Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Case Report - (2013) Volume 2, Issue 1
Varying concentrations of Chlorpromazine Hydrochloride (CPZ) were exposed to a 266 nm laser beam for varying periods of time ranging from 4 to 24 hrs and the products of irradiation were evaluated for activity against a panel of bacteria that consisted of representatives of Gram-positives and Gram-negatives that expressed different degrees of efflux pump activity, and compared to the parental unexposed compound with prolonged irradiation, whereas the antibacterial activity of the product against Staphylococcus aureus and Escherichia coli strains was many folds greater, no activity against their efflux pumps was noted. The activity of the products of irradiation against Salmonella enterica serovar Enteritidis was slight. However, the products of prolonged irradiation of CPZ produced increasingly significant concentration dependent inhibition of efflux by the Salmonella strains.
<Keywords: Laser irradiation; Photodegradation; Chlorpromazine (CPZ); Escherichia coli; Salmonella enterica serovar Enteriditis; Antimicrobial activity; Inhibition of efflux pumps.
Exposure of compounds to a high energy laser beam for varying periods of time is known to increase their biological activity [1-3]. Exposure of Chlorpromazine Hydrochloride (CPZ) to a 266 nm laser beam has been shown to increase its activity against a reference strain of Staphylococcus aureus ATCC 25923 [3]. The study has been extended to include a larger range of concentrations of CPZ exposed to prolonged periods of time to a 266 nm laser beam at an average energy of 6.5 mJ. The products of irradiation were examined for altered antibacterial activity against panels of Gram-positive and Gram-negative bacteria that differed with respect to the expression of their efflux pumps as well as for activity against their efflux pumps. The results of this study support the idea that exposure of an antibacterial agent to a high energy laser beam at a wavelength that matches the maximum absorbance of the compound is a prospective way to obtain molecules with increased antibacterial activity as compared to the parental compound. With respect to activity against the efflux pumps of the studied bacteria, the products produced from the irradiation of CPZ inhibited the efflux pump of the Salmonella strains in a concentration dependent manner and the degree of inhibition related to the prolongation of irradiation of CPZ.
Culture media and reagents
Mueller-Hinton (MH) (Sigma, Madrid, Spain), Luria Bertani (LB) and Tryptic Soy Broth (TSB) media Oxoid (Basingstoke, Hampshire, UK) were purchased in powder form. Stock solution of Ethidium Bromide (EB) (Sigma Aldrich) at 100 mg/L prepared in distilled water. CPZ from Sigma (Madrid, Spain), higher than 98.9% pure, was dissolved in distilled water to yield a concentration of 20 mg/mL immediately before use and protected from environmental (natural and/or artificial) light.
Bacterial strains
The panel of bacteria for evaluation of products of irradiation of CPZ consisted of Gram-positive wild-type Staphylococcus aureus American Type Culture Collection (ATCC) 25923 and Staphylococcus aureus HPV 107(representative of the MRSA Iberian clone; it was isolated at a Portuguese hospital in 1992 and is characterized by resistance to several classes of antibiotics, particularly ß-lactams, aminoglycosides, fluoroquinolones, macrolides, rifampicin and tetracycline [4,5]). The ATCC wild-type strain has an intrinsic efflux pump system and the HPV 107 strain has a plasmid containing the QacA gene which renders the bacterium multidrug resistance [6].
The Gram-negative bacteria are Escherichia coli K-12 AG100 (wildtype), Escherichia coli K-12 AG100A (efflux pump AcrAB-TolC deleted [7]), Escherichia coli AG100TET8 (induced to high level resistance to tetracycline and over expresses its AcrAB-TolC) [7]; Salmonella enterica serovar Enteritidis NCTC 13349 (wild-type), Salmonella enterica serovar Enteritidis 104 (clinical strain), Salmonella enterica serovar Enteritidis 5408 (clinical stain), Salmonella Enteritidis 104CIP and 5408CIP strains were derived from their respective parental strains by gradual exposure to ciprofloxacin which resulted in resistance to this antibiotic [8]. Whereas induced resistance of these progeny strains was due from over-expression of the acrB transporter of the acrAB-TolC efflux pump [8], with respect to the 104cip, resistance was in part due to the presence of a mutation in gyrA and two mutations in the stress gene sox [8].
Minimum inhibitory concentration (MIC)
MIC of the compounds were determined by the microplate broth microdilution method according to Clinical and Laboratory Standards Institute (CLSI) recommendations [9] and is defined as the lowest concentration of compound for which no visible growth is present. The MIC for each compound was determined at least three separate times.
Real-time ethidium bromide accumulation assay
The real-time activity of the compounds against efflux pumps of the panel of bacteria at concentrations at and below ½ their MIC has been described in detail [10]. This fluorometric method employs a Rotor- Gene™ 3000 Thermocycler with real time analysis software (Corbett Research, Sydney, Australia) that follows the accumulation of the universal substrate EB [9] and the data interpreted represent changes in the degree of fluorescence produced due to effects of the compounds on the activity of the efflux pump [9]. Briefly, the strains were cultured in appropriate medium until they reached an optical density at 600 nm (OD600) of 0.6 and were then centrifuged at 13,000 rpm for 3 minutes. Pellets were resuspended in Phosphate-Buffered Saline (PBS), washed twice and resuspended in PBS containing glucose (concentration of 0.4%). The OD600 was adjusted with PBS to 0.6, and aliquots of 45 μL were transferred to microtubes of 0.2 mL volume. 5 μL of distilled, sterile water blank control or 5 μL of the compounds dissolved in water at different concentrations was individually added to the tubes, followed by the addition of 45 μL of PBS containing a concentration of EB adjusted for different strains and by 50 μL of bacterial cells. The amount of fluorescence representing accumulated EB by the bacterium was monitored on a real-time basis and the graphs obtained reflected the degree of fluorescence generated per unit period of time.
To determine efflux-modulation activity, it is necessary to use a concentration of any potential efflux pumps inhibitor that is ½ MIC or below [11], which does not affect the viability of the bacterium [12]. The MIC of the products of prolonged irradiation of CPZ against the panel of Gram-positive and Gram-negative bacteria is described by table 1; the values of the results of MIC for the irradiated compounds that present a significant antimicrobial activity against bacteria than the un-irradiated one are highlighted in this table.
| Strains | CPZ unirradiated | CPZ irradiated 4 h | CPZ irradiated 8 h | CPZ irradiated 16 h | CPZ irradiated 24 h |
|---|---|---|---|---|---|
| MIC (mg/L) | MIC (mg/L) | MIC (mg/L) | MIC (mg/L) | MIC (mg/L) | |
| Staphylococcus aureus ATCC 25923 | 25 | 3.125 | 3.125 | 3.125 | 3.125 |
| Staphylococcus aureus HPV 107 | 25 | 3.125 | 3.125 | 3.125 | 3.125 |
| Escherichia coli K-12 AG100 | 25 | 3.125 | 3.125 | 3.125 | 3.125 |
| Escherichia coli K-12 AG100A | 25 | 1.56 | 1.56 | 1.56 | 1.56 |
| Escherichia coli AG100TET8 | 50 | 3.125 | 3.125 | 3.125 | 3.125 |
| Escherichia coli AG100ATET8 | 50 | 6.25 | 6.25 | 6.25 | 6.25 |
| Salmonella Enteritidis NCTC 13349 | 50 | 25 | 25 | 25 | 25 |
| Salmonella Enteritidis 104 | 50 | 25 | 25 | 25 | 25 |
| Salmonella Enteritidis 104CIP | 100 | 25 | 25 | 25 | 25 |
| Salmonella Enteritidis 5408 | 50 | 25 | 25 | 25 | 25 |
| Salmonella Enteritidis 5408CIP | 50 | 25 | 25 | 25 | 25 |
Table 1: Minimum Inhibitory Concentrations (MIC) of the compounds for Gram-negative and Gram-positive strains.
• The MIC for CPZ irradiated for 4, 8, 16, and 24 hrs against the S. aureus ATCC 25923 and HPV 107 was 8 times lower than that of the un-irradiated CPZ control.
• The MIC for CPZ irradiated for 4, 8, 16, and 24 hrs against the wild-type E. coli K-12 AG100 was 8 times lower than the one obtained for the un-irradiated CPZ control.
• The MIC for the above irradiated CPZ products against E. coli strains K-12 AG100A, AG100TET and AG100ATET was as much as 16 fold lower than the one obtained with the un-irradiated CPZ control.
• For Salmonella Enteritidis strains, the above irradiated CPZ products produced a marginal two fold decrease (barely significant) in the MIC as compared to the un-irradiated CPZ.
These results suggest that the activity of irradiated products of CPZ is significantly greater than that produced by the un-irradiated control CPZ.
The study of accumulation of the effect of irradiated products of CPZ on the retention of EB (inhibition of efflux) was conducted at ½ their MICs and demonstrated that there is no significant activity against the efflux pumps of S. aureus ATCC 25923 and S. aureus HPV 107 and those of E. coli K-12 AG100, K-12 AG100A, AG100TET8 and AG100ATET8 (data not shown). It should be noted that concentrations of the irradiated CPZ products above the MIC did inhibit efflux (data not shown).With respect to the effect of the products of irradiation on the efflux pump of the Salmonella strains, as evident by the example provided by figure 1 demonstrating an inhibition of efflux of the Salmonella 104. As noted by figure 1, the products produced from the irradiation of CPZ inhibited the efflux pump of the Salmonella strains and with exception of the 24 hr irradiated product, the degree of inhibition was related to the prolongation of irradiation of CPZ. This was also true for the other strains. Not shown is the concentration dependent inhibition of efflux by the most prolonged irradiation products of CPZ. It should be noted that the effect of the products of irradiation appear to be more pronounced on the efflux pump of the 104 parental strain than that of its ciprofloxacin induced over-expressed AcrB transporter progeny 104CIP. The reason for this is that because the latter strain has a far greater number of efflux pumps, inhibition requires higher amounts of inhibitor. Unfortunately, this basic understanding is lacking in most studies of inhibitors of efflux pumps of bacteria.
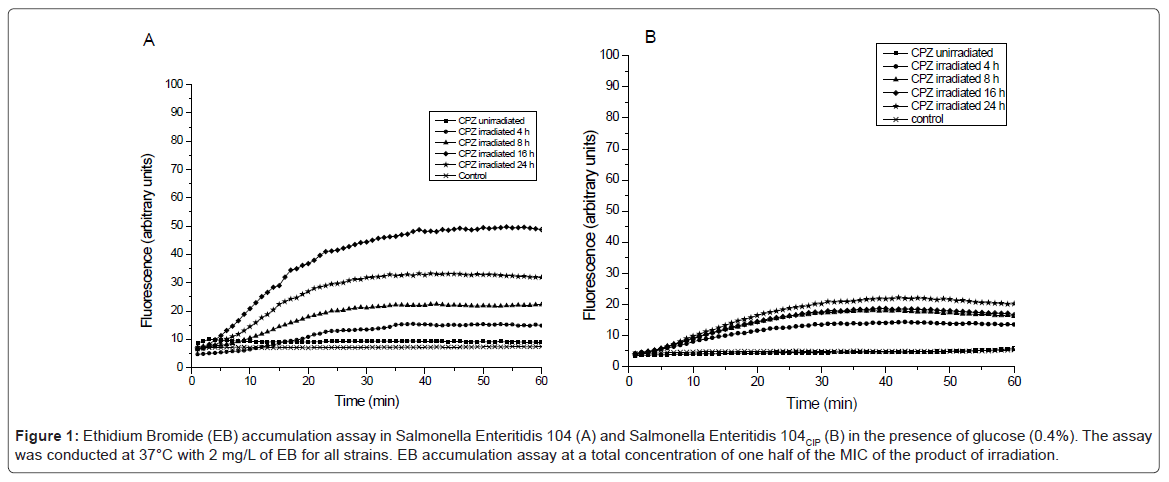
Figure 1: Ethidium Bromide (EB) accumulation assay in Salmonella Enteritidis 104 (A) and Salmonella Enteritidis 104CIP (B) in the presence of glucose (0.4%). The assay was conducted at 37°C with 2 mg/L of EB for all strains. EB accumulation assay at a total concentration of one half of the MIC of the product of irradiation.
The results obtained in this study suggest that the effect of irradiating a compound, in our case CPZ, with a 266 nm laser beam generates new species, and with exception of the Salmonella enterica serovar Enteritidis strains, the products of the irradiated CPZ demonstrated greater antimicrobial properties than the un-irradiated product against the bacteria employed in this study Although the products of irradiation produced a barely significant reduction of the MIC of CPZ, they inhibited the efflux pump systems only of Salmonella enterica serovar Enteritidis.
Most of medicinal compounds, developed during the 20th century have their origins in phenothiazines [13]. Phenothiazines at relatively high concentrations have activity against bacteria, where they can mediate effects such as: direct inhibition of replication [14], reduced antimicrobial resistance via increased drug efflux [15], inhibition of bacterial motility [16],enhanced killing of intracellular bacteria [17], elimination of plasmids [18], and inhibition of efflux pumps of Grampositive and Gram-negative bacteria[19].
Phenothiazines can eliminate the plasmids from bacteria (plasmid curing) due to the smaller concentration (MIC) of the agent needed to inhibit plasmid replication as opposed to those required for the inhibition of replication of the bacterium harboring the plasmid [20,21].
Phenothiazines have been shown to inhibit the NorA efflux pump of S. aureus [20], the Qac efflux pump of the plasmid carried by a S. aureus multidrug resistant strain [6] and the AcrAB efflux pump of E. coli [15].
Phenothiazines affect the activity of genes that regulate and code the AcrAB efflux pump of E. coli [7,8,15] and Salmonella enterica serovar Enteritidis [8].
Because in our study the antimicrobial activity of the irradiated products of CPZ were many fold greater than that produced by unirradiated CPZ, and with respect to the Salmonella strains, had significant activity against the organism’s efflux pump, it would of course be interesting to determine whether the products of irradiation also affect the genes that regulate and code for the efflux pump of Salmonella as previously shown for the phenothiazine thioridazine [8].
The antimicrobial activity of the irradiated products of CPZ was identical against the S. aureus strains ATCC 25923 and HPV 107. However, because the irradiated products at ½ their MIC have no activity against the efflux pumps of these strains, the antimicrobial activity noted must be due to some other mechanism.
Due to the composition and structure of the cell envelope, Gramnegative bacteria have much higher intrinsic levels of resistance to various antibiotics, antiseptics, dyes, and detergents than Grampositive bacteria do [22]. Salmonella Enteritidis has at least nine multidrug efflux pumps [23]. One of these efflux pumps, AcrAB, is the most efficient, playing a role in both drug resistance and virulence [24]. Although for E. coli there are 36 drug transporters, belonging to the 4 major families of efflux transporters: MF, RND, SMR and ABC, only 21 of them can confer drug resistance either as single drug (e.g. drugspecific transporters such as TetA(B)) or as several unrelated drugs (e.g. multidrug transporters such as AcrB) [25]. For the ABC family only one ABC transporter confers drug resistance [26]. Because our study shows that the irradiated products have no activity against the efflux pump system of E. coli strains whereas they inhibit the efflux pump system of the Salmonella strains, it may be that the active products have activity against a non-AcrAB-tolC pump of the latter organism. If this is true, then this would be the first time that a compound(s) would have this type of efflux pump-selective activity. At this time the identity of the compound(s) that produce an inhibition of efflux by Salmonella is not known and is the subject of our current study.
The authors from NILPRP acknowledge financing of the research by Program LAPLAS 3 PN 09 39/2009, Alexandru T and Danko B were supported by STSM grants from COST Action BM0701 and A from COST action CM0804. Romanian National Authority for Scientific Research, CNCS – UEFISCDI by project number PN-II-ID-PCE-2011-3-0922 and , CNDI – UEFISCDI by project number PN-II-PT-PCCA-2011-3.1-1350. The authors acknowledge grants from the European Union co-funded by the European Social Fund (TAMOP- 4.2.2/B-10/1-2010-0012, and TAMOP-4.2.2A-11/1/KONV-2012-0035) A. Militaru and V. Nastasa were supported by projects POSDRU 107/1.5/S/80765 and POSDRU/88/1.5/S/56668, respectively. A. Martins and L. Amaral acknowledge the grants SFRH/BPD/81118/2011 and SFRH/BCC/51099/2010, respectively, provided by the Fundação para a Ciência e a Tecnologia, Portugal.