 per group) were treated with DADS (50, 100, and 200 mg/kg) or normal saline (NS), respectively. The serum samples were collected for the analysis of biochemistry parameters after the experiment termination. The results are expressed as mean ± SD.
DADS induces G2/M phase cell cycle arrest in xenograft models of human gastric cancer
DADS-induced cell cycle arrest has been demonstrated in association with proliferation inhibition in other tumor cells [27,28], and we therefore proposed that the observed growth inhibition of gastric tumors in xenograft models may also partly result from cell cycle arrest induced by DADS. We thus treated nude mice injected with MGC803 cells as described in Figure 4. MGC803 tumor tissue specimens were collected from the xenografted mice and gastric tumor cells were prepared as described in Materials and Methods. The cell cycle distribution of the gastric tumor cells was monitored by flow cytometry analysis. According to Figure 6, DADS treatment arrested gastric tumor cells in the G2 phase of the cell cycle, as seen in positive sodium butyrate-treated controls. Induction of G2 phase arrest was dose-dependent, which was consistent with the suppression of tumor growth produced by DADS. The percentage of G2 phase cells was 13.1% in the saline control group. In contrast, the percentages of G2 phase cells in the DADS-treated groups (50, 100, and 200 mg/kg) were 14.9%, 29.1%, and 44.1%, respectively. This suggests that high doses of DADS exerted a more potent effect than low doses, causing accumulation of gastric tumor cells in the G2/M phase of the cell cycle. Administration of DADS thus significantly retarded gastric tumor growth in xenografted models in association with DADS-mediated G2/M phase cell cycle arrest and suppression of cancer cell proliferation.
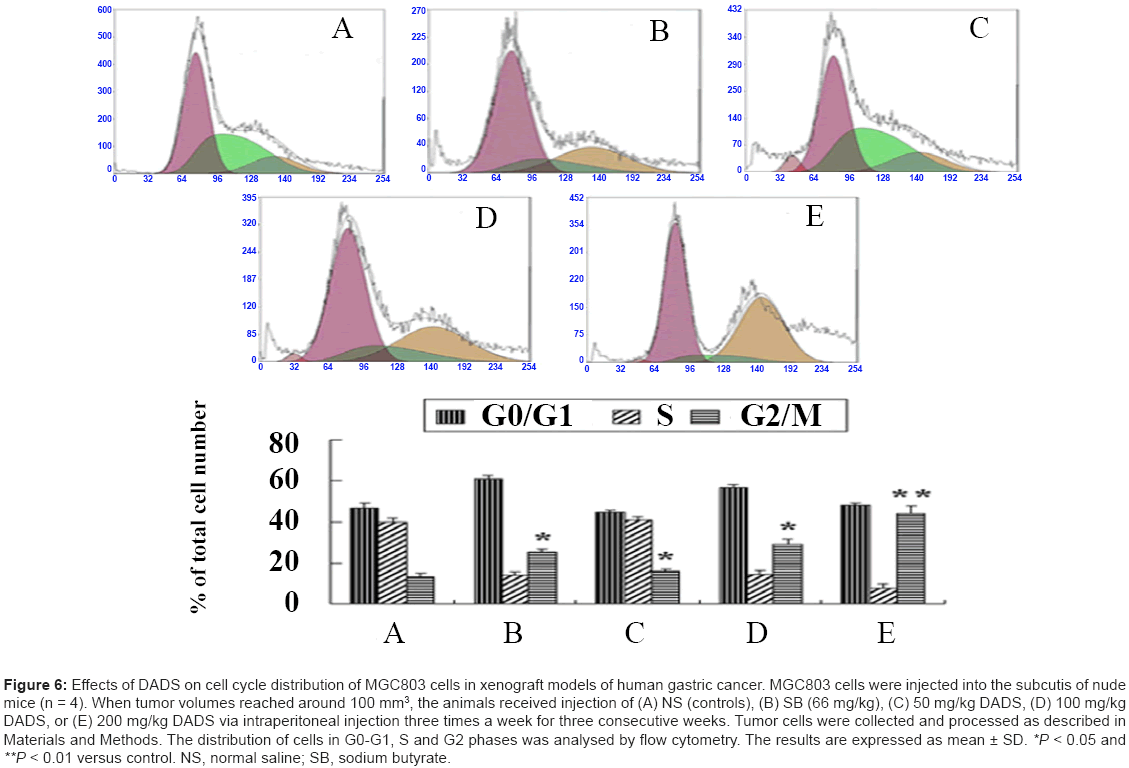
Figure 6: Effects of DADS on cell cycle distribution of MGC803 cells in xenograft models of human gastric cancer. MGC803 cells were injected into the subcutis of nude mice (n = 4). When tumor volumes reached around 100 mm3, the animals received injection of (A) NS (controls), (B) SB (66 mg/kg), (C) 50 mg/kg DADS, (D) 100 mg/kg DADS, or (E) 200 mg/kg DADS via intraperitoneal injection three times a week for three consecutive weeks. Tumor cells were collected and processed as described in Materials and Methods. The distribution of cells in G0-G1, S and G2 phases was analysed by flow cytometry. The results are expressed as mean ± SD. *P < 0.05 and **P < 0.01 versus control. NS, normal saline; SB, sodium butyrate.
DADS increases the acetylation levels of histones H3 and H4 in xenografted models of human gastric cancer
Treatment with DADS has been reported to increase acetylation of histones H3 and H4 in normal rat colonocytes [29]. In the present In vitro study, levels of acetylated histones H3 and H4 were elevated after incubation of MGC803 cells with DADS. To identify the ability of DADS to induce hyperacetylation of histones H3 and H4 In vivo, we performed western blot analysis using total protein lysates extracted from MGC803 gastric cancer tissues from xenografted mice. Compared with the positive (sodium butyrate) controls, hyperacetylated histones H3 and H4 were detected in the DADS-treated groups with increasing DADS doses. Doses of 100 and 200 mg/kg DADS induced acetylation of histone H3 with significant 1.8- and 1.7-fold increases (Figure 7A and 7B), and acetylation of histone H4 with significant 1.3- and 1.5- fold increases (Figure 7C and 7D). These results indicate that highdose DADS had a more pronounced effect on acetylation of histone H3 compared with histone H4, relative to the effect of treatment with 50 mg/kg DADS. DADS thus caused hyperacetylation of histones H3 and H4 in xenograft models of human gastric cancer, and preferentially induced acetylation of histone H3 compared to histone H4. These results generally support those of the In vitro experiments.
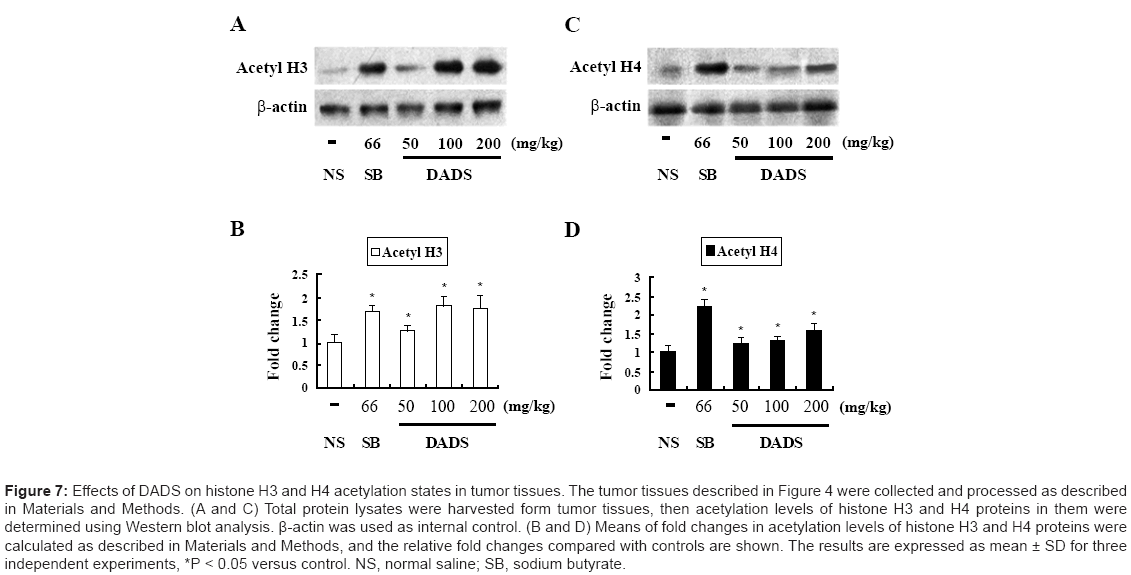
Figure 7: Effects of DADS on histone H3 and H4 acetylation states in tumor tissues. The tumor tissues described in Figure 4 were collected and processed as described in Materials and Methods. (A and C) Total protein lysates were harvested form tumor tissues, then acetylation levels of histone H3 and H4 proteins in them were determined using Western blot analysis. β-actin was used as internal control. (B and D) Means of fold changes in acetylation levels of histone H3 and H4 proteins were calculated as described in Materials and Methods, and the relative fold changes compared with controls are shown. The results are expressed as mean ± SD for three independent experiments, *P < 0.05 versus control. NS, normal saline; SB, sodium butyrate.
DADS increases P21WAF1 protein expression in xenograft models of human gastric cancer
Given that DADS induced hyperacetylation of histones H3 and H4 and cell cycle arrest, we examined its effect on P21WAF1 protein expression in MGC803 gastric tumor tissues from xenografted mice using western blot analysis. Treatment with DADS increased P21WAF1 protein expression in a similar manner to treatment with sodium butyrate (Figure 8A and 8B). Administration of 50, 100, and 200 mg/ kg DADS produced significant 1.2, 1.7 and 1.8 fold increases in P21WAF1 protein levels, respectively, compared to the saline control group. The synchronized effects in terms of increased histone acetylation and P21WAF1 protein levels suggest that upregulation of P21WAF1 expression may be associated with hyperacetylation of histones H3 and H4 induced by DADS in gastric cells in xenografted mice.
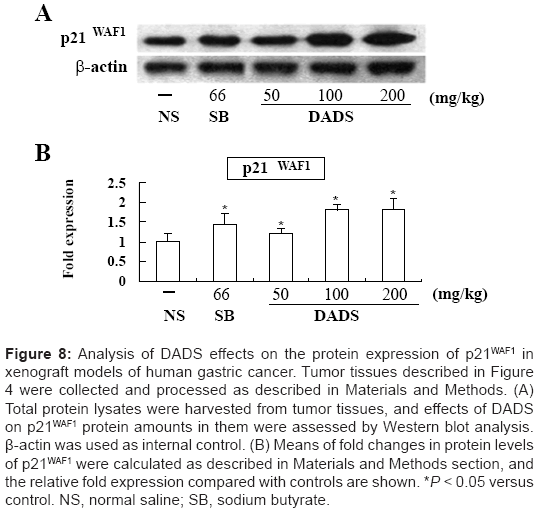
Figure 8: Analysis of DADS effects on the protein expression of p21WAF1 in xenograft models of human gastric cancer. Tumor tissues described in Figure 4 were collected and processed as described in Materials and Methods. (A) Total protein lysates were harvested from tumor tissues, and effects of DADS on p21WAF1 protein amounts in them were assessed by Western blot analysis. β-actin was used as internal control. (B) Means of fold changes in protein levels of p21WAF1 were calculated as described in Materials and Methods section, and the relative fold expression compared with controls are shown. *P < 0.05 versus control. NS, normal saline; SB, sodium butyrate.
Changes in tumor cell morphology and expression of P21WAF1 and proliferating cell nuclear antigen (PCNA) in xenograft models of human gastric cancer
Using tumor tissue sections stained with H&E, we examined the morphological differences in tumor cells between untreated and DADStreated xenografted mice. Representative sections are shown in Figure 9. Light microscopy showed the typical features of poorly differentiated adenocarcinoma in transplantation tumors in the control group. Cancer cells were larger, fusiform, polygonal or elliptical, with a diffuse distribution, arrangement disce, no nest-like structure, high nuclear/ cytoplasmic ratio, karyotin anachromasis, large, obvious nucleoli, and more karyomitosis. After treatment with DADS, cells appeared round or oval, of uniform size, with nest-like arrangements, clear boundaries, abundant cytoplasm, karyotin thinning, small nucleoli, less nuclear division, a reduced karyoplasmic ratio, and decreased heteromorphism. These observations imply that DADS could induce cell differentiation In vivo, in addition to suppressing cell proliferation, both of which might be related to increased P21WAF1 expression.

Figure 9: Analysis of the effects of DADS on cell morphology and the expression of p21WAF1 and PCNA in tumor tissue specimens from xenograft models of human gastric cancer. The left or right panel shows a representative gastric tumor tissue section from MGC803 cell xenograft mice that treated with normal saline as controls or 100 mg/kg DADS, respectively. The slides were H&E stained or stained for p21WAF1 and PCNA (×400 magnification).
We analyzed P21WAF1 expression to confirm the effects of DADS in tumor cells in MGC803 xenografted mice. Treatment with DADS resulted in an apparent increase in P21WAF1 expression that was consistent with the result of western blot analysis (Figure 9). Inhibition of PCNA expression by DADS has been demonstrated in breast cancer cells [25]. PCNA expression was detected in untreated controls, but was decreased in the DADS-treated group (Figure 9).
Epigenetic alterations are known to be as important as genetic alterations in cancer development and progression. Together with DNA hypermethylation, histone hypoacetylation is among the most common epigenetic modifications in cancers, and its implication in dysregulated gene expression has emphasized the importance of retroconversion of histone deacetylation [10]. This has been supported by the successful application of HDAC inhibitors in clinical trials for some cancer therapies [30]. Notably, a previous review discussed the cancer chemopreventive properties of DADS as an HDAC inhibitor, based on experimental studies In vitro and/or In vivo [31], and characterized DADS as a potential anticancer agent with no gross toxicity [25]. DADS has thus attracted increasing interest through its ability to target histone acetylation.
In this study, we provided the first validation that treatment with DADS augmented the acetylation of histones H3 and H4 in gastric cancer MGC803 cells both In vitro and In vivo. In vitro experiments showed that acetylated histone H3 increased after treatment of cells with 30 mg/L DADS for 6 h; this effect was maintained for up to 24 h and then decreased slightly at 24 h. This dynamic was partially consistent with previous results for human colon tumor cells and erythroleukemic cells. The transitory effect of DADS on histone H3 acetylation may be the result of its rapid metabolism [9,23]. In contrast, hyperacetylation of histone H4 required longer treatment with DADS or a higher dose compared to histone H3. DADS and other dietary agents such as butyrate are generally weak ligands of HDAC, and their inhibitory effect on HDAC activity requires higher concentrations than those of non-dietary HDAC inhibitors such as trichostatin A [30,32]. Moreover, DADS-triggered hyperacetylation of histones may take time to allow DADS-targeting of HDAC or the metabolic conversion of DADS to its more effective metabolites [32]. For example, the DADS metabolite allyl mercaptan can exert a more potent inhibitory effect on HDAC than DADS [9]. Moreover, prolonged hyperacetylation of histones H3 and H4 was obtained in MGC803 cell xenografts In vivo by successive administration of DADS to nude mice for 3 weeks. It is possible that persistent DADS-induced hyperacetylation of histone H3 or histone H4 associated with cell growth inhibition may need repetitive treatments In vitro or administrations In vivo to maintain a durable effect. This hypothesis is supported by experiments in human colon tumor cells and bladder cancer cells [24,33]. In general, however, DADS was able to increase histone acetylation, especially of histone H3, in MGC803 cells both In vitro and In vivo. This effect may result from inhibition of HDAC activity [9,23].
Numerous studies have verified that increased acetylation of histones mediated by various HDAC inhibitors could selectively reactivate tumor suppressor genes in diverse cancers [33,34]. Among these, P21WAF1 is one of the most commonly induced genes [35-40]. It is a vitally important cyclin-dependent kinase inhibitor with important roles in regulating cell cycle progression and inducing cell cycle arrest in G1 or G2 phase [13-15]. It has been shown that the P21WAF1 gene promoter was unmethylated in eight gastric tumor cell lines, and HDAC inhibitors (n-butyric acid and trichostatin A) but not a DNA demethylating agent (5-aza-2'-deoxycytidine) were able to activate P21WAF1 gene transcription, compared with other cyclindependent kinase inhibitor genes of the KIP family, such as p27 and p57 [41]. Dysregulation of deacetylation may thus play a major role in epigenetic alterations leading to the silencing of this gene in gastric cancer, while HDAC inhibitors could reactivate the gene by modulating histone acetylation. The association between reduced P21WAF1 expression and histone deacetylation is further supported by studies showing that histones H3 and H4 in the P21WAF1 promoter and coding regions were hypoacetylated in gastric carcinoma specimens, and that hyperacetylation of histones H3 and H4 was correlated with increased P21WAF1 expression in gastric carcinoma cell lines treated with trichostatin A [42].
Mounting evidence has shown that a reversal of abnormal histone deacetylation is involved in DADS-induced growth inhibition of various human tumors cell lines [9,23]. Our previous study demonstrated that DADS-induced G0/G1 phase cell cycle arrest and growth inhibition of human leukemic HL-60 cells In vivo and in vitro were associated with hyperacetylation of histones and increased P21WAF1 expression [43]. The present study found that the growth rate of MGC803 xenografted neoplasms decreased gradually in a dosedependent fashion, accompanied by G2/M phase cell cycle arrest after DADS administration to nude mice, in accord with the In vitro results. Furthermore, both P21WAF1 expression and acetylated histones H3 and H4 increased coincidentally in MGC803 cell xenografts in nude mice, especially after administration of high-dose DADS. These results suggest that DADS-induced G2/M phase cell cycle arrest and growth inhibition of human gastric cancer cells may be associated with increased P21WAF1 expression, resulting in turn from hyperacetylation of histones H3 and H4.
In addition, we observed signs of cell differentiation and decreased PCNA expression in tumor cells from xenografted mice treated with DADS. However, the present and previous studies [18-22,25,44] could not exclude the possibility that the growth inhibition of xenografted gastric cancer cells triggered by DADS may in part result from the induction of cell differentiation and/or suppression of cell proliferation mediated by molecules other than P21WAF1.
It has been proposed that the induction of P21WAF1 expression by HDAC inhibitors can be p53-independent [45-47] or p53-dependent [48]. To the best of our knowledge, allele loss or p53 mutation is one of the most frequent events in gastric cancer [49,50]. Previous data showed that P21WAF1 expression induced by trichostatin A was barely influenced by dominant-negative mutation of p53, whereas histone H4 acetylation was reduced in cells expressing dominant-negative p53 [42]. The current study demonstrated that treatment with DADS had a more potent effect on hyperacetylation of histone H3 than of histone H4. We therefore hypothesize that hyperacetylation of histone H3 may play a more vital role in DADS-reactivation of P21WAF1 expression than histone H4, and that histone H3 might be a preferential target for HDACs, the activity of which could be restrained by DADS in gastric tumor cells. However, further studies are needed to confirm this hypothesis. A recent study indicated that the acetylated histone H4 N-terminal tail peptide potentiated p53-mediated transactivation of target genes through interaction with G9a histone methyltransferase and HDAC1, inhibiting tumor cell growth and enhancing apoptosis [51]. We thus considered the possibilities that: (a) acetylated histone H4 induced by DADS promotes histone H3 acetylation by interaction with and inhibition of HDAC1, facilitating P21WAF1 gene transcription; and (b) DADS-induced cell cycle arrest and growth inhibition may involve acetylated histone H4-mediated expression of other tumor suppressor genes, including P21WAF1 through other molecular mechanisms. An association between DADS-induced P21WAF1 expression and hyperacetylation of histones H3 and H4 could explain the induction of G2/M phase cell cycle arrest in human gastric cancer MGC803 cells In vivo and In vitro. Further experiments using chromatin immunoprecipitation assays are needed to confirm that DADS-induced hyperacetylation of histones H3 and H4 could enhance transcriptional activity of the P21WAF1 gene promoter region.
In addition, a previous study reported that DADS inhibited the growth of human colon cancer cell xenografts in mice, without affecting body weight, liver, kidney and spleen weights, or BUN level [52]. Furthermore, no differences in body weights between DADS-treated and untreated human breast cancer xenografted mice were observed [25]. These observations indicate that DADS has no gross toxicity. On the basis of serum biochemical analysis and histopathological examination of major organ tissues, the results of the current study support a lack of gross toxicity for DADS.
In summary, this study demonstrated that DADS, as a putative HDAC inhibitor, can induce G2/M phase cell cycle arrest and growth inhibition of human gastric cancer cells In vivo and In vitro. The mechanism underlying these effects of DADS may relate to increased P21WAF1 expression, which could in turn result from hyperacetylation of histones. In terms of epigenetics, the modulation of deregulation of histone acetylation may represent one of the mechanisms responsible for the anti-gastric cancer activity of DADS.
This work was supported by The National Natural Scientific Foundation of China (No. 30600285, No. 31000629, No. 31100935, No. 81102854), The Key Project Foundation of the Science and Technology program of Hunan Province of China (No. 04SK1004), The Key Project of Scientific Research Foundation of Hunan Province Education Department of China (No. 04A047, No. 09A077), The Patency Foundation of Innovation Platform of Hunan Provincial University of China (No. 09K074) and The Key Hunan Provincial Natural Science Foundation of China (No. 07JJ3033).