Cell & Developmental Biology
Open Access
ISSN: 2168-9296
ISSN: 2168-9296
Review Article - (2016) Volume 5, Issue 1
hTERT (human telomerase reverse transcriptase) is the catalytic subunit of telomerase enzyme, and is essential for its functions. The aim of this review was to compare the TERT in human and other species including microorganism, vertebrates and mammals, in terms of its functions and regulation. According to literature, the catalytic subunit of telomerase in animals contains many conserved domains and residues, which have crucial roles in its functions. Moreover, the structure and biology of human telomerase seem to be more similar to that of dog compared other animals. Thus interestingly, unlike the mouse that is seemingly not a proper model for evaluation of telomerase activity and its regulation, dog may be an appropriate model for the experimental investigations of telomerase function and therapeutic strategies in cancer studies.
<Keywords: Telomerase; hTERT; Human; Vertebrate; Yeast
Telomere consists of 6 nucleotide sequences (GGATTT) at the terminal region of the eukaryotic chromosomes. During chromosome replication, DNA polymerase cannot replicate the ends of chromosomes at somatic level, and gradual loss of telomeres occurs after each round of replication [1-3]. Somatic cells have a limited capacity for proliferation known as the Hayflick limit, which is due to this loss of chromosome ends, and the number of replication beyond this limitation leads to apoptosis. Telomerase is a ribonucleoprotein enzyme that is responsible for maintaining the chromosome integrity in proliferative cells, by providing specific replication machinery at the chromosome ends [4,5]. It has essential roles in cell immortalization, tumorigenesis and progression of most cancers, as it is shown that telomerase has a high activity in 70-90% of malignant human tissues and many immortalized cell lines [6-9].
Telomerase is a multi-subunit enzyme, composed of RNA and proteins including Telomerase reverse transcriptase enzyme (TERT), Telomerase RNA (TER), ES1P catalytic subunit ES3P, KU heterodimers, Dyskerin and TEP that are required for assembly and maturation of telomerase. In human, TERT is known as hTERT, which is catalytic subunit of telomerase and is characterized by having reverse transcription (RT) activities. Also, TER subunit in human is known as hTER, which acts as a template for reverse transcription [10-14].
Telomerase is tightly regulated in several levels, including transcriptional and post-transactional. On one side, alternative splicing has an essential role in the regulation of hTERT mRNA expression [13,15], and on the other side, studies have shown that telomerase function is regulated by post-translational regulation in serine/ threonine or tyrosine residue of hTERT [16]. In addition, several factors including p53, c-myc, AKT kinase, sp1, WT1, NF1, c-myb, AP1, and C-Ets2 regulate the catalytic subunits of telomerase [17-22].
It is demonstrated that human telomerase components are largely conserved in different species among mammals, and vertebrates, due to very important function of this enzyme [23-26]. Moreover, an association between regulatory factors in human telomerase and other species is stated [27,28]. Considering the numerous studies in this field, our aim in this review was to explore the relationship between human telomerase activity and functional hTERT with some candidate species. We compared function and sequences of hTERT with several species of mammals, vertebrates and microorganisms, in order to be able to suggest the best possible animal model for telomerase studies.
Yeast
Yeast telomerase is a ribonucleoproteins complex, composed of TERT (catalytic subunit), RNA template and protein subunits, however, its structure is not yet well known. In saccharomyces cerevisiae, there are a number of known subunits including Est1p, Est3p, Sm and Est2p (TERT) proteins in [29]. The comparison of TERT in yeast, compared to vertebrates, mammals and humans shows fundamental conserved residues in among species (Figure 1), which are essential for telomerase function. However, there are few regions in yeast telomerase which show specificity to this organism. The only less conserved in yeast can be considered as “nonconserved region” (N- region) of TERT, which seems to be species-specific characteristics. These regions are essential for maintaining telomerase and its performance, so the deletion based mutation in these regions affects the assembly of telomerase complex. On the other hand, deletion of the entire yeast TERT C-terminal does not interfere with the enzyme functions, but significantly reduces enzyme processivity.
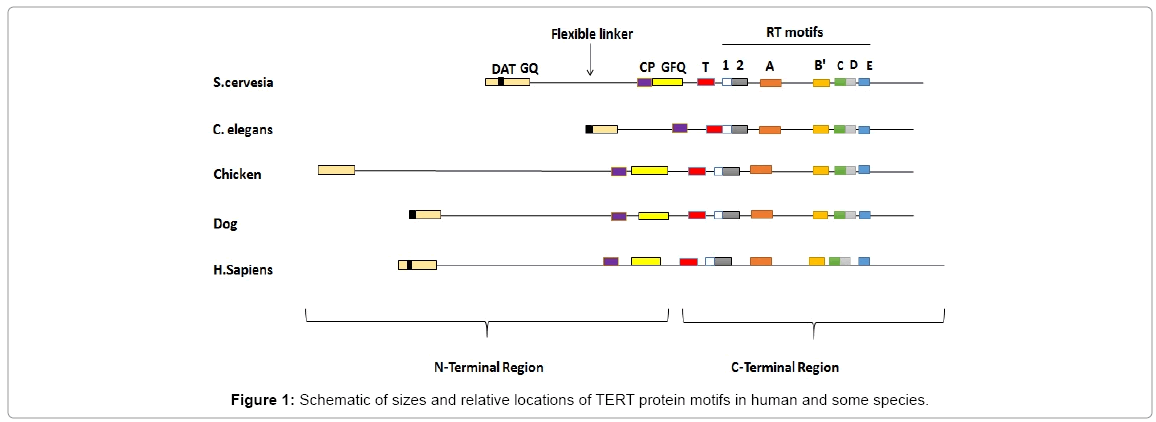
Figure 1: Schematic of sizes and relative locations of TERT protein motifs in human and some species.
In yeast, N-region proximal to GQ domains is an essential package for the telomerase processing and exhibiting the binding capability of nucleic acid. However, in human this package has important role in enzymatic processing. Furthermore, the domain of dissociation activities of telomerase is mapped to amino acid residues 69 to 134 in hTERT” and has a key role in elongation of telomere in vivo [27]. In addition, RT motif in yeast is highly conserved (includes 200-300 amino acids). Based on studies done in yeast and humans, a similar TERT and RT structure implies a similar function.
The main characteristic of yeast telomerase is an inability to synthesize long extension product. In yeast and human, the synthesis of 3’ telomeric DNA terminus is with the help of RNA template and short primers. DNA synthesis stops repeatedly during replication of template, therefore the majority of yeast telomerase products are inefficient, which suggests that yeast telomerase is inefficient not only at the 5’ end of template but also in all templates areas [27,29] (Figure 1).
C-terminal region is highly conserved in human and other species. Note the flexible linker of chicken is longer than that of the mammalian species. In addition structure of TERT between human and dog are more conserved compared to other species.
TERT in fishes
Among fishes, TERT is well-studied in the zebrafish and pufferfish.
Zebrafish (Danio rerio): The length of Telomere in zebrafish is similar to that of human. Zebrafish telomerase reverse transcriptase (zfTERT) cDNA is 126 KD, and the range of other cDNAs is between 103 to 133 KD. Amino acid sequence that is coded by zfTERT is 36% identical to human and 32% to mouse. In addition, zfTERT contains 7 RT motifs and T motif which is especially found in TERT but not in other reverse transcriptase. Unlike mammalians, the zebrafish has a very high telomerase activity in somatic tissues. For this reason, the organs of adult zebrafish grow throughout the life. Telomere lengths are different in several different organs and tissues in zebrafish lifelong, alternative lengthening of telomeres (ALT), mechanism may be involved in this case in order to maintain telomeres in the absence of telomerase [26].
In mouse and c.elegans, mutation in either of the telomerase components including TERT and TR does not affect the phenotype in the first generation, but after several generations defects and chromosomal instability appear. However, because telomeres of zebrafish and humans are shorter than mouse, defects that result from mutation in each of telomerase component appears in the first generation. The TERT-/- zebra fishes show their phenotypes after first generation and stop cell proliferation in many tissues, such as gut and testis, which is controlled by p53 [26].
Pufferfish (fugue): The N-terminal region in pufferfish, recognized as fTERT, contains three regions with functional capacity known as motifs GQ, CP and QFP. The motifs CP and QFP, are accompanied by T- motif locating at the center of TERT, which are capable to mediate, and distinguish the binding of telomerase RNA in human, yeast and tetrahymena. The fTERT promoter contains Transcription Factor Binding Sites which are also traceable in the hTERT promoter including E2F-1, c-Myc, Mad 1, estrogen. In addition, Sp1 and USF are notable as well; and transcription factors play roles in the regulation of hTERT. However, the common transcriptional role of TFBSs in the hTERT and fTERT promoters may reflect the sharing mechanisms in mammalian and fish as two diverse species. Surprisingly, it is reported that TERT has a “single canonical E-box” within the promoters which locates at “position _68 to _72, relative to the TSS” in variety of species including mouse (mTERT), chicken (cTERT) and pufferfish (fTERT) Besides, the Sp1 is capable to cooperate with c-Myc and activate the hTERT transcription. Imperatively, the hTERT promoter including its two response elements lead to bind estrogen to its receptor through which the transcription of hTERT would be enhanced. Furthermore, the Sp1 and estrogen binding have role in the fTERT promoter [30].
Comparison of fTERT between human, chicken, mouse, and hamster showed lack of TATA box in fTERT promoter .However, promoters of fTERT and cTERT, by having the CCAAT box may characterize “TATA-less promoters” with an initiating role. Although the fTERT promoter has no CpG islands, but the human-TERT promoters and chicken seem to be GC-rich [31].
TERT in birds
Chicken: Among birds, TERT is well-studied in the chicken. The chicken telomerase reverse transcriptase (chTERT) proteins have 45% similarity to humans. The chTERT property can be distinguished from hTERT and other vertebrates by larger sizes of chTERT proteins due to greater linker region in N-terminal domain (144 amino acid greater than human). The chTERT genes are located at 2q2, near the telomere. Amongst many transcription factors, those binding to 5’ UTR or promoter of chTERT are similar to human (Table 1). In spite of this fact, there are binding site of p53, WT1 and c-Ets2 in hTERT promoter that not present in chTERT promoters. Exclusively there are many binding site for c-myb in 5’ UTR of chTERT. However, there is only one E-box in -264 of chTERT promoters but two E-box in -34 and -242 of hTERT [24].
| Factors Species |
P53 | c-myc | AP1 | NF1 | WT1 | C-Ets2 | SP1 | c-myb |
|---|---|---|---|---|---|---|---|---|
| Human | Rep | Act | Act | Act | Rep | Act | Act | Act |
| Dog | Rep | Act | Act | Act | Rep | Act | Act | Act |
| Mouse | Rep | No effect | ||||||
| Chicken | _ | Act | Act | _ | _ | Act | Act | |
| Zebrafish | Rep | |||||||
| Pufferfish | - | - | Act | Act | - | - | Act | Act |
Table 1: The comparison of factors binding to hTERT promoters in different species.
Moreover there is CCAAT motifs in chTERT but is not found in hTERT promoter. This motif is essential for transcription initiation of TATA poor promoters, which usually is adjacent to transcription start site. Both hTERT and chTERT have many CpG Island in 5’ UTR and coding region, this shows that their function is regulated by methylation mechanism [23,24,28]. It could be concluded that regulation of chTERT and hTERT expression is similar but they have apparently different structures (Table 1).
TERT in mammalians
Telomerase structure and biology has been studied in several mammalian species include pig and dog.
Pig: Telomere sequences in pig is revealed to be TTAGGG which is similar to human, although its length in pigs is 10-30 KD which is longer than humans’. Lymph node, lung, and kidney tissues of male and female have high telomerase activities, but male pigs unlike females display no or very low levels of telomerase activity in the liver. In contrast, the telomerase activity of spleen in female pigs is higher than males. Moreover telomerase activity is low in ovary of female pigs. In contrast the humans, telomerase activity in ovary is associated to proliferation of endometrial tissue during the menstrual. The pig telomerase biology is more similar to mouse and rat rather than other animals and human [32].
Dog: The dog TERT protein composed of 1123 amino acids with a molecular weight of -124 kDa. Dog telomere length is 12-23 kb. Comparing the dog TERT with other mammals, it seems that the dog TERT have highest level of similarities to human TERT. Besides, telomerase activity in canine cell line is similar to human.
TERT in many eukaryotes such as yeast, ciliates, plants and mammals, is a large catalytic protein domain, which composed of several RT motifs, including 1, 2, A, B, C, D, and E which are essential for catalytic activity. TERT specific motif that is called T-motif, is located at the center of dog TERT proteins. The comparison of canine TERT with vertebrates revealed that the sequences of TERT N-terminal in this species are very similar [33,34]. The hTERT are more closely resembles to dog TERT rather than other mammals. Only Minor differences exist between human and dog sequences and we tried to refer to some of them; the core promoter of hTERT located at 300 bp upstream of start codon (ATG), but in dog TERT core promoter is located 314 bp away from start codon. Besides, there are several Sp1 binding sites located within this region and there is one MZF2 (zinc finger 2 protein) binding site in-605 of canine TERT promoter. MZF2 inhibits the expression of TERT promoters, and several MZF2 binding site exist in the hTERT promoter, four of which located at 200 bp upstream of hTERT core promoter.
In canine TERT there is an E-box at -1397; whilst, two E-boxes present in hTERT promoter. WT1 binding site located in the core promoter region of canine hTERT but in -358 bp of hTERT promoter. Telomerase levels are undetectable or low in both canine and human somatic tissues, and telomere gets shorter through each division. At a glance, differences and similarities between telomerase expressions in various tissues of humans are compared with several species in Table 2. Considering these facts that the dog physiology has many similarities with human [25,35,36] and that telomerase is highly expressed in most canine cancers (up to 90%), it seems that regulation of the dog telomerase catalytic subunits is very similar to humans (Table 2).
| Tissue Species |
Ovary | Testis | Respiratory system | Lymphocyte | Heart | Kidney | Spleen | Liver | Gastro enteral system |
|---|---|---|---|---|---|---|---|---|---|
| Human | High | High | - | Low | - | - | Low | - | Low |
| Mouse | - | High | Moderate | Moderate | - | - | Moderate | High | High |
| Zebrafish | High | High | Very high | Low | High | ||||
| Pig | Low | High | Very high | Very high | Very high | ||||
| Dog | High | Low | Moderate | Low | Moderate | - | |||
| Cat | High | Low | Moderate | Low | Moderate | ||||
| Chicken | High | High | low | Very high | High | High | |||
| Pufferfish | High | High | High | moderate | moderate | moderate |
Table 2: Comparison of telomerase expression in human, mammalians and vertebrates tissues.
Mouse: The biology of telomerase in humans and mice are very different. Human telomeres are much shorter than that of mice. Telomere shortening has a pivotal role in tumor growth in humans but has limited role in mouse. Most mouse somatic cells, unlike human, gain telomerase activity and express hTERT [25,37]. Transcriptional activity of the hTERT promoter is much less than the TERT promoter in mouse that may be due to no conserved CG-box regions in hTERT promoter which are responsible for this suppression. So cis-regulation has been suggested to be important for regulation of the TERT transcription in tissue-specific manner [38]. By considering these facts, regulation of telomerase function in mouse and humans are highly different. However, mouse TERT (mTERT) promoter like hTERT has two E-boxes and also has p53 binding site. Mutations in E-box reinforce both hTERT and mTERT promoter expression; in fact, the hTERT promoter is activated by c-myc and deactivated by p53 while the mTERT promoter does not respond to either of them [39].
Studies on human ovarian epithelial cells, vascular endothelial cells, and breast cancer cells suggested that an Estrogen receptor Responsive Element (ERE) exists in the promoter region of hTERT, but there is not in the mouse TERT promoter [40]. In another study on MEF and HEK cell lines, it was determined that the hTERT promoter was inhibited by p53, p63 and p73. Although p53 inhibit the hTERT expression through inhibition of c-myc, in mouse cells inhibit the TERT expression via Ebox/E2F pathway [41]. There are 12 CpG Island at 615bp upstream of the transcription start site in mTERT promoter; these regions are hypomethylated (30% in adult liver, 20.8% in fertilized egg). This could be a potential explanation of higher telomerase activity in mice compared to human. Thus, methylation plays significant role in regulation of mTERT expression [42,43]. Finally as an overview, the key characteristics of TERT in different species are provided which is reflective of common behavior and diversity as well (Table 3).
| Findings | References | Findings | References |
|---|---|---|---|
| Human | Dog | ||
| TGF-beta treatment of MCF-7 cells_ repressed the hTERT promoter activity in a dose-dependent manner. | Yang et al. [1] | Molecular targets, telomerase biology and tumor geneticsin dogs, cats and humans. | Pang et al. [33] |
| hTERT promoter is GC-rich, lacks TATA and CAAT boxes_ contains binding sites for several transcription factors | TERT mRNA expression is associated with telomerase activity in dogs | Nasir et al. [34] | |
| hTERT expression subject to multiple levels of control_ regulated by different factors | Multiple transcription factor binding sites to the canine TERT promoters_E-box, Sp1, AP1, MZF-2 and ER/Sp1 binding sites_ similar to hTERT | Long et al. [25] | |
| The hTERT expression is partly regulated by atypical alternative splicing | Wong et al.[15] | Dog telomere biology is similar to that in humans | Nasir et al. [36] |
| Yeast | |||
| Re-amplification of the products could help the investigators to determine the majority of telomerase-positive samples. | Mehdipour[16] | Utility of budding to identify a large multi-subunit RNP enzyme (383 kDa, including hTERT-Cdc13 and hTR. | Wong et al. [29] |
| Regulatory regions of the hTERT promoter _positive and negative regulation of telomerase | Horikawa et al.[19] | Structure of the core telomerase components_RNAand telomerase reverse transcriptase subunit | Neal et al. [27] |
| Zebrafish | |||
| Telomerase repressed in normal human somatic tissues but reactivated in cancer | Kim NW et al.[6] | Zebrafish is effective model for the discovery of new drugs able to reactivate telomerase in dyskeratosis congenital DC patients. | Anchelin M et al.[26] |
| Mutations in sporadic melanoma cases at two positions in the TERT promoter _generated binding motifs for Ets/TCF transcription factors. | Horn S et al. [13] | ||
| Chicken | |||
| The hTERT protein plays a key role in the activation of telomerase in cancer cells | Nakayama J et al. [11] | Several transcription factor binding motifs in the 5' flanking/promoter region of chTERT _similar to hTERT (E-box, Ik1, MAZ, Sp1 sites)_several c-Myb sites in chTERT only | Delany ME et al. [24] |
| Shelterin protein interacts with hTERT and recruits hTERT onto the telomeres_ novel function of hTERT adds a new element to the molecular model of telomere length maintenance. | Zheng YL et al. [3] | Telomerase activity in chickens similar to mice. | Venkatesan RNet al. [3] |
| Association between hTR expression and telomerase activity | Hosseini-Asl S et al. [44] | TERT and TR transcript levels correlate with telomerase activity_TR is the rate-limiting factor in telomerase-negative tissues. | O'Hare TH et al. [28] |
| Mouse | |||
| Expression of hTR and hTERT may be independent of tumors’ stage. | Hosseini-Asl S et al. [44] | Essential role for telomerase and telomeres, in the maintenance of genomic integrity and viability of high-renewal organ systems. | Lee HW et al. [37] |
| Several enzymes are necessary for telomerase functioning that facilitate new approaches for inhibit the telomerase in treating cancer. | Holysz H et al. [5] | The condensed chromatin environment of hTERT locus was crucial to its silencing during cell differentiation. | Wang S et al. [38] |
| Progression of malignancy is dependent on activation of telomerase | Counter CM et al. [7] | large body mass presents an increased telomerase activity and cancer risk | Gorbunova V et al. [45] |
| Pufferfish | |||
| Activation of telomerase is an event that starts mostly at low grades of brain including meningioma and astrocytoma tumors. | Kheirollahi M [9] | The level offTERT expression was found to be higher in actively dividing cells and reduced at inactivity cell cycle regulate TERT and possibly telomerase activity. | Yap WH et al. [30] |
| Pig | |||
| Importance of using hTERT genes FISH probes for cases with cancer cervix | Eid MM et al. [22] | Somatic pig tissues levels of telomerase activity more similar to mouse and contrasts with humans and dog. | Fradiani PA et al. [32] |
Table 3: Exploration of Key facts in TERT territory in different species at a glance.
It worth’s to highlight the facts of similarity in specific physiologic and anatomic pattern for vertebrates regarding the selected biological characteristics (Table 4).
| Characteristics | Key similarities to humans | Key differences | ||
|---|---|---|---|---|
| Zebrafish and Pufferfish | ||||
| Lifespan | Lifespan of 3-5 years; generation time of 3 months | |||
| Anatomy | Vertebrate body plan | Fishes; Aquatic adaptations include streamlined body plan and different locomotor strategies | ||
| Diet and metabolism | Omnivorous | Poikilothermic, grows optimally at 28.5°C | ||
| Reproductive system | Molecular and embryological biology of germ-cell development; cellular anatomy of germ-cell organs, the testis and ovary | No sex chromosomes; fertilization is ex vivo (that is, no uterus or the related internal female reproductive organs); oocytes are surrounded by a chorion, not the zona pellucida, which must be penetrated by sperm; non-lactating; no breast equivalent | ||
| Gastrointestinal system | Major organs: liver, exocrine and endocrine pancreas, gall bladder; zonal specializations along the length of the absorptive alimentary tract; immune cells in lamina propria | Lack an acidified digestive organ; have an intestinal bulb rather than stomach; intestinal Paneth cell not present | ||
| Respiratory system | Cellular gas exchange; oxygenation is dependent on circulation and hemoglobin carriage | Respiration occurs in gills, not lungs; no pulmonary circulation; possess an endoderm-derived swim bladder (functioning as a variable buoyancy device), which corresponds embryologically but not functionally to the lungs | ||
| Cardiovascular system | Multi-chamber heart with an atrium and ventricle; circulation within arteries and veins; separate lymphatic circulation | Has left–right distinctions in cardiac anatomy, but does not have separate left–right circulations, that is, the heart has only two chambers; so far no evidence for secondary heart field derivatives; lymph nodes have not been described | ||
| Chicken | ||||
| Lifespan | Life span of 8 year: generation time of 6 months | |||
| Anatomy | Vertebrate body plan | Bird; light skeletal system and light but powerful musculature; have more cervical (neck) vertebrae; | ||
| Diet and metabolism | Omnivorous; homeothermic | The average body temperature of a chicken is 41-45°C; | ||
| Reproductive system | Males have two testes | Females have two ovaries; right testes are also smaller than the left; egg-laying; have no phallus; sperm is stored in the seminal glomera. | ||
| Gastrointestinal system | Small intestine; chicken has crop; gizzard | |||
| Respiratory system | Have lungs | Do not have a diaphragm; the gas exchange occurs in the walls of microscopic tubules, called 'air capillaries; transferring more oxygen with each breath.' | ||
| Cardiovascular system | Four chamber heart, a right atrium and ventricle which receive deoxygenated blood from the body and send it to the lungs and a left atrium and ventricle which receive oxygenated blood from the lungs and send it to the body. | Has to work much harder than a human heart, the inside walls of the atria and ventricles are much smoother than those of the human. And the valves, though present, are much simpler. The ventricles of the bird heart have more muscle mass and less chamber space than those of a human; faster heart rate | ||
| Mouse | ||||
| Lifespan | Life span of 2-3 year: generation time of 8-9 week | |||
| Anatomy | Vertebrate body plan; mammals | Size; skull and tail morphology and in the shape and orientation of shoulder and pelvic girdle bones; Articular cartilage is thinner in mice and the subchondral bone plate more variable in thickness; cortex mostly comprises circumferential lamellae; structure of the dentition | ||
| Diet and metabolism | Omnivorous; the average body temperature is 36.9°C; homeothermic | Gnawer | ||
| Reproductive system | Similar ovaries, fallopian tubes, uteri, and placentation; histologic appearance and basic functions; | There are several structural and functional differences; prostate is divided into four lobes, where each lobe is histologically distinct; contains additional accessory sex glands that are not found in the human | ||
| Gastrointestinal system | Abundant bacterial flora within the colon; smooth muscle–enveloped tube with innermost mucosa (barrier epithelium, lamina propria, and muscularis mucosae), submucosa, muscularis propria, and variable serosa or adventitia | The morphologic appearance of the different sections of the upper digestive tract | ||
| Respiratory system | Both have two lungs; diaphragm is present; trachea has cartilage rings. | Anatomy and histology of the lungs; higher basal metabolic rate and more rapid respiratory rate in mice. The relative size of airway lumens of mice is larger; Mouse has smaller lungs, and branching occurs after bronchiole levels; Fungus seen in mouse lung | ||
| Cardiovascular system | Four-chambered hearts: left and right atria, a dominant left ventricle, and a thinner-walled right ventricle; the ratio of heart to body weight | Mouse heart is far smaller than the human heart; thinner walls of mouse arteries and the prominent presence of cardiomyocytes around mouse pulmonary veins | ||
| Pig | ||||
| Lifespan | Life span of 8 year: generation time of 12 month | |||
| Anatomy | Vertebrate body plan, mammals; major chest and abdominal muscles found in humans are present in the pig; dental anatomy | Different facial expression; bigger ears, a tail; different feet; position and number of toes on their feet; location of chest muscles that attach to the shoulder girdle; gluteal muscles | ||
| Diet and metabolism | Omnivorous; homeothermic | Average body temperature 38.8°C | ||
| Reproductive system | Ovaries; testes; prostate gland | In female urogenital sinus is present; oviducts each leads to horn of the uterus; Uterus two horn. In male penis underneath the ventral skin surface, posterior to the umbilical cord; |
||
| Gastrointestinal system | Esophagus; stomach, small and large intestines. Mesenteries | Colon is spiral | ||
| Respiratory system | Multi-lobed lungs; bronchial tubes leading to the lungs | Lungs have thinner walls; alveoli are less densely packed; pigs don't have a diaphragm. Instead, they use muscles in their throat sacs to help draw in air and push it back into the lungs; they don't breathe through their mouths; use muscles beneath their jaws to help move air | ||
| Cardiovascular system | Size; the heart is in the same place between the lungs, the liver and gall bladder | Aortic-mitral fibrous continuity was reduced in the outlet component of the porcine left ventricle, with approximately two-thirds of the aortic valve being supported by left ventricular musculature; the arterial duct (ductus arteriosus) leads directly from the pulmonary trunk to the aorta. | ||
| Dog | ||||
| Lifespan | Life span of 13 year: generation time of 3 year | |||
| Anatomy | Vertebrate body plan, mammals; Dog anatomy includes the same internal structures that are in humans; brain have dedicated voice areas; brains, are also sensitive to acoustic cues of emotion; | have disconnected shoulder bones (lacking the collar bone of the human skeleton); The olfactory bulb in dogs is roughly forty times bigger than the olfactory bulb in humans, The teeth are designed to crush dense objects; | ||
| Diet and metabolism | The average body temperature is 37-39°C; homeothermic | Carnivorous | ||
| Reproductive system | In the female, the reproductive system is composed of the ovaries, oviducts, uterus, cervix, and vagina; the ovaries are the site of production of the unfertilized eggs; the eggs pass from the ovaries into the oviducts; The developing embryos mature within the uterus, attached to its walls by the placenta which also surrounds them; uterus of the dog is Y-shaped. In the male Testicles, ductus or vas deferens, prostate gland, and penis. Sperm production and storage occurs within the testicles. | The epididymis is comparatively large; | ||
| Gastrointestinal system | Physiology of stomach; small intestine; large intestine; esophagus trace | Stomach is more acidic; Digestive tract is a lot smaller | ||
| Respiratory system | Have right and left lungs; diaphragm muscle; trachea has cartilage rings; the body's carbon dioxide is replaced with oxygen | It is also a unique cooling system | ||
| Cardiovascular system | heart has 4 chambers; The chambers on the right side are completely separate from the chambers on the left side; two arteria and two ventricle; a dominant left ventricle, and a thinner-walled right ventricle ; coronary arteries; coronary sinus remains similar in size to humans | Smooth inner lining of the ventricle; dog heart smaller than human hearts. | ||
Table 4: Similarities and difference facts between human and some vertebrate animals.
In spite of the similarities and the differentiae facts amongst these species, the main leading conclusion relies on’ development and evolution’. However, our aim was to achieve the applicability of one of the mentioned animals as experimental tool in human in purpose of modeling the telomerase biological pattern and drug innovation.
As a complementary insight, some regions of N-terminus and C-terminal of TERT that have essential roles in telomerase activity are conserved in human and other species, while some other regions in N-terminus of hTERT, that have critical roles in telomere maintenance and the ability of telomerase process, are not conserved.
Although the structure of the yeast TERT gene is very similar to hTERT, its performance is different due to the extra regulatory proteins in human telomerase. On the other side, the regulation of expression in chicken TERT and hTERT are similar but their structure is different. The hTERT and mouse TERT sequences are similar but remarkable differences in regulation of their functions may be due to the extra proteins in human telomerase structures and its’ different transregulation. Notably, the structure of the catalytic subunit, the regulation of expression and telomerase function in dog is more similar to humans compared to other animals.
According to previously conducted studies (Table 3), telomere length and telomerase function is associated to body size. Since dogs have a size larger than other studied animals, and also that human and dog have lots in common regarding anatomy, metabolism, alimentary tract, reproductive-e, respiratory- and cardiovascular system (Table 4), it is probably why the structure and biology of human telomerase is more similar to dog than other animals (Figures S1 and S2). Tumor development mechanism and also similar cancer stem cells are conserved between humans and dogs, as well (Table 3). Due to the biological similarities, we can predict similar response to treatment in cancers. These results suggest that dog can be an excellent model for exploring telomerase and diseases related to its abnormal activity.