Advanced Techniques in Biology & Medicine
Open Access
ISSN: 2379-1764
ISSN: 2379-1764
Review Article - (2013) Volume 1, Issue 2
The proximate composition of four egg stages and 1st zoeal stage of Macrobrachium idella idella were investigated. The protein content of egg was gradually decreased from egg stage-1 (69.32%) to egg stage-IV (54.56%) and the protein content in Ist zoeal stage was found to be 52.42%. The carbohydrate content increased from egg stage-I (3.24%) to freshly hatched Ist zoea. Lipid values showed a decreasing trend from egg stage-I (15.86%) to freshly hatched Ist zoea (4.82%). The water content uniformly increased from egg stage I (62.26%) to freshly hatched Ist zoea stage (79.68%). Among total 9 individual saturated fatty acids reported, the palmitic acid (C16:0) was maximum in all egg stages and in Ist zoea. Which is followed by stearic (C18:0) and myristic (C14:0) acids. The total essential amino acids were showed increasing trend from the egg stage I to IV. However they were little bite low in I zoea stage when compared to egg stage IV. As in essential amino acids, non essential amino acids were also followed almost similar trend. Among total 9 amino acids, 5 amino acids (glutamine, asparagine, glycine, cystine and alanine) were gradually increased from egg stage I to IV.
Keywords: RNA interference, Gene silencing, Recombination
The females of most decapods extrude their eggs and carry them under the abdomen until hatching. The eggs are rich in yolk substances that are used as embryonic development progresses [1-5]. The utilization of yolk substances closely correlates with the embryonic development. Protein is one of the main components of yolk, plays an important role in both morphogenesis and energy supply in embryos [2,3]. Lipid in the eggs of the most of the crustacean is one of the main energy sources that the females stored. During the embryonic development stages, lipid is not only a kind of energy source, but also components of biological membranes and pigments of compound eyes [4,5]. There is no information available on the biochemical composition during embryonic development and freshly hatched zoea of crustaceans in general and palaemonid prawn, Macrobrachium idella idella in particular. So the present study is designed to know the biochemical changes in the embryonic and freshed hatched first zoeal stages of edible prawn, Macrobrachium idella idella.
Collection
The prawn, Macrobrachium idella idella was collected from Ponanthittu (Lat.11°28`50.50º N; Long. 79°45`28.23º E) waters which is located 2 km south to Parangipettai and connected with Vellar estuary. The prawns were caught by the fisherman with the help of trap, line and hand- net and scoop net. The specimens were collected from fisherman as the sizes of the collected specimens were ranged from 30 to 125 mm in length. Totally 210 specimens were collected and transported to the laboratory in live condition by keeping them in bucket containing freshwater and aeration. After reaching the laboratory they were washed carefully with distilled water to remove dust and algal particles and ice killed.
The freshly collected berried females during the months of November and December 2006 were segregated into four arbitrary stages as described by Aubson and Patlan, Sumitra and Easterson, Rodriguez (1977) and Rodriguez (1985), respectively [6-9]. The color and diameter of the eggs were noted before segregation and small clumps of eggs was snipped from random locations in each clutch using sharp scissors. All the developing embryos were examined with a MEIJI binocular dissecting microscope (100X) to ensure that only viable embryos were sampled and the color of the embryo were also observed. The diameter of the eggs was measured using a micrometer mounted in the ocular of a dissecting microscope. Eggs were classified into 4 stages based on the following characteristic features.
Characteristic features of different egg stages
Egg stage-I: The eggs were opaque, greenish, round or oval in shape. The diameter of the egg was found to be 0.45 mm.
Egg stage – II: The eggs were translucent, light green in color, oval with a narrow peri-vitelline space at one end and small transparent plate (the blastoderm) were easily distinguished. The diameter of the egg was calculated as 0.57 mm.
Egg stage-III: The eggs were translucent, brownish-yellow in color. The embryo further developed with anterior transparent plate and differentiated into cephalic lobe. The diameter of the egg was measured as 0.58 mm.
Egg stage – IV: The eggs were transparent, dull whitish in color. The embryos were well-developed with conspicuous black eye spots at the anterior and rudiments of cephalothoracic appendages were developed, the posterior region was Coiled, and the diameter of the egg was found to be 0.65 mm.
The eggs were blot dried with filter paper and transferred to the previously weighed dry paper. The dry weight of the eggs was determined after drying them in a hot-air oven at 60ºC for 24 hours. Then the samples were stored in desiccators with calcium chloride until analyzed.
Analysis of proximate composition
The protein, carbohydrate and lipid contents were estimated by adopting the standard methods of Raymont et al., Dubois et al. and Folch et al. [10-12], respectively.
Moisture
The body tissue as a whole was dried in an electric oven at 60°C until a constant weight was reached and then it was weighed in an electronic balance to the nearest 0.1mg. The percentage of dry matter in the body was calculated. The differences in weight between wet and dried tissues represented the weight of water in the body tissue, which was expressed as percentage.
Estimation of Fatty acids
The fatty and methyl esters of the sample was injected into the gas chromatography (GC-6890) capillary column coated with 5% phenyl silicone at a temperature from 170º to 300ºC for 23.33 minutes. Flame ionization detector was used for analysis. Based on the retention time, the different fatty acids in the samples were identified. Triplicate was maintained for each experiment.
Estimation of amino acids
The amino acids were determined by an automatic amino acid in HPLC analyzer (Lachromehitachi). Five microliter of amino acids standards mixture sample were injected into the column (DENALIC18 5MICROMM 4.6 mm×150 mm). The flow rate was about 1 ml per minute, ambient temperature of 23°C was maintained and sample was detected at 254nm by following the method of Baker and Han [13].
Statistical analysis
The data were subjected to One- way analysis of variance (ANOVA) and difference between means were determined by Duncan’s multiple range tests (P<0.05) using SPSS version 17.0.
The proximate composition of four egg stages and 1st zoeal stage of Macrobrachium idella idella are presented in Table 1.
| S. No | Constituents | I-Egg stage | II- Egg stage | III-Egg stage | IV-Egg stage | Ist Zoeal stage |
|---|---|---|---|---|---|---|
| 1 | Protein | 69.32 ± 0.40a | 64.88 ± 0.19b | 59.82 ± .18c | 54.56 ± .20d | 52.42 ± 0.45e |
| 2 | Carbohydrate | 3.24 ± 0.23c | 3.56 ± 0.05b | 3.65 ± 0.11b | 3.86 ± 0.14b | 4.16 ± 0.19a |
| 3 | Lipid | 15.86 ± 0.29a | 12.45 ± 0.12b | 8.08 ± 0.11c | 5.96 ± 0.25d | 4.82 ± 0.13e |
| 4 | Moisture | 62.26 ± 0 .13e | 64.42 ± 0.13d | 72.42 ± 0.21c | 78.52 ± 0.27b | 79.68 ± 0.19a |
Different superscript showed in a row significantly different (P<0.05).
Table 1: The proximate composition (%) of M. idella idella during different embryonic and Ist zoea stage. (Values expressed in %, dry weight except moisture. Each value is the mean of 3 sample ± SD).
Protein
The protein content of egg was gradually decreased from egg stage-1 (69.32%) to egg stage-IV (54.56%) and the protein content in Ist zoeal stage was found to be 52.42%. The differences in the protein content of different eggs stages and Ist zoea are statistically significant (Figure 1).
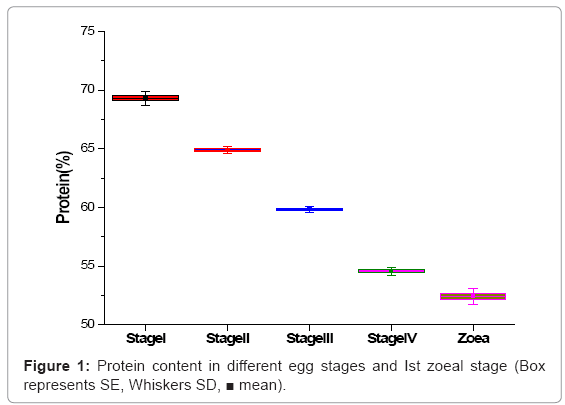
Figure 1: Protein content in different egg stages and Ist zoeal stage (Box represents SE, Whiskers SD, ■ mean).
Carbohydrate
The carbohydrate content increased from egg stage-I (3.24%) to freshly hatched Ist zoea. The increasing trend of the carbohydrate from egg stage I to freshly hatched Ist zoea stage was statistically significant. But among the egg stages from II to IV the carbohydrate content was not statistically significant (Figure 2).
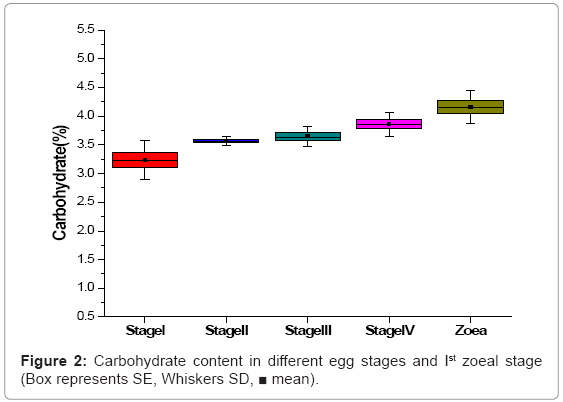
Figure 2: Carbohydrate content in different egg stages and Ist zoeal stage (Box represents SE, Whiskers SD, ■ mean).
Lipid
Lipid values showed a decreasing trend from egg stage-I (15.86%) to freshly hatched Ist zoea (4.82%). This decrement of lipid content was statistically significant (Figure 3).
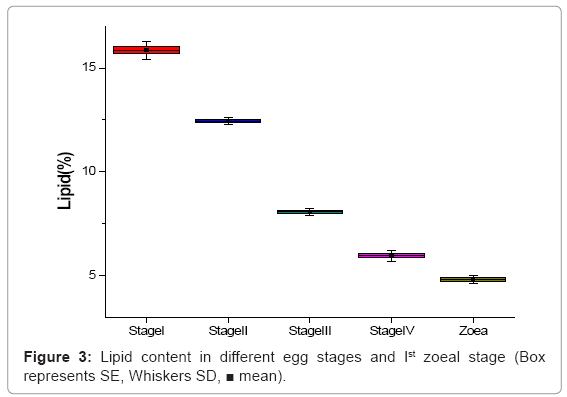
Figure 3: Lipid content in different egg stages and Ist zoeal stage (Box represents SE, Whiskers SD, ■ mean).
Moisture
The water content uniformly increased from egg stage I (62.26%) to freshly hatched Ist zoea stage (79.68%) and this was statistically significant (Figure 4).
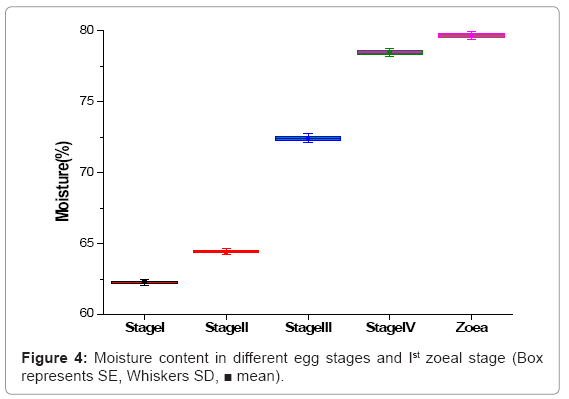
Figure 4: Moisture content in different egg stages and Ist zoeal stage (Box represents SE, Whiskers SD, ■ mean).
Fatty acids
Among total 9 individual saturated fatty acids reported, the palmitic acid (C16:0) was maximum in all egg stages and in 1st zoea. The palmitic acid was followed by stearic (C18:0) and myristic (C14:0) acids. But myristic (C14:0) acid in egg stage III was comparatively less than other egg stages and zoeal stage. The tridecyclic (C13:0) acid was not detectable in stage III and nonadecyclic (C19:0) was not detectable in stage IV and in 1st zoeal stage (Table 2). The total saturated fatty acids decreased from egg stage I to Ist zoea. But the total saturated fatty acids between egg stages II to III was not statistical different.
| Fatty acids (%) | Egg stage-I | Egg stage-II | Egg stage-III | Egg stage-IV | 1st zoea stage |
|---|---|---|---|---|---|
| C12:0 | 1.30 ± 0.26a | 1.22 ± 0.03a | 0.45 ± 0.13c | 0.56 ± 0.14bc | 0.83 ± 0.15b |
| C13:0 | 0.47 ± 0.12a | 0.23 ± 0.07ab | ND | 0.17 ± 0.05ab | 0.25 ± 0.08ab |
| C14:0 | 4.08 ± 0.27a | 3.87 ± 0.24ab | 1.38 ± 0.41d | 3.58 ± 1.37b | 3.02 ± 1.26c |
| C15:0 | 1.04 ± 0.14abc | 1.28 ± 0.33ab | 1.30 ± 0.21a | 0.94 ± 0.14bc | 0.86 ± 0.17c |
| C16:0 | 25.33 ± 0.21a | 24.46 ± 0.79b | 24.14 ± 0.09bc | 23.97 ± 0.63c | 21.19 ± 0.37d |
| C17:0 | 1.92 ± 0.33a | 1.75 ± 0.24ab | 1.99 ± 0.33a | 1.42 ± 0.23b | 1.03 ± 0.08c |
| C18:0 | 6.58 ± 1.72b | 5.51 ± 1.53c | 7.84 ± 0.83a | 4.97 ± 0.78d | 4.88 ± 0.20d |
| C19:0 | 0.38 ± 0.19a | 0.20 ± 0.04ab | 0.37 ± 0.11a | ND | ND |
| C20:0 | 0.37 ± 0.14a | 0.46 ± 0.16a | 0.53 ± 0.08a | 0.66 ± 0.10a | 0.54 ± 0.08a |
| Total | 41.47 ± 1.63a | 38.98 ± 2.09b | 38.00 ± 1.39b | 36.27 ± 0.80c | 32.6 ± 0.67d |
Different superscript showed in a row significantly different (P<0.05).
ND: Not Detected
Table 2: Saturated fatty acids (Each value is the mean of 3 sample ± SD) of developing eggs and 1st zoeal stage of M. idella idella.
The monounsaturated fatty acid C18:1n9c (Oleic) was maximum followed by C18:1n7c. The C14:1n5c was not detectable in egg stages II and IV and also C16:1n9c was not detectable in egg stage II. The C17:1n6c was not detectable in egg stages III and IV and C18:1n5c was not detectable in egg stage II. The fatty acid C19:1n9c was not detectable in stage IV and Ist zoea stage (Table 3). The total amount of monounsaturated fatty acids gradually decreased from egg stage I to Ist zoea and the decrement was statistically significant.
| Fatty acids | Egg stage-I | Egg stage-II | Egg stage-III | Egg stage-IV | 1st zoea stage |
|---|---|---|---|---|---|
| C14:1n5c | 0.16 ± 0.03a | ND | 0.12 ± 0.02a | ND | 0.15 ± 0.02a |
| C16:1n5c | 0.58 ± 0.08a | 0.54 ± 0.07a | 0.45 ± 0.06a | 0.21 ± 0.05a | 0.60 ± 0.1a |
| C16:1n9c | 0.43 ± 0.07b | ND | 0.35 ± 0.07bc | 0.45 ± 0.08b | 1.31 ± 0.20a |
| C17:1n8c | 0.98 ± 0.14b | 1.17 ± 0.16ab | 1.37 ± 0.18a | 0.91 ± 0.13b | 1.47 ± 0.12a |
| C17:1n6c | 0.32 ± 0.05a | 0.22 ± 0.04a | ND | ND | 0.27 ± 0.02a |
| C18:1n9c | 25.04 ± 0.83a | 19.77 ± 0.70c | 25.34 ± 0.87a | 22.40 ± 0.95b | 18.65 ± 1.94d |
| C18:1n7c | 6.23 ± 1.20b | 5.40 ± 1.5c | 8.17 ± 1.22a | 3.44 ± 0.14d | 3.33 ± 0.11d |
| C18:1n5c | 0.42 ± 0.05a | ND | 0.26 ± 0.05ab | 0.30 ± 0.02ab | 0.33 ± 0.08ab |
| C19:1n9c | 0.22 ± 0.02a | 0.15 ± 0.03a | 0.31 ± 0.02a | ND | ND |
| Total | 34.38 ± 1.49b | 27.25 ± 1.43d | 36.37 ± 1.49a | 27.71 ± 1.38c | 26.11 ± 1.60e |
Different superscript showed in a row significantly different (P<0.05).
ND: Not Detected
Table 3: Monounsaturated fatty acids (Each values is the mean of three values ± SD) of developing eggs and 1st zoeal stage of M. idella idella.
The total amount of polyunsaturated fatty acids was irregular in eggs as well as zoeal stage. Among 5 polyunsaturated fatty acids, C18:2n6c was found to be maximum followed by C20:3n6c and C20: 4n6c (Table 4). The polyunsaturated fatty acids were showed significantly variation among egg stages I to 1st zoea.
| Fatty acids | Egg stage-I | Egg stage-II | Egg stage-III | Egg stage-IV | 1st zoea stage |
|---|---|---|---|---|---|
| C18:2n6c | 6.34 ± 0.34b | 5.67 ± 0.25cd | 5.88 ± 0.19c | 7.12 ± 0.29a | 5.46 ± 0.31c |
| C18:3n6c | 0.54 ± 0.06a | 0.48 ± 0.08a | 0.47 ± 0.04a | 0.78 ± 0.07a | 0.43 ± 0.06a |
| C20:2n6c | 0.62 ± 0.06b | 0.74 ± 0.08ab | 0.97 ± 0.09ab | 1.12 ± 0.11a | 0.94 ± 0.09ab |
| C20:3n6c | 1.45 ± 0.10ab | 1.62 ± 0.05a | 1.74 ± 0.07a | 1.16 ± 0.13b | 0.57 ± 0.04c |
| C20:4n6c | 0.86 ± 0.07b | 0.69 ± 0.07b | 1.97 ± 0.08a | 1.73 ± 0.05a | 2.00 ± 0.08a |
| Total | 9.81 ± 0.64c | 9.2 ± 0.52d | 11.03 ± 0.45b | 11.91 ± 0.65a | 9.4 ± 0.58d |
Different superscript showed in a row significantly different (P<0.05).
Table 4: Polyunsaturated fatty acids (Each values is the mean of three values ± SD) of developing eggs and 1st zoeal stage of M. idella idella.
Amino acids
The total essential amino acids were showed increasing trend from the egg stage I to IV. However they were little bit low in Ist zoea stage when compered to egg stage IV. But the differences are statistically different except egg stages IIIrd and IVth. Totally 10 essential amino acids were observed. Among 10 amino acids the essential amino acids viz leucine, lysine, methionine and arginine was increased gradually as development progressed in egg stages. However valine, isoleucine, phenylalanine and threonine had a change of ‘hig-low-hig’ (Table 5).
| Amino acids | Egg stage-I | Egg stage-II | Egg stage-III | Egg stage-IV | I zoea stage |
|---|---|---|---|---|---|
| Leucine | 3.25 ± 0.22c | 3.64 ± 0.22b | 3.95 ± 0.20ab | 4.12 ± 0.26a | 4.02 ± 0.15ab |
| Lysine | 3.22 ± 0.19c | 3.34 ± 0.15bc | 3.64 ± 0.29ab | 3.88 ± 0.28a | 3.68 ± 0.19ab |
| Valine | 2.92 ± 0.21ab | 2.74 ± 0.22b | 3.14 ± 0.29a | 3.24 ± 0.25a | 3.19 ± 0.16a |
| Isoleucine | 2.36 ± 0.28ab | 2.42 ± 0.29ab | 2.62 ± 0.25a | 2.24 ± 0.18ab | 2.18 ± 0.10b |
| Phenylalanine | 2.02 ± 0.17a | 2.00 ± 0.12a | 2.14 ± 0.12a | 2.21 ± 0.20a | 2.16 ± 0.17a |
| Threonine | 1.86 ± 0.18a | 2.04 ± 0.15a | 2.16 ± 0.07a | 2.14 ± 0.09a | 2.03 ± 0.18a |
| Methionine | 1.08 ± 0.21b | 1.22 ± 0.15ab | 1.38 ± 0.28ab | 1.46 ± 0.23ab | 1.52 ± 0.21a |
| Arginine | 2.98 ± 0.19b | 3.12 ± 0.21ab | 3.23 ± 0.22ab | 3.42 ± 0.29a | 3.38 ± 0.21a |
| Histidine | 1.42 ± 0.31a | 1.41 ± 0.18a | 1.56 ± 0.21a | 1.48 ± 0.20a | 1.34 ± 0.21a |
| Tryptophan | 1.88 ± 0.24a | 1.92 ± 0.22a | 1.54 ± 0.22ab | 1.22 ± 0.21bc | 1.12 ± 0.11c |
| Total EAA | 22.99 ± 2.30d | 23.85 ± 2.65c | 25.36 ± 2.94a | 25.41 ± 4.08a | 24.62 ± 2.85b |
Table 5: Essential amino acids (%, dry weight) in M. idella idella during different embryonic development stages and I zoeal stage (Values are mean of three values ± SD).
As in essential amino acids, non essential amino acids were also followed almost similar trend. Among total 9 amino acids, 5 amino acids (glutamine, asparagine, glycine, cystine and alanine) were gradually increased from egg stage I to IV. But other non essential amino acids (serine, tyrosine, proline and taurine) were irregular (Table 6). Even though the total nonessential amino acids were gradually increased from egg stage I to egg stage IV. The increment is statistically significant.
| Amino acids | Egg stage-I | Egg stage-II | Egg stage-III | Egg stage-IV | I zoea stage |
|---|---|---|---|---|---|
| Gultamine | 5.12 ± 0.15c | 5.16 ± 0.17c | 5.68 ± 0.26b | 6.08 ± 0.21a | 5.89 ± 0.27ab |
| Asparagine | 3.32 ± 0.21c | 3.48 ± 0.31bc | 3.72 ± 0.17b | 4.14 ± 0.15a | 3.81 ± 0.21ab |
| Glycine | 2.06 ± 0.21b | 2.12 ± 0.11b | 2.32 ± 0.21ab | 2.68 ± 0.19a | 2.44 ± 0.31ab |
| Alanine | 2.04 ± 0.18c | 2.16 ± 0.27bc | 2.31 ± 0.22bc | 2.86 ± 0.21a | 2.54 ± 0.30ab |
| Serine | 2.03 ± 0.26a | 1.99 ± 0.18a | 2.24 ± 0.13a | 2.25 ± 0.25a | 2.04 ± 0.27a |
| Cystine | 1.76 ± 0.21c | 2.04 ± 0.15bc | 2.42 ± 0.26ab | 2.74 ± 0.13a | 2.36 ± 0.18ab |
| Tyrosine | 1.72 ± 0.08b | 2.04 ± 0.14ab | 1.92 ± 0.07ab | 2.06 ± 0.13ab | 2.13 ± 0.12a |
| Proline | 1.68 ± 0.17a | 1.76 ± 0.19a | 2.02 ± 0.14a | 1.92 ± 0.11a | 1.76 ± 0.15a |
| Taurine | 1.72 ± 0.24ab | 1.84 ± 0.16a | 1.32 ± 0.13c | 1.40 ± 0.39bc | 1.28 ± 0.20c |
| Total NEAA | 21.45 ± 2.15d | 22.59 ± 2.69c | 23.95 ± 2.72b | 26.13 ± 2.95a | 24.25 ± 2.75b |
| Total Amino acids | 22.99 ± 2.30d | 23.85 ± 2.65c | 25.36 ± 2.94a | 25.41 ± 4.08a | 24.62 ± 2.85b |
| (EAA+NEAA) | 44.44 ± 4.46d | 46.44 ± 4.47c | 49.31 ± 0.68ab | 51.54 ± 0.68a | 48.87 ± 0.68b |
Different superscript showed in a row significantly different (P<0.05).
Table 6: Non essential amino acids (%, dry weight) in M. idella idella during different embryonic development stages and Ist zoeal stage (Values are mean of three values
± SD).
In the present study the protein content in egg stage I was maximum (69.32%) due to the deposit of yolk. But the protein levels started decreasing when the development progressed. As the embryo developing, the yolk deposited is utilized for the development. This may be the reason for the low value of protein seen in egg stage IV. According to Radhakrishnan [14] the protein values decreased gradually as developing stages proceeds in Portunus pelagicus and
Portunus sanguinolentus. The same observation was made by Sumitra et al. [7], in Emerita holthuisi and Krishnan [15] in Thalamita crenata. Needham [16] stated that the terrestrial animal utilized0020fat as the source of energy for their development whereas in aquatic forms, protein was the source of energy. The protein content of the yolk is important for the tissue differentiation and organization particularly for the cuticle layers, muscle, the digestive and nervous system [17]. Barnes, Pandian and Katre [18,19] reported that the protein in developing eggs is progressively depleted and they also suggested the possible utilization of protein during embryogenesis to meet the metabolic demand.
The present study reveals that carbohydrate value showed an increasing trend from egg stages I to IV. The carbohydrate played major part for the development of earlier stages in. Macrobrachium rosenbergii [20]. Sumitra [7] also reported gradual increasing trend in M. idella of Cochin backwaters. But in Portunus pelagicus and Portunus sanguinolentus [14] and T. crenata [15] the carbohydrate value showed gradual decrease when development proceeded. Ajith Kumar [21] stated that the carbohydrate showed an increasing trend from I to IV stages in Macrobrachium idella idella as in the present study.
Lipid plays a central role in the embryonic metabolism as they represent the most important energy source and form atleast 60% of the total energy expenditure of the developing crustacean embryo [22]. Large decapod eggs have a relatively high fat percentage and low density when compared to small eggs [23]. Lipid value obtained in the present study varied between 15.86% (stage-I) to 5.96% (stage- IV) which is comparatively higher than that of Macrobrachium idella idella available from Cochin backwaters [7] but showed slight variation with who studied in the same species from Vellar estuary [21]. But, lipid values are shown gradual increase with development in Portunus pelagicus and Portunus sanguinolentus [14]. Needham [16] classified the crustacean eggs as cleidoic and non-cleidoic types of eggs. The cleidoic eggs are not dependent on the environment for water and salt (ash); oxidation of protein is suppressed to considerable extend and fat oxidation is greatly enhanced, serving as main source for the embryonic metabolism. A similar pattern has been reported for Callinectes sapidus [24] and Xantho bidentatus [17]. The crab Caridina nilotica and prawn Macrobrachium malcolmsonii derived more energy from fat [25,26]. In the present study it is clear that the lipid value at initial stage was higher but reducing when the egg started developing especially during III and IV stages. This result attributes that fat also play major role during embryonic development.
Developing eggs of crustaceans absorb water [27]. In Macrobrachium rosenbergii the egg volume increased slowly from fertilized egg stage to zoea stage indicated that there were prominent correlation between egg volume and water content [20]. The absorption of water increases the internal osmotic pressure resulting in osmotic hatching of egg and accumulation of water decreases the specific gravity of developing eggs and finally the free floating planktonic larvae released. The increase in water content of the eggs is due to the absorption of water through the egg membrane of the retention of metabolic water since water is the ‘byproduct’ of respiration. The oxidation of 100 gm of lipid releases 107.7 gm of water, the same amount of protein produces 111.3 gm of water and carbohydrate oxidation liberates 55.5 gm of water [28]. The weight of eggs during zoea stage increased since fertilized stage, mainly because of the increased water content. Water provided a liquid environment for embryo and the higher water pressure during zoea stage might be cause of the embryo breaking through the egg membrane in preparation for hatching [20].
In the present study the lipid acted as the most important energy source in the embryonic metabolism of Macrobrachium idella idella. The total lipid content of egg stage-I (15.86 ± 0.18%) expected for formation of the organs, lipid was mainly used as a source of energy. Rapid decrease of total lipids in egg stages-III and IV was closely associated with the formation and development of many organ anlages. Brain anlage and heart anlage came into being. Compound eyes turned larger, blood started to flow and heart beats at about 200 times per minutes in zoea stage [29,30]. More energy was needed and some fatty acids turned into component of organs in these two stages. In zoea stage, lipids in yolk were used up. The zoea stage of Macrobrachium rosenbergii lasted about 90 h, and more energy was needed. So some fatty acids were probably used in the synthesis of organs during these stages [20]. The consumption pattern of different fatty acids in eggs of Macrobrachium rosenbergii during the embryonic development did not differ markedly from that of other crustaceans [31-33]. The most important fatty acids were C16:0, C18:0, C16:1, C18:1n-9 and C18:2n-6 in eggs of Macrobrachium rosenbergii. In the present study most important fatty acids components in Macrobrachium idella idella were C16:0, C18:0, C18:1n-9 as in Macrobrachium rosenbergii. The first zoea stage of Macrobrachium idella idella is non-feeding stage, the remaining nutriment favours their independence of external energy resources when external feeding begins and would increase the changes for the first successful moult. Yao Jun-jie [20] Reported that in Macrobrachium rosenbergii the remaining yolk in the eggs are utilized by the first larval stage as the first larva is non-feeding stage.
It has been revealed that the amino acids [34], monosaccharides [35] and nucleosides [36] could be transported into the embryos of the marine invertebrates. Rosa et al. [33] assumed that embryos of Nephrops norvegicus could absorb some compounds. In the present study, the content of essential amino acids increased in eggs, may be the result of organic compounds could be transported into the embryos of Macrobrachium idella idella. Even though the amino acids (EAA) and total amino acids (TAA) contributed to the completion of the embryonic development of Macrobrachium idella idella the ratio of EAA and TAA content remained unchanged during different embryonic development stages. But when compared first zoea stage to the egg stages the EAA and TAA were less than egg stages. It may be because of the first zoea stage does not feed externally and utilizes the yolk present inside the zoea stage. And the zoea doesn’t have the capacity of absorbing organic compounds from the external water source as in the egg stages. The unchanged EAA and TAA were also determined in the eggs of Macrobrachium rosenbergii [20].
The increase in TAA is indicative of the fact that protein in yolk act as the main structural substance during embryonic development of Macrobrachium idella idella. Increases in TAA were also reported in the eggs of Nephrops norvegicus [33] and in Macrobrachium rosenbergii [20]. Quantitively the most important amino acids were leucine, lysine, valine, arginine and glutamic acid during the embryonic development of Macrobrachium idella idella. Regarding the function of single amino acid, leucine is a ketone-producing amino acid. It could be transformed into acetyl-CoA and acetyl-acetic acid, which are important intermediates in carbohydrate and lipid metabolism [37]. Arginine was proven to be crucial in energy metabolism by maintaining glycolysis under hypoxic conditions [38].Valine is a carbohydrate-producing amino acid and may be associated with carbohydrate metabolism through citric acid cycle. Content of glutamic acid was high during embryonic development, which may have resulted from nitrogen metabolism in the eggs of Macrobrachium idella idella. Glutamic acid turned into glutamine, which is deaminated to produce NH3 [38]. NH3 can be excreted along with Cl-. An increase in the content of NH4Cl after the blastula stage also suggests that NH4+ and Cl- are being excreted together. Tyrosine can be used to synthesize melanin [38], which plays central role in the accumulation of compound eye pigments. High content of these amino acids is closely correlated with their important role in the embryo.