Journal of Horticulture
Open Access
ISSN: 2376-0354
ISSN: 2376-0354
Research Article - (2016) Volume 3, Issue 3
The polymerization or depolymerization of the cytoskeleton is very important in various aspects of plant development. ADF (actin-depolymerizing factor) is a typical actin binding protein that can mediate the polymerization or depolymerization of the actin cytoskeleton, but its role in fruit ripening remains unclear. Here, we characterized an ADF gene, PuADF. The expression of PuADF was evaluated in young leaves, stems, flowers, and roots as well as in fruits. The expression of PuADF was induced by ethephon treatment but inhibited by 1-MCP treatment. To explore the network of PuADF function in ‘Nanguo’ pear fruits ripening, we screened a cDNA library from ‘Nanguo’ pear fruits using PuADF as bait. Two potential interactors of PuADF were identified, PuAS (Asparagine Synthetase) and PuDAD1 (Defender Against Death 1), both of which have been associated with PCD (Programmed Cell Death). This direct interaction was further confirmed by yeast two-hybrid analyses. The expression levels of PuAS and PuDAD1 were affected by ethylene. Our results indicated that PuADF is involved in ethylene-mediated fruit ripening and might be related to PCD during fruit ripening. The possible mechanisms are discussed.
<
Keywords: ADF; Cytoskeleton; Fruit ripening; PCD; Pyrus ussuriensis
The cytoskeleton is part of the cellular structure and plays important roles in maintaining cell shape, in trafficking of various materials, and in cell division. In particular, its role in cell signal transduction has attracted increasing attention. The actin cytoskeleton has high instability, meaning that it can polymerize and depolymerize in response to the environment both in vivo and in vitro . These cytoskeletal proteins can respond to signal stimuli, such as hormones and pathogens, to regulate cell physiology. The actin filaments of Commelina communis L. guard cells are localized in the cortex of these cells, arranged in a regular pattern, radiating from the stomatal pore; however, with abscisic acid treatment, the actin began to disintegrate within a few minutes and was completely disintegrated in 1 h [1]. In vetch root hairs, the number of actin filaments increased in 3 to 15 min after treatment with a host-specific Nod factor [2].
ADF (actin-depolymerizing factor) is a typical actin binding protein that can cause the depolymerization of actin filaments [3]. Several ADFs have been demonstrated to function in severing actin filaments [4-7]. In the emergent and elongated maize root hairs, ZmADF3 was shown to redistribute from a diffuse to a tip-concentrated localization [8], which is similar to a previous study in which ADFs were localized to the leading edge in other migrating cells [9].
There are nine ADFs in Arabidopsis [10-13]. A previous study reported that the normal actin cytoskeleton in Arabidopsis was disrupted by overexpression of AtADF1 and suggested that AtADF1 may play a role in cell expansion, organ growth and flowering [13]. The GFP-AtADF1 fusion protein has also been shown to bind to actin filaments and affect F-actin organization in transformed BY2 cells [12]. In tobacco, there is a pollen-specific ADF, NtADF1. GFP-NtADF1 was shown to localize in the subapical region in elongating pollen tubes. The actin cytoskeleton was disrupted and the growth rates were reduced in the elongating pollen tubes in which NtADF1 was overexpresse [9]. These results indicate that NtADF1 plays an important role in regulating the balance of actin polymerization and depolymerization in pollen tube growth process [9]. At present, studies on ADF have mainly focused on the existing polarized growth of pollen tubes and root hairs, whereas the roles of these proteins in other organs remain unclear.
In banana, the cytoskeleton protein actin bundling protein ABP135 showed different expression levels during the different peel color index (PCI) ripening stages [14]. Moreover, it has been reported that cell shape also changes greatly during fruit ripening; for example, the cells of ‘Gala’ apple fruits became much bigger during storage, showing swelling of the cell wall and a loose arrangement [15]. These results indicated that the proteins maintaining the cytoskeleton might degrade and be unable to support the normal cell shape during fruit softening. ADF is an actin binding protein and is known to function in certain processes, but whether ADF is involved in fruit ripening and softening remains unclear.
The ‘Nanguo’ pear (Pyrus ussuriensis ) fruit is typically climacteric, and ethylene is the main factor controlling the ripening of climacteric fruit. During ripening, the fruits produce a large amount of ethylene, along with a rapid drop in firmness [16]. Previously, we compared the transcriptomes of pre- and post-climacteric fruits of ‘Nanguo’ pear, and we found that the expression level of an ADF gene, named PuADF, was down-regulated during fruit ripening. Here, we detected the expression pattern of PuADF during fruit development and ripening, as well as in ethylene- and 1-MCP-treated fruits. Moreover, we screened a Y2H (yeast two-hybrid) cDNA library constructed with ‘Nanguo’ pear fruits cDNA and obtained two proteins that interacted with PuADF, PuAS and PuDAD1, which are thought to function in programmed cell death (PCD).
Plant materials and treatments
Young leaves, stems, flowers and fruits were collected from mature ‘Nanguo’ pear (P. ussuriensis ) trees that were grafted on rootstocks of ‘Shanli’ (P. ussuriensis maxim) and growing at the experimental farm of Shenyang Agricultural University (Shenyang, China). Roots were collected from ‘Shanli’ pears. Fruits were sampled every 15 d from 30 days after full bloom (DAFB) to commercial harvest. The fruits harvested at 145 DAFB (commercial maturity, Sep. 16, 2014) were stored at room temperature (RT, 24°C) for 15 d and sampled every 5 d. In addition, two other groups of ‘Nanguo’ fruits collected at 145 DAFB were subjected to 1 μL/L of 1-MCP (1-methylcyclopropene, an ethylene inhibitor) for 24 h and 1000 ppm ethephon solution (an ethylene producer) for 15 s in an airtight container according to Tan et al. [17]. After treatment, the fruits were held at RT for 15 d and sampled every 5 d for RNA extraction. At each sampling point, 5 fruits were sampled for measurement of ethylene production and flesh firmness, and then fruits were sliced, pooled and frozen in liquid N2, and stored at -80°C for later use.
Measurement of ethylene production rates and flesh firmness
The measurements of flesh firmness and ethylene production rates were performed according to Tan et al. [17]. For measurement of ethylene production rates, intact fruits were enclosed in an airtight container (0.86 L, 24°C) equipped with septa, and 1 ml of headspace gas was sampled using a syringe. The ethylene concentration was measured with a gas chromatograph (Agilent 7890A, USA) equipped with a flame ionization detector according to Tan et al. [18]. Three fruits per sample were measured.
Flesh firmness was measured using a portable pressure tester (FT-327, Facchini, Italy) fitted with an 11-mm-diameter probe. Four skin discs (approximately 2.5 cm in diameter) were removed from opposite sides of each fruit. The probe was pressed into the tissue of the cut surface to a depth of 8-9 mm in a single smooth motion. Measurements from three fruits were pooled for each sample.
Quantitative RT-PCR
Total RNAs were extracted by using the modified CTAB method [19]. Then 1 μg of total RNA was used to synthesize first-strand cDNA using a PrimeScript First Strand cDNA Synthesis Kit (Takara, Japan).
Quantitative RT-PCR (qRT-PCR) was conducted as described by Tan et al. [17]. Specific primers for each gene were designed using Primer5 and are listed in Supplementary Table S1. The pear Actin gene was used as an internal control. Three replications were conducted.
Sequence analysis
The gene BLAST search and structure analysis were performed using the National Center for Biotechnology Information database (NCBI, http://www.ncbi.nlm.nih.gov/) and the Genome Database for Rosaceae (GDR, https://www.rosaceae.org/node/1). A phylogenetic tree for each gene was generated by using the NJ method with 1000 bootstrap replicates in MEGA version 5.0 [20].
Screening of the Y2H library
The ORF of PuADF was cloned into the pGBKT7 vector (Clontech) and confirmed by sequencing with T7 primers. Then, the resulting recombinant plasmid was transformed into the yeast strain Y2H Gold using the “Yeastmaker Yeast Transformation System 2” kit (Clontech).
The pGBKT7-PuADF was used as bait to screen the Y2H library with the GAL4-based “Matchmaker Gold Yeast Two-Hybrid System” kit (Clontech) according to the manufacturer’s instructions. Briefly, 4 ml of the bait strain overnight cultures (SD/-Trp liquid medium), 1 ml of library aliquot (1.2 × 107 cells) and 45 ml 2 × YPDA medium were combined in a 2 L flask and incubated at 30°C for 20 h with shaking at 50 rpm and then checked for the formation of zygotes. If zygotes appeared, the cells were collected and resuspended in 10 ml of 0.5 × YPDA medium. The mated cell cultures were spread on the DDO/X/A plates (containing double dropout SD medium that lacks tryptophan and leucine but is supplemented with X-α-Gal) and kept at 30°C. Positive colonies that appeared within 5 d were streaked onto QDO/X/A plates for high stringency screening. The colonies appeared within 5 d on these QDO/X/A plates were predicted to be positive hybrids, and then the plasmids were rescued using the “Easy Yeast Plasmid Isolation” Kit (Clontech). Then, they were transformed into the E. coli strain TOP10 and plated on LB plates with 100 mg/ml ampicillin to select the prey plasmids. To reveal the identity of the prey plasmids, the inserts were sequenced using the T7 primer.
Yeast two-hybrid assays
The CDS of PuAS and PuDAD1 were introduced into pGADT7 (Clontech, CA, USA). These AD vectors were separately cotransformed with the pGBKT7-PuADF vector into AH109 cells. After being grown on SD/-Leu/-Trp medium at 30°C for 3-4 d, the clones were subsequently cultured on SD/-Ade-His-Leu-Trp at 30°C for 3-4 d and then stained with X-α-gal (Clontech) to detect any interactions.
Cloning and expression analysis of PuADF
‘Nanguo’ pear (P. ussuriensis ) fruits undergo typical respiratory climacteric during ripening, with firmness dropping rapidly and producing large amounts of ethylene [16]. In our previous transcriptomes analysis, we obtained a fragment of an ADF gene from ‘Nanguo’ pear, and this fragment showed down-regulation during fruit ripening. Using this fragment as bait, we screened the pear genome and got the full length sequence of this gene. Then we designed fulllength primers and cloned the full-length cDNA and genomic DNA of this gene from ‘Nanguo’ pear fruits. The sequence was found to be identical to one obtained from the NCBI database, and this gene was named PuADF. Its cDNA is 438 bp in length and encodes 145 amino acids. Its genomic DNA contains 2 exons (286 bp and 152 bp) and 1 intron (663 bp) (Figure 1A). To examine the evolution of PuADF protein, we analyzed the phylogenetic relationship of PuADF and its homologs from other plants. First, we got the homologs of PuADF from the NCBI database as following method: using the PuADF amino acid sequence as bait, we screened the NCBI database and selected the genes whose homology were more than 40% to PuADF to build a NJ tree using the MEJA 5.0. Our results showed that PuADF could be clustered together with MdADF (Figure 2).
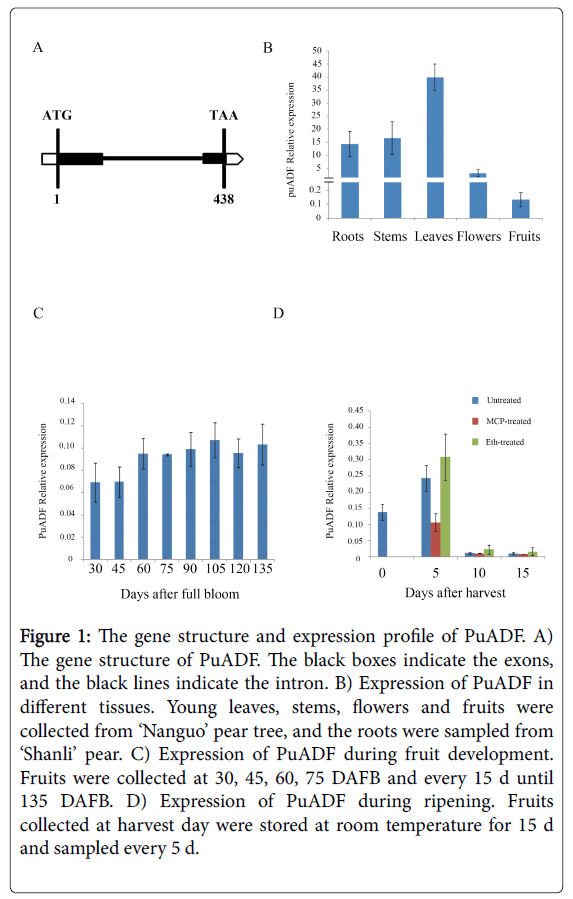
Figure 1: The gene structure and expression profile of PuADF. A) The gene structure of PuADF. The black boxes indicate the exons, and the black lines indicate the intron. B) Expression of PuADF in different tissues. Young leaves, stems, flowers and fruits were collected from ‘Nanguo’ pear tree, and the roots were sampled from ‘Shanli’ pear. C) Expression of PuADF during fruit development. Fruits were collected at 30, 45, 60, 75 DAFB and every 15 d until 135 DAFB. D) Expression of PuADF during ripening. Fruits collected at harvest day were stored at room temperature for 15 d and sampled every 5 d.
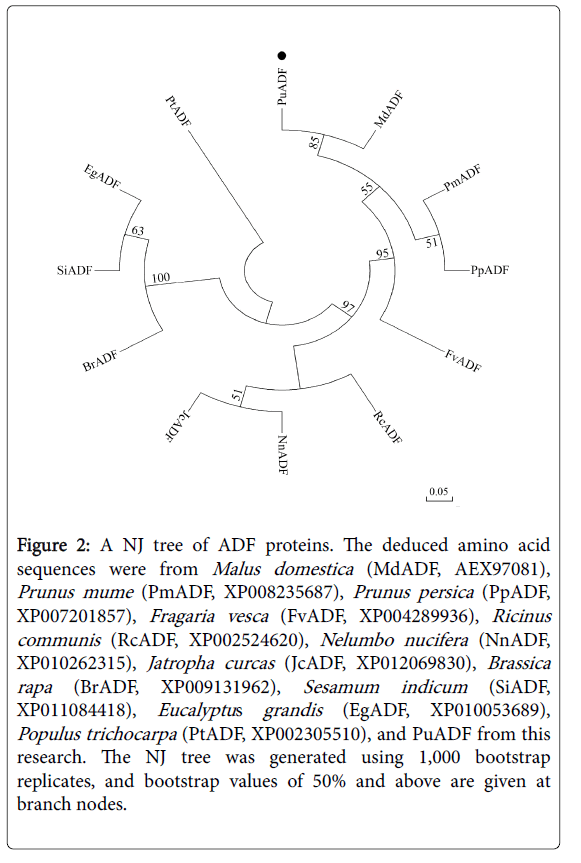
Figure 2: A NJ tree of ADF proteins. The deduced amino acid sequences were from Malus domestica (MdADF, AEX97081), Prunus mume (PmADF, XP008235687), Prunus persica (PpADF, XP007201857), Fragaria vesca (FvADF, XP004289936), Ricinus communis (RcADF, XP002524620), Nelumbo nucifera (NnADF, XP010262315), Jatropha curcas (JcADF, XP012069830), Brassica rapa (BrADF, XP009131962), Sesamum indicum (SiADF, XP011084418), Eucalyptus grandis (EgADF, XP010053689), Populus trichocarpa (PtADF, XP002305510), and PuADF from this research. The NJ tree was generated using 1,000 bootstrap replicates, and bootstrap values of 50% and above are given at branch nodes.
The expression profile of PuADF was then examined in roots, stems, leaves, flowers and fruits of ‘Nanguo’ pear. PuADF was expressed in all the tissues examined, with the expression level in leaves being much higher than in other tissues (Figure 1B). PuADF was also expressed in fruits, although at a very low level (Figure 1B).
The expression level of PuADF showed no obvious differences during the development of ‘Nanguo’ pear fruits (Figure 1C). To understand whether ethylene influences the expression of PuADF, we treated ‘Nanguo’ pear fruits with ethephon or 1-MCP (a C2H4 antagonist). In untreated fruits, the PuADF expression level was highest at 5 days after harvest (DAH) and then decreased rapidly during the storage period (Figure 1D). In the ethephon-treated fruits, the expression of PuADF was greatly induced (Figure 1D); while in the 1-MCP-treated fruits, the expression level of PuADF was inhibited (Figure 1D). These results showed that PuADF was influenced by ethylene and might be related to fruit ripening.
Screening of the Y2H library
To explore the network of PuADF function in ‘Nanguo’ pear fruit ripening, we used PuADF as bait for screening a Y2H library. Two genes showed great abundance, appearing in 10 out of 50 samples. One belongs to the AS (Asparagine Synthetase) family and the other one belongs to the DAD (Defender Against Death) family. We cloned the full-length cDNAs of these two genes and named them PuAS and PuDAD1. PuDAD1 is 360 bp in length, with no introns, and encodes 119 amino acids; it contains a DAD domain and can be clustered together with PpDAD1 (Prunus persica ) and PmDAD1 (Prunus mume ) (Supplementary Figure 1A and Supplementary Figure 2). PuAS is 1761 bp long, contains 10 exons and 9 introns (Supplementary Figure 1B), and can be clustered together with MdAS (Malus domestica ) (Supplementary Figure 3).
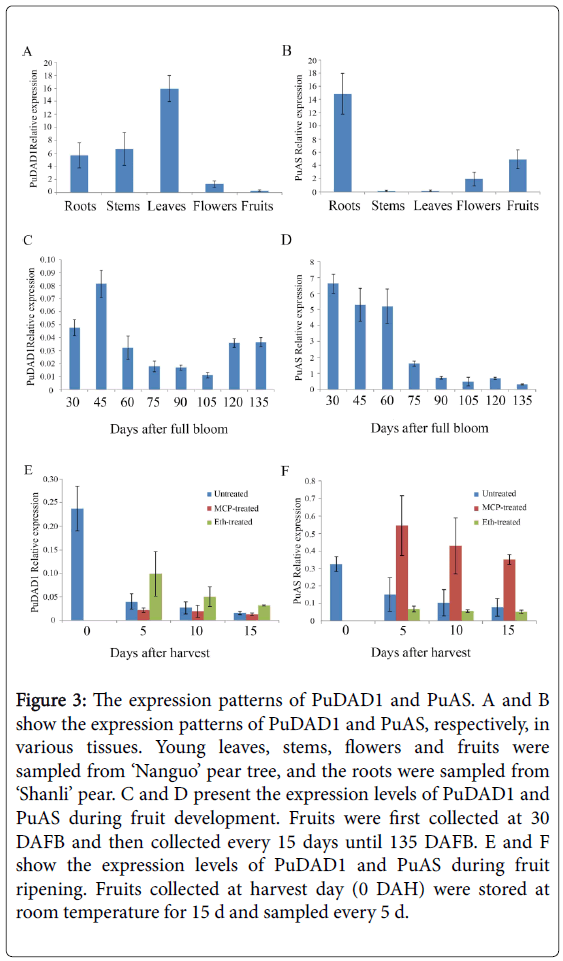
Figure 3: The expression patterns of PuDAD1 and PuAS. A and B show the expression patterns of PuDAD1 and PuAS, respectively, in various tissues. Young leaves, stems, flowers and fruits were sampled from ‘Nanguo’ pear tree, and the roots were sampled from ‘Shanli’ pear. C and D present the expression levels of PuDAD1 and PuAS during fruit development. Fruits were first collected at 30 DAFB and then collected every 15 days until 135 DAFB. E and F show the expression levels of PuDAD1 and PuAS during fruit ripening. Fruits collected at harvest day (0 DAH) were stored at room temperature for 15 d and sampled every 5 d.
Expression profiles of PuDAD1 and PuAS
We next examined the expression of PuDAD1 and PuAS in young leaves, stems, flowers, fruits, and roots. PuDAD1 was expressed in all the tissues examined, and the highest expression level was observed in leaves. However, in fruits, PuDAD1 expression was very low (Figure 3A). PuAS was also expressed in all the tissues examined, but its highest expression was in roots, followed by fruits, and then the other three tissue types studied (Figure 3B).
Then, the expression levels of these two genes were examined in ‘Nanguo’ pear fruits at different stages during development and ripening. The expression level of PuAS decreased gradually with the progress of fruit development (Figure 3D), whereas PuDAD1 expression showed no consistent pattern but was expressed at very low levels throughout the stages of fruit development (Figure 3C).
The effects of ethylene and 1-MCP treatments on PuDAD1 and PuAS
To understand whether the expression of PuDAD1 and PuAS is under the control of ethylene, we evaluated the expression levels of these two genes in 1-MCP- and ethephon-treated ‘Nanguo’ pear fruits. Compared with untreated fruits, ethephon greatly increased the expression of PuDAD1 at all sampling points, whereas 1-MCP did not significantly affect PuDAD1 expression (Figure 3E). In contrast, the expression of PuAS was greatly increased by 1-MCP and not significantly affected by ethephon (Figure 3F).
Confirmation of the interaction between PuADF and PuAS or PuDAD1
The CDS of PuAS and PuDAD1 without signal peptides were introduced into the pGADT7 vector, and then each plasmid was cotransformed with pGBKT7-PuADF into Y2H Gold. As shown in Figure 4, PuADF could interact with PuAS or PuDAD1 in yeast cells.
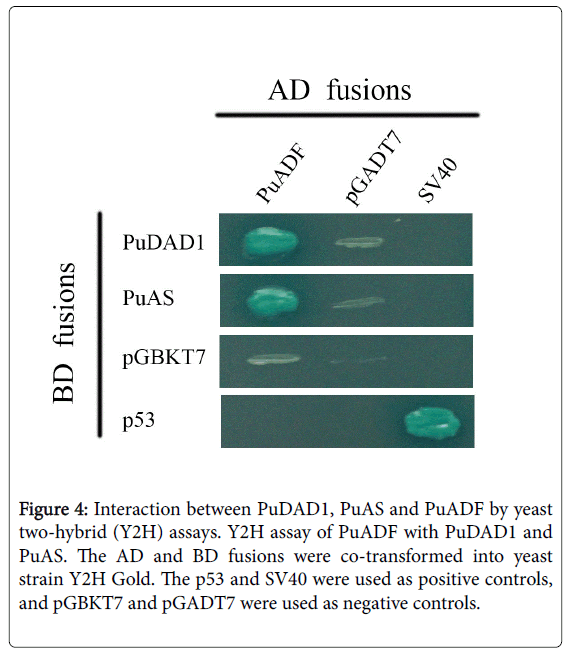
Figure 4: Interaction between PuDAD1, PuAS and PuADF by yeast two-hybrid (Y2H) assays. Y2H assay of PuADF with PuDAD1 and PuAS. The AD and BD fusions were co-transformed into yeast strain Y2H Gold. The p53 and SV40 were used as positive controls, and pGBKT7 and pGADT7 were used as negative controls.
Fruit softening is the most important sign that the fruit is beginning to break down, and the softening rate directly determines the fruit storage quality. ‘Nanguo’ pear is a typical climacteric fruit that produces high levels of ethylene during ripening, accompanied by a rapid decrease in firmness [16]. Organ senescence is an example of PCD in plants [21], and fruit ripening is considered a specialized form of senescence [22]. Interestingly, a putative apoptosis-related protein 19 was up-regulated through the banana PCI (peel color index) ripening stages [14].
ADF proteins belong to the family of actin binding proteins and can mediate the depolymerization of actin filaments [3]. In the AtADF1 overexpression lines, the normal actin cytoskeleton in Arabidopsis was disrupted [13]. With the overexpression of the pollen-specific NtADF1, the pollen tube actin cytoskeleton was disrupted, and the growth rate was reduced [9]. In our study, the expression level of PuADF was down-regulated during the storage period, in a pattern similar to the decrease in firmness. These results indicated that much of the actin cytoskeleton might have been depolymerized during the storage of ‘Nanguo’ pear fruits and that the remaining actin cytoskeleton was unable to maintain the fruit structure, contributing to the decrease in fruit firmness. Further, the expression level of PuADF could be induced by ethephon and inhibited by 1-MCP treatment, indicating its involvement in ethylene-mediated fruit ripening, and the low expression level of PuADF might delay the depolymerization of actin filaments and slow the drop in fruit firmness. However, more evidence is necessary to confirm the role of PuADF in maintaining fruit firmness.
Dramatic actin rearrangements are linked to the progression of PCD either by triggering PCD initiation or by modulating its progress [23]. In the Trichoplusia ni BTI-Tn-5B1-4 cell line, azadirachtin could induce programmed cell death, accompanied by cytoskeletal damage [24]. It has been demonstrated that the actin depolymerization induced by self-incompatibility activated a PCD network [25]. In Picea abies, when the actin microfilaments are disrupted, the speed of tube cell PCD increases, and the formation of the suspensor is inhibited [26].
To better understand the function of PuADF, we screened a ‘Nanguo’ pear fruits cDNA library using PuADF as bait. We obtained two proteins that interacted with PuADF, PuDAD1 and PuAS. DAD1 proteins have been shown to be conserved among human [27], mouse [28], Caenorhabditis elegans [29], Arabidopsis thaliana [30], pea [31] and other species. In mammals, DAD1 is thought to function in protecting cells against PCD (apoptosis) [32]; for example, a hamster cell line shows increased apoptosis when DAD1 protein is lost [32]. In Arabidopsis, the expression level of DAD1 was reduced during the development of siliques [30]. Similarly, in pea, the DAD1 expression level decreases during petal senescence, a process that involves PCD [31]. The expression of MdDAD1 is induced during senescence of leaves, petals and fruit [32]. Here, PuDAD1 was down-regulated during the storage period and could be induced by ethephon treatment. These results suggest the possibility that ethylene might pass the signal directly or indirectly via PuADF through its protein interaction with PuDAD1, resulting in PCD and accelerating fruit softening. Although Pennell and Lamb [33] have proposed that ethylene can act in concert with other signal molecules to control PCD, confirmation of that possibility requires additional investigation.
AS was thought to be involved in protein metabolism and might be associated with senescence programs [34]. The difference between ripening and senescence has yet to be well described [35], and both processes may contribute to the softening of fruit. In our study, PuAS expression was increased by 1-MCP treatment (Figure 3F), suggesting that it may contribute to maintaining the firmness of fruit during ripening. Moreover, PuAS could interact with PuADF, and the expression levels of both proteins decreased during fruit ripening, along with the decrease in firmness. These findings provide a new link between fruit ripening and senescence.
In conclusion, PuADF is involved in ethylene-mediated fruit ripening and can interact with PuAS and PuDAD1, both of which are involved in PCD.
This work was supported by the Innovative Program for Fruit Tree Research of Liaoning Province, China (LNGSCYTX-13/14-4).