Drug Designing: Open Access
Open Access
ISSN: 2169-0138
ISSN: 2169-0138
Research Article - (2014) Volume 3, Issue 3
In spite of the many studies carried out on glioblastoma multiforme (GBM) in recent years and the implementation of promising new therapeutic strategies, median survival rates in GBM patients have not been improved. A key issue in the fight against a tumor is learning its biology which is only possible in laboratory conditions that accurately reproduce its physiological microenvironment. GBM is among the most hypoxic and glycolytic tumors of the central nervous system. The most recent data suggest that these two metabolic features should be recognized as the most important markers of the GBM aggressive phenotype associated with its resistance to chemo- and radiotherapy. However, until now, the effect of modulation of the tumor’s microenvironment on drug efficacy has been evaluated only in a few studies.
Since hypoxia inside a tumor creates protection for GBM cells from chemotherapy and the spatial arrangement of hypoxia in GBM is a dynamic process (the level of pO2 inside a tumor is from 0% to 5%), our experiments performed on the T98G glioblastoma cell line were carried out in a range of oxygen availability conditions and then compared to standard laboratory conditions. Moreover, the influence of high or standard glucose concentration or its deprivation in the culture medium on temozolomide effect (the standard chemotherapeutic for GBM) was estimated. We examined the influence of temozolomide on cell viability, division and apoptosis as well as on migration of tumor cells by using the time-lapse real-time observations.
Our study showed that glioblastoma cells cultured in conditions of chronic hypoxia were almost completely resistant to the effect of the drug. Moreover, we observed that increased glucose availability in the GBM microenvironment was associated with the enhanced resistance of the GBM to the cytotoxic effect of temozolomide in hypoxic conditions which was manifested by an increased cellular viability and division and a diminished percentage of apoptotic cells.
<Keywords: Glioblastoma multiforme, Chemo/radioresistance of glioblastoma, Hypoxia in brain tumors, High-glycolytic tumors
Glioblastoma multiforme (GBM) is the most aggressive primary tumor in humans and still a fatal form of the central nervous system cancer [1,2]. Since all GBM treatment methods used so far, have failed, including the most promising gene therapy, there is a need for a search for other, as yet unknown targets for future therapeutic strategies. The difficulties in glioblastoma treatment stem from the acquisition of radio- and chemoresistance by its cells, by mechanisms which remain undiscovered [3]. It is currently suggested that crucial factors in glioblastoma treatment resistance comprise weak drug penetration into the tumor cells due to hypoxia inside the tumor and its strongly glycolytic character [4]. It would, therefore, seem that pharmacological modulation of the tumor’s metabolism and microenvironment could improve therapeutic efficacy [5].
The tumor microenvironment is entirely different from the microenvironment in healthy tissues, as has been demonstrated by studies with the use of Eppendorf mini-electrodes [6]. It has been proven that partial oxygen concentration inside the tumor approximates 0-5% while normal oxygen concentration in the CNS ranges from 10% to 13% [7]. Such hypoxic niche conditions induce multidirectional changes in the proteome and genome, leading to: promotion of an aggressive phenotype of tumor cells facilitating their growth and spread, distortion of the cell cycle, enhanced angiogenesis and differentiation of tumor progenitor cells, which finally results in treatment resistance [8].
Although the consequences of hypoxia are widely known, the significance of this factor in clinical practice is often questioned and marginalized. However, in the light of the above-mentioned effects of hypoxia, studies on GBM should be conducted under conditions characteristic for the tumor and not under so-called standard conditions, i.e. in an atmosphere containing ca. 20% oxygen, which are hyperoxic conditions for glioblastoma cells.
Apparently, in spite of the heterogeneity and genetic instability of GBM cells, hypoxia and increased glucose turnover are common to all types of these tumors. Hypoxia inside the tumor is a dynamic process which means that as the tumor grows, cells arranged in layers and surrounding blood vessels can be oxygenated while others located further than 15 μm away from a blood vessel can suffer acute or transient hypoxia, while the palisade cells of the tumor migrate from hypoxic to more oxygenated regions [9-11]. It seems that cells that adapt to hypoxia have the greatest clinical significance of all the cells constituting the tumor biomass. These cells are characterized by permanent genetic changes and unique cellular energy metabolism, which is distinguished by a lack of vulnerability to apoptosis and enhanced glucose metabolism described as the Warburg effect (for the discovery of which Otto Warburg was awarded the Nobel Prize in 1931). To date few studies have demonstrated that GBM cells utilize only glucose as an energy source but this fact indicates that they lack metabolic flexibility [12-14].
The present studies were conducted on the human glioblastoma cell line T98G in the 4th, 7th and 10th passages. We evaluated the efficacy of temozolomide, the first-choice chemotherapeutic in GBM patients. This is an alkylating and anti-angiogenic drug that increases sensitivity to radiotherapy. Although this is not a causal treatment, it improves the quality of life of a large percentage of patients. The cultures were maintained in a range of atmospheres containing different oxygen concentrations (1-5%, 10% and 20%) and in a medium supplemented with different glucose contents (0; 1 and 4.5 g/L). The experiments were designed to reproduce acute, gradual and chronic hypoxia and hypoxia/reoxygenation and to check: 1) how changes in glucose concentration affect temozolomide efficacy and 2) what micro environmental conditions of the glioblastoma induce the greatest resistance to this drug. Cell viability, apoptosis and cell division were evaluated in cultures exposed to or deprived of cytotoxic temozolomide concentrations.
Cell cultures
T98G cell line was originally established from a glioblastoma multiforme tumor in a 61-year-old Caucasian male and purchased from Sigma-Aldrich (St Louis, MO, USA). Medium for cell culture, gentamicin and fetal bovine serum were purchased from Gibco-BRL. Plasticware for cell cultures was from Falcon (Lexington, TN), Nunc (US) and Eppendorf (Germany). Temozolomide was obtained from Sigma Aldrich.
Study design
Glioblastoma cell line T98G cells from an original vial were thawed and a part of them was supplemented with an equal amount of a cryoprotectant and placed in a liquid nitrogen Dewar flask. The remaining cells were immediately used for experiments. Cultures were established and maintained in sterile conditions in a laminar flow cabinet (New Air, USA) and in incubators for standard laboratory conditions in an atmosphere containing 20% O2 (New Air) and for ischemic conditions in 1–10% O2 (Galaxy 48R, New Brunswick, USA). Experiments were conducted in the 4th, 7th and 10th passages on the second day after trypsinization. The following determinations were made prior to the experiments:
1. Cell density was evaluated by cell counters: Eve (Life Technologies, Grand Islands NY), Scepter (Millipore, Germany), flow cytometer Muse (Millipore, Germany), and spectrophotometer Tecan (TK Biotech, Switzerland). The experiment was conducted on cultures with cell density of 1.9×105 cells
2. Cytotoxic concentration of temozolomide was determined with the use of the MTT test, annexin V assay, the 7-AAD staining protocol, using a flow cytometer (Muse) and cell counter (Eve) equipped with a module for determination of cell viability. The studies showed the toxic effect of temozolomide at 1000 μM which is presented in Figure 1.
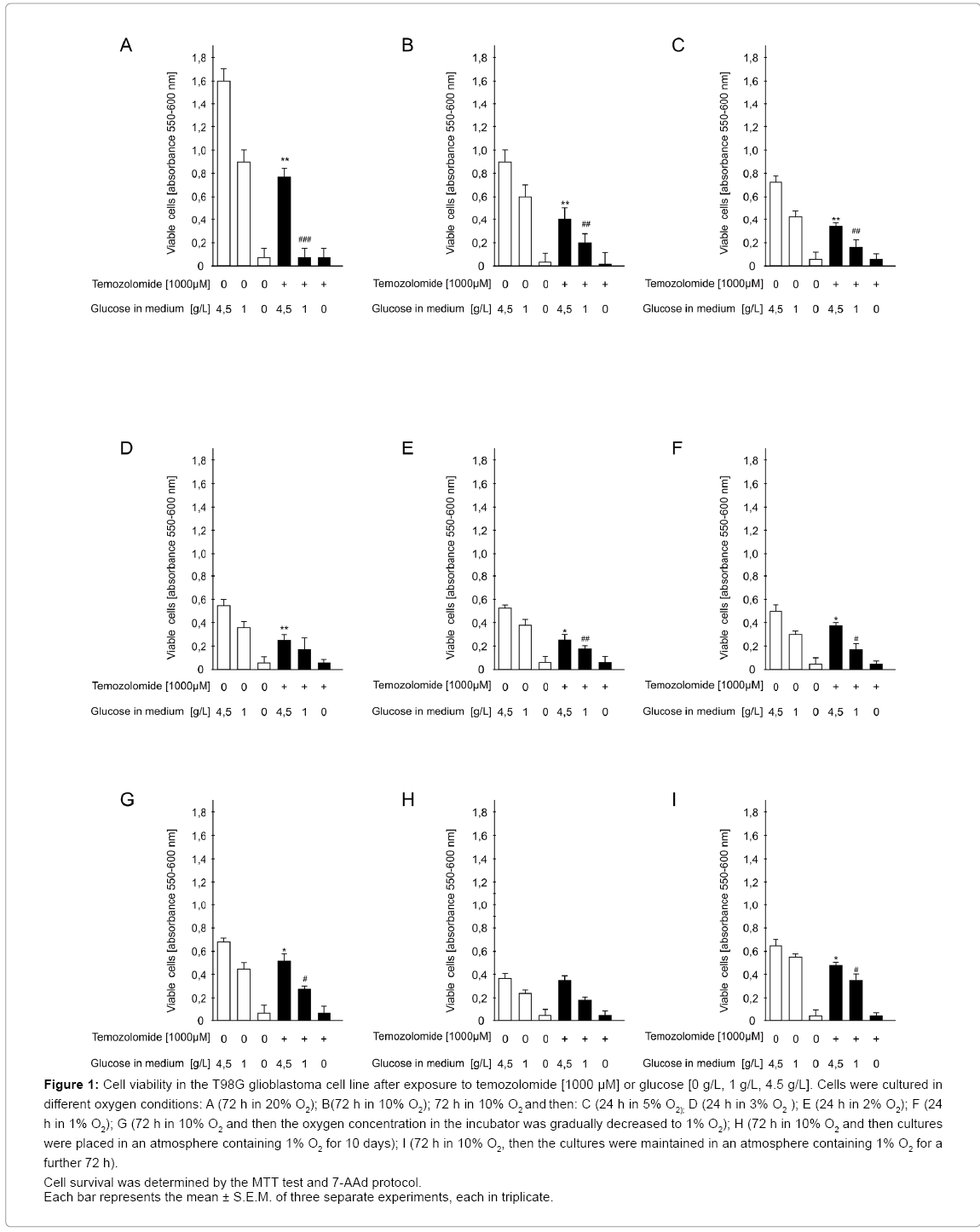
Figure 1: Cell viability in the T98G glioblastoma cell line after exposure to temozolomide [1000 μM] or glucose [0 g/L, 1 g/L, 4.5 g/L]. Cells were cultured in different oxygen conditions: A (72 h in 20% O2); B(72 h in 10% O2); 72 h in 10% O2 and then: C (24 h in 5% O2); D (24 h in 3% O2 ); E (24 h in 2% O2); F (24 h in 1% O2); G (72 h in 10% O2 and then the oxygen concentration in the incubator was gradually decreased to 1% O2); H (72 h in 10% O2 and then cultures were placed in an atmosphere containing 1% O2 for 10 days); I (72 h in 10% O2, then the cultures were maintained in an atmosphere containing 1% O2 for a further 72 h).
Cell survival was determined by the MTT test and 7-AAd protocol.
Each bar represents the mean ± S.E.M. of three separate experiments, each in triplicate.
Glioblastoma cell line T98G cultures were maintained in culture flasks (25 cm3 lub 65 cm3) in DMEM supplemented with 10% FBS, 1% gentamicin, 1g/L D-glucose, L-glutamine, 25 mM Hepes, pyruvate. The medium was replenished every third day. When cells reached 90% confluence, cultures were trypsinized and passaged to flat-bottomed 96-well plates (to determine biochemical parameters) and to 35-mm Petri dishes (for microscopic examination). The studies were carried out in several experimental models differing in the micro environmental conditions in which temozolomide was tested for the effect on glioblastoma cell viability, apoptosis and cell division. The results for each experiment came from 3 independent repetitions. In parallel, cell cultures were examined using real-time time-lapse microscopic recording. Conditions of each experiment are described below.
Experiment I: standard laboratory conditions: Cells were cultured in an atmosphere containing 20% O2. On the second day after trypsinization, the medium was replaced with a fresh portion of the same composition containing glucose at 4.5 g/l or 1 g/l or without glucose. In these conditions, cells were exposed or not to temozolomide at a concentration of 1000 μM. Cultures were maintained for 72 h.
Experiment II: conditions close to physiological: Cells were incubated in an atmosphere containing 10% O2. On the second day after trypsynization, the medium was replaced as described in Experiment I. Cultures were maintained for 72 h.
Experiment III: acute and moderate ischemia: Cells were incubated in an atmosphere containing 10% O2. On the second day after trypsynization, culture medium was replaced by a specific medium as described in Experiment I. Then, the plates were exposed to ischemic conditions. i.e. to an atmosphere containing 1, 2, 3, 4 or 5% O2 for 24 h. Culture in an atmosphere containing 1% O2 simulated “acute ischemia”, while the remaining oxygen concentrations mirrored a moderate ischemia.
Experiment IV: gradually progressing ischemia: Cells were cultured in an atmosphere containing 10% O2. On the second day after trypsinization, after the medium had been replaced as in Experiment I, oxygen concentration was gradually decreased in an incubator until 1% O2 was reached. Cells were cultured under such conditions for 72 h. This experimental model aimed to reconstruct situation “in vivo” when ischemia develops gradually within the tumor and tumor cells gradually adapt to it.
Experiment V: chronic ischemia: Cells were cultured in an atmosphere containing 10% O2. After medium replacement as in Experiment I, culture plates were placed in an incubator in an atmosphere containing 1% O2 Cultures were maintained in such conditions for 10 days.
Experiment VI: hypoxia/reoxygenation: Cells were cultured in an atmosphere containing 10% O2. After medium replacement as in Experiment I, cultures were maintained in an atmosphere containing 1% O2 for 72 h and 5% O2 for the next 72 h. The aim of this experimental model was to simulate hypoxia/reoxygenation to which a pool of tumor cells is exposed in vivo.
Cell viability
Two methods were used for evaluation of cell viability: MTT assay and 7-AAD staining protocol.
In the MTT assay, the ability of cells to convert MTT (Sigma- Aldrich) into formazan is a measure of mitochondrial activity and in consequence, cell viability Mosman et al. [15]. MTT (0.25 mg/ml) was added to the medium for 3 h (37°C) before the end of the experiment. After washing with phosphate buffered saline (PBS), the cells were lysed in 100 μl of dimethyl sulfoxide (Sigma–Aldrich) to release the blue reaction product formazan. Absorbance at the wavelength of 570 nm was read on a microplate reader (Multiscan, Labsystems, Helsinki, Finland).
7-AAD staining protocol
This method is based on evaluation of functional disturbances of the cell membrane, which is evaluated using a flow cytometer and a cell viability assay kit. When integrity of the cell membrane of viable cells is preserved, 7-AAD does not stain intracellular components because it does not penetrate into the cell. The results were read after a 5-minute incubation of cells with the reagent at room temperature, according to the manufacturer’s recommendations.
Apoptosis
Apoptosis was assessed using a flow cytometer (Muse) and annexin V apoptosis detection kit. This protein shows a strong affinity for phosphatidylserine, which is translocated to the surface of the cell membrane in an early stage of apoptosis. After trypsynization and neutralization, cells were exposed to temozolomide, incubated for 20 min in the dark with Annexin V Dead Cell Reagent (100 μl), and then the protocol recommended by the manufacturer was followed.
Determination of mitotic potential
The intensity of cell division was evaluated using an immunoenzymatic assay kit based on determination of bromodeoxyuridine (Brdu) level (Abcam, Cambridge, UK), which is incorporated into DNA of dividing cells. The determination was conducted according to the manufacturer’s instructions. Absorbance was measured in a Multiscan RC microplate reader (Labsystems, Helsinki, Finland).
Time-lapse recording
Cell were cultured on Petri dishes in standard laboratory conditions in media differing in glucose and temozolomide concentrations, after cells reached 90% confluence. Cell cultures were observed for 72 h using a fluorescence cell analyzer JuLi (NanoEntec, South Korea). A real-time image was captured every 20 min and time-lapse movie was compiled from the whole observation period.
Statistical analyses
Statistical analysis of the data was performed using a two-way ANOVA followed by the post-hoc Tukey test. Differences were considered as significant when p<0.05. Statistical analysis was performed using GraphPad Prism 4.01 software (GraphPad Software, San Diego, CA, USA).
Experiment I
Evaluation of viability, apoptosis and intensity of cell division in glioblastoma cells exposed to temozolomide in the medium with or without glucose under standard conditions: Temozolomide reduced glioblastoma cell viability by 65% and 85% in the medium with glucose at 4.5 g/l and 1 g/l, respectively, vs. drug-untreated cells (p<0.01 or p<0.001, respectively). A significant decrease in cell viability was also seen in cultures maintained in the glucose-free medium, both exposed and not exposed to the drug, by 89% (p<0.001) and 91% (p<0.001), respectively, compared to cultures on the glucose 4.5 g/L-supplemented medium (Figure 1A).
Absolute absorbance values in the MTT viability test in cultures cultivated under standard conditions were much higher than in cultures maintained in an atmosphere containing 10% O2.
The percentage of apoptotic cells was inversely correlated with the percentage of viable cells (Figure 2A).
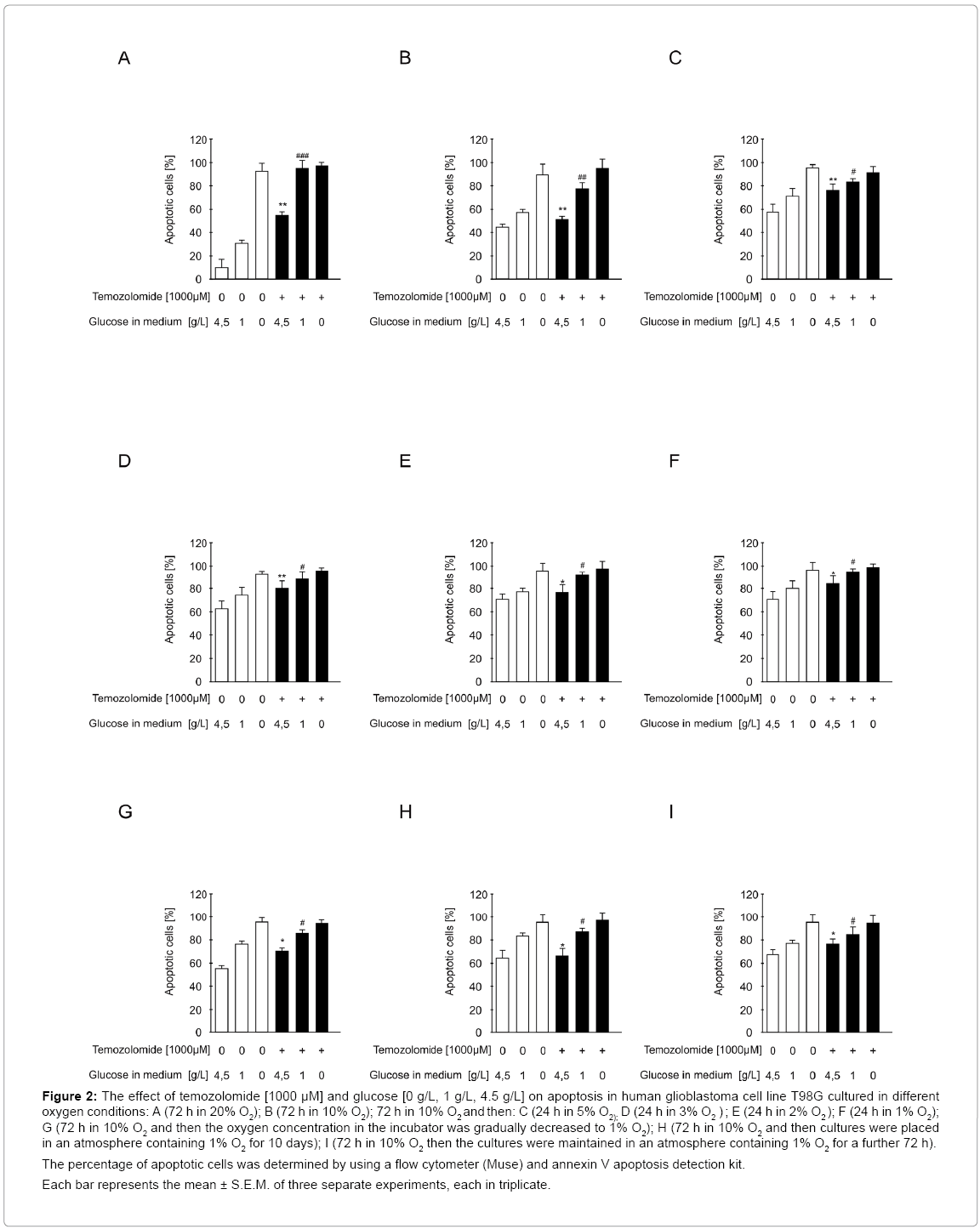
Figure 2: The effect of temozolomide [1000 μM] and glucose [0 g/L, 1 g/L, 4.5 g/L] on apoptosis in human glioblastoma cell line T98G cultured in different oxygen conditions: A (72 h in 20% O2); B (72 h in 10% O2); 72 h in 10% O2 and then: C (24 h in 5% O2); D (24 h in 3% O2 ) ; E (24 h in 2% O2 ); F (24 h in 1% O2); G (72 h in 10% O2 and then the oxygen concentration in the incubator was gradually decreased to 1% O2); H (72 h in 10% O2 and then cultures were placed in an atmosphere containing 1% O2 for 10 days); I (72 h in 10% O2 then the cultures were maintained in an atmosphere containing 1% O2 for a further 72 h).
The percentage of apoptotic cells was determined by using a flow cytometer (Muse) and annexin V apoptosis detection kit.
Each bar represents the mean ± S.E.M. of three separate experiments, each in triplicate.
Cell division under standard conditions was intense (like viability) and depended on glucose concentration in the medium. Temozolomide reduced the intensity of glioblastoma cell division by 61% (medium with glucose 4.5 g/L) (p<0.01) and 76% (medium with glucose 1g /L) (p<0.01) vs. drug-free cultures. The intensity of cell division in cultures exposed to temozolomide and those that were drug-free maintained on a glucose-free medium was similar (Figure 3A), and was higher compared to the cells cultured in an atmosphere containing 10% O2.
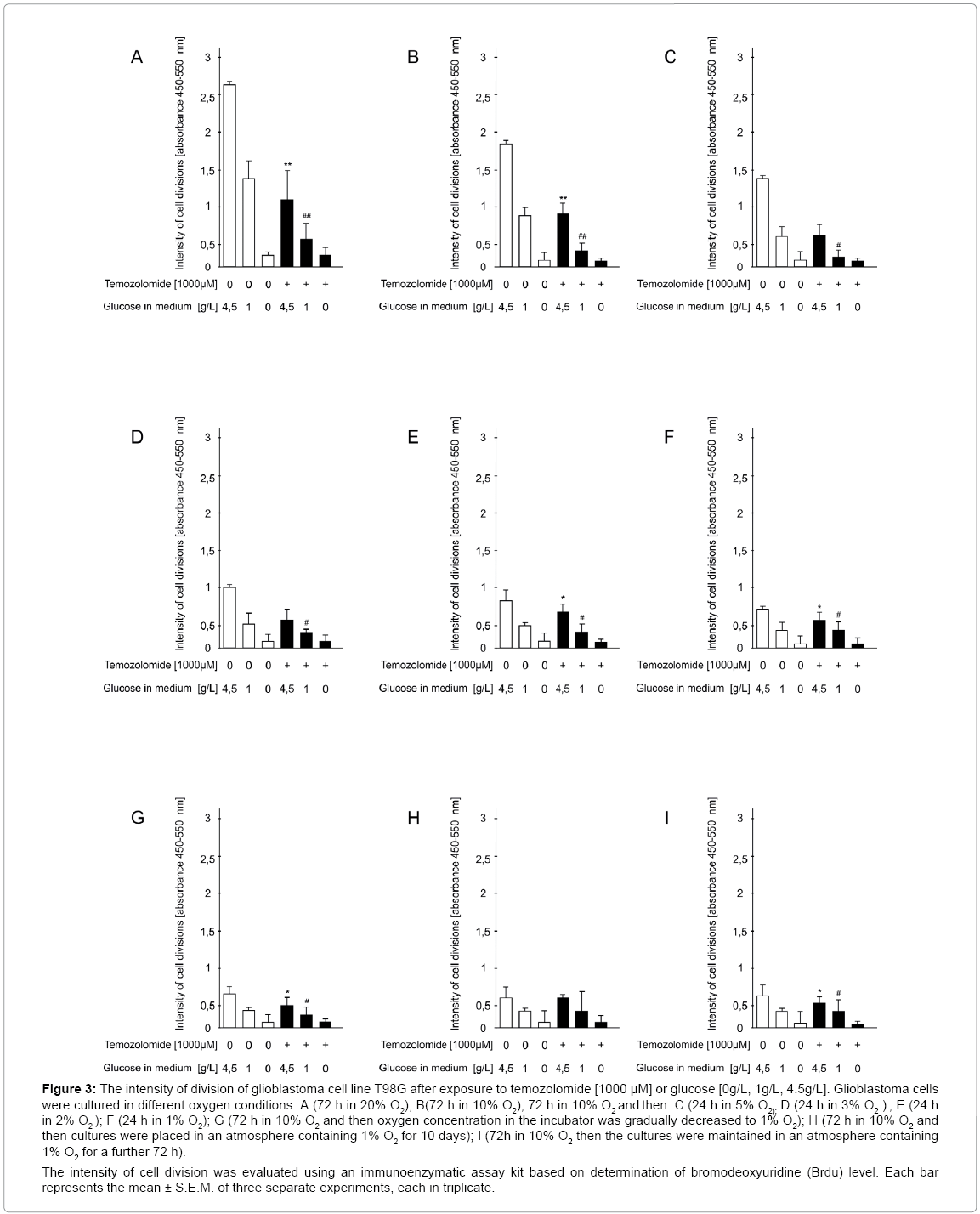
Figure 3: The intensity of division of glioblastoma cell line T98G after exposure to temozolomide [1000 μM] or glucose [0g/L, 1g/L, 4.5g/L]. Glioblastoma cells were cultured in different oxygen conditions: A (72 h in 20% O2); B(72 h in 10% O2); 72 h in 10% O2 and then: C (24 h in 5% O2); D (24 h in 3% O2 ) ; E (24 h in 2% O2 ); F (24 h in 1% O2); G (72 h in 10% O2 and then oxygen concentration in the incubator was gradually decreased to 1% O2); H (72 h in 10% O2 and then cultures were placed in an atmosphere containing 1% O2 for 10 days); I (72h in 10% O2 then the cultures were maintained in an atmosphere containing 1% O2 for a further 72 h).
The intensity of cell division was evaluated using an immunoenzymatic assay kit based on determination of bromodeoxyuridine (Brdu) level. Each bar represents the mean ± S.E.M. of three separate experiments, each in triplicate.
Experiment II
Evaluation of viability, apoptosis and intensity of cell division in glioblastoma cells exposed to temozolomide in the medium with or without glucose under conditions close to physiological: Temozolomide reduced glioblastoma cell viability by 55% (medium with glucose 4.5g/L) (p<0.01) and by 66% (medium with glucose 1g/L) (p<0.01) compared to the drug-free group. Only elimination of glucose from the medium caused a marked decrease in cell viability that was comparable in both drug-exposed and unexposed cultures (by 87% (p<0.001) and 90% (p<0.001), respectively) compared with drug-free culture on the medium with glucose 4.5 g/L (Figure 1B).
The percentage of apoptotic cells was inversely correlated with the percentage of viable cells and comparable with the percentage of dead cells (Figure 2B).
In this experimental model, the intensity of cell division also depended on glucose concentration in the medium. Temozolomide reduced the intensity of glioblastoma cell division by 50% (medium with glucose 4.5 g/L) (p<0.01) or 63% (medium with glucose 1g/L) (p<0.01) vs. drug-unexposed cultures. However, on the glucose-free medium, there were no differences in cell division intensity between temozolomide-exposed and unexposed cultures (Figure 3B).
Experiment III
Evaluation of viability, apoptosis and intensity of cell division in glioblastoma cells exposed to temozolomide in the medium with or without glucose under conditions of acute and moderate ischemia
Moderate ischemia: The cytotoxic effect of temozolomide in cells cultured in an atmosphere containing 5%, 4% (data not shown) or 3% O2 was similar. Temozolomide reduced glioblastoma cell viability by 47% (5% O2) (p<0.01) or 44% (3% O2) (p<0.01) (medium with glucose 4.5 g/L) vs. drug-free control cultures. When the culture medium was supplemented with a lower glucose concentration (1 g/L), the respective values amounted to: 56% (5% O2) (p<0.01) or 50% (3% O2) (p<0.01). The decrease in cell viability in cultures cultivated on the glucosefree medium was significant both in drug-exposed cultures: by 86% or 89% (5% or 3% O2) (p<0.001) (medium with glucose 4.5 g/L), and drug-unexposed cultures: by 90% or 87% (5% or 3% O2, respectively) (p<0.001) vs. drug-free control cultures (medium with glucose 4.5 g/L) (Figure 1C and 1D).
The percentage of apoptotic cells was inversely correlated with the percentage of viable cells and comparable with the percentage of dead cells (Figure 2C and 2D).
Temozolomide reduced cell division intensity by 43% (5% O2) (p<0.01) or 33% (3% O2) (p<0.01) (medium with glucose 4.5 g/L). When the medium was supplemented with a lower glucose content (1 g/L), the cell division intensity was decreased by temozolomide by 52% (5% O2) (p<0.01) and 37% (3% O2) (p<0.01) vs. drug-free cultures. On the glucose-free medium, cell division was blocked to a similar extent in the presence and absence of temozolomide (Figure 3C and 3D).
Acute ischemia: Temozolomide decreased glioblastoma cell viability by 47% (2% O2) (p<0.01) and 30% (1% O2) (p<0.01) (medium with glucose 4.5 g/L) vs. drug-free control. A similar effect was observed on the medium supplemented with a lower glucose content (1 g/L), namely, the drug reduced cell viability by 43% (2% O2) (p<0.01) and 37% (1% O2) (p<0.01). Elimination of glucose from the medium caused a considerable decrease in cell viability both in cultures incubated with and without the drug by 90% and 87% (2% O2), (p<0.001) and 83% and 85% (1% O2) (p<0.001) respectively, vs. drug-free control cultured on the medium with glucose 4.5 g/L (Figure 1E, 1F).
The percentage of apoptotic cells was inversely proportional to the percentage of viable cells and comparable with the percentage of dead cells (Figure 2E and 2F).
Temozolomide reduced the intensity of glioblastoma cell division by 23% (2% O2) (p<0.01) and 16% (1% O2) (p<0.01) (medium with glucose 4.5 g/L) or by 26% (2% O2) and 16% (1% O2) (p<0.01) (medium with glucose 1g/L). When cells were cultured on the glucosefree medium, no differences in cell division intensity were observed between temozolomide exposed and unexposed cell cultures (Figure 3E, 3F).
Experiment IV
Evaluation of viability, apoptosis and intensity of cell divisions in glioblastoma cells exposed to temozolomide in the medium with or without glucose under conditions of gradually progressing ischemia
Temozolomide reduced glioblastoma cell viability in cultures on the medium supplemented with glucose 4.5 g/L by 33% (p<0.01) and with glucose 1 g/L by 37% (p<0.01) compared with drug-free control cells. When cells were cultured on the glucose-free medium, no significant differences in cell viability were noted between temozolomide exposed and unexposed cultures (Figure1G).
The percentage of apoptotic cells was inversely proportional to the percentage of viable cells (Figure 2G).
Temozolomide decreased the intensity of cell division on the medium supplemented with glucose 4.5 g/L and 1 g/L by 29% and 37% (p<0.01) respectively, vs. appropriate drug-unexposed control cultures. On the glucose-free medium, cell division intensity was similar independently of the presence or absence of temozolomide (Figure 3G).
Experiment V
Evaluation of viability, apoptosis and intensity of cell divisions in glioblastoma cells exposed to temozolomide in the medium with or without glucose under conditions of chronic ischemia: Suppressive effect of temozolomide on cell viability was very weak, amounting to 5% and 13% (p<0.01) in cultures on the medium containing glucose 4.5 g/L and 1 g/L, respectively, vs. drug-unexposed cultures. On the medium without glucose, the decrease in cell viability was similar irrespectively of the presence of the drug amounting to ca. 92% (p<0.01), vs. drugunexposed cultures on the medium with glucose 4.5 g/L (Figure 1H).
The percentage of apoptotic cells was inversely proportional to the percentage of viable cells (Figure 2H).
The inhibition of cell division in cultures maintained on the glucose-free medium was similar both when the cells were exposed to temozolomide or not (Figure 3H).
Experiment VI
Evaluation of viability, apoptosis and intensity of cell division in glioblastoma cells exposed to temozolomide in the medium with or without glucose under conditions of hypoxia/reoxygenation: Temozolomide reduced glioblastoma cell viability incubated on the medium containing glucose 4.5 g/L and 1g/L by 32% and 37% (p<0.01) respectively, vs. control cultures. Elimination of glucose from the medium markedly decreased cell viability to a comparable extent in drug-exposed and unexposed cultures by 88% and 90% (p<0.001), respectively, vs. drug-free cultures on the medium with glucose 4.5 g/L (Figure 1I).
The percentage of apoptotic cells was inversely proportional to that of viable cells (Figure 2I).
In this experimental model, the suppressive effect of temozolomide on the intensity of glioblastoma cell division was dependent on glucose concentration in the medium, namely, it decreased by 20% (p<0.01) at a glucose concentration of 4.5 g/L and to 24% (p<0.01) at 1 g/L vs. respective drug-unexposed cultures. On the glucose-free medium, there were no differences in cell division intensity between drugexposed and unexposed cultures (Figure 3I).
Experiment VII
Time-lapse imaging of glioblastoma cell migration in cultures exposed to temozolomide in the medium with or without glucose under standard laboratory conditions:
Real-time recording of glioblastoma cells demonstrated that:
A) Cells exposed to temozolomide 1000 μM cultured on the glucose-free medium began to die already after 1 h (Record 1).
B) Cells exposed to temozolomide cultured on the medium containing glucose at 1 g/L began to die after 35 h (Record 2).
C) Cells unexposed to temozolomide cultured on glucose-free medium began to die after 5 h. The analysis demonstrated the shrinkage of glioblastoma cell soma and processes, and the loss of cell adhesion to the culture plate bottom (Record 3).
D) Cells unexposed to temozolomide cultured on the medium containing glucose at 1 g/L migrated and divided intensively (Record 4).
In spite of the availability of temozolomide therapy, which is considered to be the best pharmacotherapy for GBM, improvement of preoperative diagnostic imaging methods, and progress of neurosurgery techniques, only about 3% of patients suffering from this disease survive 36 months [16,17].
These data demonstrate unequivocally that the therapeutic approach used to date should be modified. Studies aiming to produce a better understanding of the biology of tumor cells offer some hope. It is believed that the aggressive character of glioblastoma cells infiltrating brain parenchyma is connected with a high preference of glioblastoma cells for glucose uptake and fluctuations in oxygen concentration inside the tumor mass. These features are decisive for “metabolic remodeling”, a hallmark of this tumor which distinguishes tumor cells from healthy ones [16,18,19].
In this study, for the first time, a number of experiments were performed in one laboratory on one cell line (T98G) and on five models of hypoxia with the aim of mimicking most accurately the fluctuation of hypoxia levels inside the tumor. Previously published data indicated that the glioblastoma mass was built of several populations of cells located in its different regions: in the oxygenated region and under chronic, moderate and acute ischemia [20,21]. A variable hypoxia level is a better reproduction of tumor character, because as the tumor grows, its developing microregions differ in intensity of cell division, cell migration, necrosis and vascularization which directly influences therapeutic efficacy [22]. For this reason, we assessed the cytotoxic effect of temozolomide in cultures maintained in different models of hypoxia in a medium supplemented with different concentrations of glucose (0g/L; 1g/L;4.5g/L). In the first phase of experiments, the cultures were maintained in an atmosphere containing 10% O2 and then under chronic, acute and gradual ischemia and during hypoxia/reoxygenation. For the purpose of comparison, the experiment was also conducted under so-called “standard conditions” (in an atmosphere containing 20% O2), which are often used in research though are not found in the physiological conditions in the CNS. The obtained results indicated that the cytotoxic action of temozolomide on human glioblastoma cell line T98G was variable. The greater the oxygen availability was, at the same glucose concentration in culture medium, the more temozolomide reduced glioblastoma cell viability and the intensity of their division, thus enhancing apoptosis. In the acute ischemia model (glioblastoma cells were cultured in an atmosphere containing 1 or 2% O2 for 24 h), addition of temozolomide to the medium reduced cell viability and division by about 30–40% (depending on glucose concentration in the medium). Analogous results were obtained in the model in which O2 concentration in the atmosphere was lowered gradually to 1% O2. On the other hand, under chronic hypoxia (cells were incubated in an atmosphere containing 1% O2 for 10 days) temozolomide did not affect cell viability whereas in the hypoxia/reoxygenation model (cells were first incubated in an atmosphere containing 1% O2 for 72 h and then at 5% O2 for a further 72 h) temozolomide induced effects similar to those obtained in cultures conducted in an atmosphere containing 1% O2.
Our results showed unequivocally that the population of cells building the tumor mass located in chronic hypoxia regions was the most difficult to treat since they did not react to temozolomide. Therefore, it seems that the poor efficacy of temozolomide in some patients may stem not only from the anatomical localization of the tumor and its perfusion level but also from chronic hypoxia within the tumor [23]. Ischemia is significant to an understanding of the mechanisms of glioblastoma drug resistance. Reports indicate that necrotic foci often occurring within the tumor are among the most important causes of treatment failure, since they lower the sensitivity of hypoxic tumor tissues to available therapies [24]. Glioblastoma cells in chronic hypoxia regions develop an array of adaptive mechanisms promoting progenitor phenotype (with nestin expression), stabilizing HIF (hypoxia induced factor), reprogramming to pluripotent cells, reducing proliferation and enhancing cell migration, according to the “go or grow” dichotomy [25]. Activation and heterodimerization of HIF protein promote changes in the proteome and genome, leading to cell cycle arrest, disturbed cell proliferation, apoptosis, and also angiogenesis, vasculogenesis, increased production of lactate, lowered pH, elevated activity of DNA repair enzymes, an increased level of glutathione and the transcription factors TGF-β and bFGF, and, importantly, enhanced expression of proteins participating in glycolysis, which is the most important bioenergetic process for glioblastoma cells, responsible for their glucose-dependent character. These and other changes conditioned by HIF limit temozolomide penetration inside the tumor, which confirms the notion that “hypoxia helps glioma to fight therapy” [26-28]. It is also thought that temozolomide affects cells located in a more oxygenated tumor region (which is concurrent with our present results) but can also induce mutations and chemoresistance in cells situated in hypoxic regions. Therefore, it seems justified to supplement the temozolomide therapies used at present with oxygen therapy and antiangiogenic drugs [29,30].
The obtained results indicated explicitly that glucose concentration in the culture medium, even at different atmospheric oxygen levels, significantly influenced temozolomide cytotoxicity. Increased glucose concentration in the culture medium (4.5 g/L) was paralleled by reduced drug toxicity. We also observed that glucose elimination from the medium in each experimental model by itself (both under oxygenated and ischemic conditions) significantly reduced glioblastoma cell viability cell division and migration according to real-time imaging, enhanced apoptosis and led to a rapid loss of cell adhesion to the culture plate which can be significant for tumor relapse. Moreover, when cultures were maintained on the glucose-free medium, no differences were observed in study parameters between temozolomideexposed and unexposed cultures. The present data are concurrent with the results reported by other authors who have demonstrated a crucial role of glucose in the glioblastoma microenvironment as a readily available energetic fuel responsible for rapid tumor growth [31-33]. These data confirm the conclusion that “as long as glucose is available to glioblastoma cells, the tumor will grow unlimitedly causing longlasting therapeutic complications” at many levels. For this reason, the idea proposed by Zhou et al. [34] of pharmacologically inhibiting tumor glycolysis or replacing glucose with an alternative energy source, e.g. β-hydroxybutyrate (β-OHB) from ketone bodies and following dietary restrictions to limit glucose consumption, seems interesting. It should be remembered that glioblastoma cells have no mechanisms of metabolic flexibility and, unlike healthy cells, cannot utilize energy sources other than glucose due to their glucose-dependence [35,36]. It has even been suggested that therapies limiting tumor energy metabolism could be more efficient (their antiangiogenic, antiinflammatory activity and ability to increase the bioenergetic potential of healthy brain cells have been discovered) and less toxic for patients than currently used therapeutic schemes [37]. However, this idea raises some doubts due to its “simplistic” mechanism.
We are aware that it is risky to unquestioningly apply results of in vitro studies on a cell line to carcinogenesis in the central nervous system. In the light of our data and literature reports, it appears that both proper metabolic compensation, especially diabetes control and modification of the antitumor pharmacotherapeutic scheme are important to improve therapeutic efficacy [38]. Chronic use of glucocorticosteroids as an adjuvant therapy in GBM patients increases the risk of hyperglycemia and consequently enhances energy metabolism in glioblastoma cells which can contribute to the escalation of carcinogenesis, thereby reducing patient survival rate [39]. It has been proven that routine intravenous administration of high doses of dexamethasone to decrease edema and compression of brain structures by growing tumor leads to acute hyperglycemia initiating hyperinsulinemia which stimulates tumor growth [40]. Results of clinical studies indicate that an elevated level of insulin enhances cell proliferation in breast and colon cancers [41,42]. The role of insulin in glioblastoma development may also be significant due to the presence of insulin receptors on tumor cells [43].
The high extracellular glucose concentration promoting aggressive glioblastoma phenotype may be one of the causes of glioblastoma resistance to temozolomide therapy [44]. It has been shown that long-term temozolomide treatment leads to increased expression of glucose transporters in glioblastoma cells (GLUT-3 and AKR1C family proteins), which may explain the sudden progression of the disease in some glioblastoma types after the commencement of therapy with this drug [45]. Resistance to temozolomide treatment may also be a result of the histological structure of the tumor. It is believed that patients with glioblastomas of astrocytic origin have a worse prognosis, while those suffering from oligodendroce-derived tumors a better one due to their sensitivity to chemotherapy [46]. Astrocytes play an important role in glucose metabolism and generation of high-energy compounds for the CNS, and, importantly, participate in protective mechanisms during acute or chronic hypoglycemia by activating the pentose phosphate pathway which leads to a reduction in oxidative stress facilitating tumor development [47].
It can be expected that suppression of the intense energy metabolism of the glioblastoma should improve therapy efficacy. Great hopes are invested in a new therapy which is to work like “dumdum weapon”; the idea exploits the high preference of glioblastoma cells for glucose, and is based on nanoparticles coated with a polysaccharide substance with a chemotherapeutic closed inside. However, we have yet to discover whether this idea will prove efficient.
The results of our studies on glioblastoma cell line T98G demonstrated that a constant presence of high glucose levels in the glioblastoma cell microenvironment can have crucial impact on carcinogenesis progression by increasing cell division or migration, and thus favoring resistance to the drug. Evaluation of apoptosis in the acute and chronic hypoxia models revealed that variable oxygenation of glioblasoma cells is vital to the acquisition of chemo-resistance by these cells. The significant differences between the results of experiments carried out in hypoxic conditions and those performed in „standard conditions” call into question the reliability of the latter in vitro experimental approach to glioblastoma studies.
Dr. Anna M Bielecka (the first author) planned and performed all experiments, prepared the manuscript and designed the concept of this experimental work (connected with glioblastoma multiforme). Prof. Ewa Obuchowicz checked this text and made corrections.
This work was supported by the grant KNW-1-013/N/2/0 from the Ministry of Science and Higher Education.