Journal of Glycobiology
Open Access
ISSN: 2168-958X
ISSN: 2168-958X
Research Article - (2013) Volume 0, Issue 0
Background: Malignant Peritoneal Mesothelioma (MPM) is a rare asbestos related peritoneal neoplasm. Survival is normally poor however, cytoreductive surgery and hyperthermic intraperitoneal chemotherapy has increased survival in some patients. Hence, new therapies are needed. MUC1 is a glycosylation dependant protein associated with tumour invasiveness, metastasis and chemo resistance. Bromelain, a cysteine proteinase, hydrolyses glycosidic bonds, whilst N-Acetyl cysteine reduces disulphide bonds in glycoprotein. Hence, we investigated the anti-tumour effect of these agents in MUC 1 expressing MPM cell lines.
Materials and Methods: The cell lines were treated to various concentrations of bromelain, NAC and combinations of NAC + bromelain, NAC + bromelain + cisplatin or 5-FU. Their cell viabilities were assessed at 48 hours with sulfhordamine B assay. Finally, with western blotting, the effect of NAC and the combination of NAC + bromelain on cellular survival proteins were investigated.
Results: Combination of NAC (10 mM) with bromelain (75 ug/ml) showed 97% and 88% cell proliferation inhibition in YOU and PET cells, respectively. In triple combination, the addition of cisplatin to only 5.0 mM NAC and bromelain increased cytotoxicity in YOU cells but absent at 10.0 mM NAC concentration. However, in PET cells, triple combinations with cisplatin had no effect.
The addition of 5-FU in triple combinations showed an increase in cytotoxicity with 5.0 mM NAC and bromelain in YOU cells. No increase in cytotoxicity was seen with addition of 10 mM NAC. In PET cells, the addition 5-FU to 5.0 mM NAC + 50 ug/ml bromelain, enhanced cytotoxicity significantly but was absent at all other combinations.
Conclusion: The combination of bromelain and NAC may be developed as anti-tumour agents for treating MPM, with a possible role in combined therapy with current chemotherapeutic agents.
<Keywords: Bromelain; N-acetyl cysteine; Peritoneal mesothelioma; Cytotoxicity
Malignant peritoneal mesothelioma is a rare form of neoplasm that occurs in the mesothelial lining of the peritoneal cavity [1]. Asbestos is a known causative agent, whilst Simian virus 40 has been suggested as a co-carcinogen [2,3]. Diagnosis often occurs late in the course of disease, owing to non-specific symptoms [4]. The median survival in untreated patients is 9 months [5]. Current treatment regime using cytoreductive surgery with intraperitoneal hyperthermic chemotherapy (HIPEC) has dramatically improved survival in some patients with known survival beyond 7 years [6]. However, owing to poor survival in many patients, there is an urgent need for new therapies. Currently several therapies are in development [7,8], although none has been recommended for clinical application.
Further, it is also known that MPM tumours express substantial amount of MuC1 that is associated with chemoresistance, invasiveness, metastasis and robust cellular proliferation [9-11]. MuC1 is a transmembrane glycoprotein that has several glcosylated motifs both in the extracellular, intra-cellular and in the cytoplasmic region [12,13]. The extracellular region (MuC1-N), after detachment from the cytoplasmic region (MuC1-C), forms a gelatinous protective layer engulfing the whole cell and hence serves as a barrier against chemotherapeutic drugs [13]. The extracellular region of MuC1-C is glycosylated at asn 36 and serves as a docking region for galacetin-3 bridge that links epidermal growth factor receptors (EGFR) and other tyrosine kinase receptors (FGFR-3, ERBB2, PDGFR-β) to MuC1 and serves to stabilize these receptors [14-16]. Further, the MuC1 domain in the cytoplasm (MuC1-CD) contains seven tyrosine phosphorylation sites, of which five have been indentified to phosphorylate on binding with proteins that carry SH2 domain [16] there by activating several survival pathways [17-19].
Bromelain is a cysteine proteinase that has been extracted from pineapple (Ananas comosus) [20] and it has been investigated for its mucolytic [21,22], anti-inflammatory [23,24], wound debridment [25,26], antithrombotic [27] and anticancer properties [28-30]. It has also shown anti-tumuroigenic properties, both in vitro and in vivo, in a number of cancers, particularly in breast, pancreas and a variety of other cancers [29-31]. A recent review on the anticancer properties of bromelain have emphasized that its proteolytic component may be mainly responsible for its anti-tumour effect [32]. Since, bromelain contains a number of other components such a cellulases, phosphatases, hydroxylases [33-35], along with a number of other protein components, the antitumor properties cannot be singly attributed to its proteolytic activity. Since, bromelain hydrolyses glycosidic linkages in glycoproteins [36]; it is inevitable that glycosylated moieties that provide oncogenic survival pathways in cells may be affected. Further, the proteolytic action of bromelain may also disrupt the glycosidic linkages in the secreted mucin and hence disrupt the mucinous barrier thereby enabling a better passage for cytotoxic drugs. This has been recently demonstrated in pancreatic cancer, with 5-Fu [13].
N-Acetyl Cysteine (NAC) is an agent commonly used for treating acetaminophen toxicity [37], however it is also used for reducing the viscosity of mucus produced in cystic fibrosis and chronic obstructive pulmonary disorder (COPD) [38,39]. NAC by virtue of its chemical composition has the ability to reduce disulphide bonds (SS) to sulphydryl (S-H) bonds [40] and hence it may act as a scission, reducing the intermolecular linkages between the glycoprotein. MuC1 has three domains, the external domain (MuC1-ED), transmembrane domain (MuC1-TD) and domain in the cellular cytoplasm (MuC1- CD). The MuC1-ED forms a jelly like barrier layer after being severed from MuC1-TD [12]. This barrier layer is a polymer that contains both intermolecular disulphide bridges as well as glycosidic linkages [12]. NAC is able to reduce S-S bond to S-H bond and thereby disintegrate the intermolecular disulphide linkages and hence render the mucinous barrier more penetratable. Further, there are five identified phosphorylating regions located in the MuC1-CD which are capable of interacting with SH2 motifs of proteins molecules and thereby initiate oncogenic pathways [16]. NAC may interact with these regions and hence contribute to antitumor effects. Hence, NAC has the chemical ability to interfere with the molecular configuration of MuC1 and thereby provide therapeutic benefits.
In the past, we have demonstrated how the viability of malignant peritoneal mesothelioma cells was affected by bromelain exposure and that the cytotoxicity of cisplatin was enhanced with adjuvant bromelain treatment (Chapter 9). In the current work we set out to investigate how the presence of both bromelain and NAC affect PET and YOU cell survival. Further, we also investigated how the combination of these two agents affected cytotoxicity induced by cisplatin and 5-Fu.
The PET and YOU cells (MPM cells) were from our cell bank. All chemical reagents used in this experiment were from Sigma chemicals or of analytical grade obtained from reputable companies. The primary and secondary antibodies for western blot were procured from reputable companies, as given in Table 1.
| Antibody | Isotype | Dilution | I T | Supplier |
|---|---|---|---|---|
| AKT | Rabbit IgG | 1:1000 | ON/4 O C | Cell Signalling, USA |
| BCL2 | “ | 1:200 | RT/2hr | Santa Cruz Biotech, USA |
| BCL-xl | “ | 1:200 | “ | “ |
| Caspase-3 | “ | 0.5ug/ml | ON/4 O C | R&D System, USA |
| Caspase-7 | “ | 1:1000 | “ | Cell Signalling, USA |
| Caspase-8 | Goat IgG | 0.5ug/ml | “ | R&D System, USA |
| Caspase-9 | Mouse IgG | 1ug/ml | “ | “ |
| IkkB | Rabbit IgG | 1:1000 | “ | Cell Signalling, USA |
| LC3B | “ | “ | “ | “ |
| NFkB | “ | “ | “ | “ |
| p-IkkB | “ | “ | “ | “ |
| p-NFkB | “ | “ | “ | “ |
| p53 | “ | 1:200 | RT/2hr | Santa Cruz Biotech, USA |
| p-AKT | “ | 1:1000 | ON/4°C | Cell Signalling, USA |
| PARP | “ | “ | “ | “ |
| Β- Actin | Mouse IgG | “ | RT/2he | Sigma Aldrich, Australia |
| MUC1 | Rabbit IgG | “ | ON/4°C | ABD- Serotech, USA |
| Anti- mouse IgG/HRP | - | “ | ON/4°C | Cell Signalling, USA |
| Anti- rabbit IgG/HRP | - | “ | “ | “ |
| Donkey anti-goat Ab | - | “ | RT/2hr | Santa Cruz Biotech, USA |
IT = Incubation Time; ON = overnight 14 hours; RT = Room temperature (25 deg Celsius)
Table 1: shows the various primary and secondary antibodies used for the detection of survival cellular proteins, along with their respective dilutions.
Cell cytotoxicity assays
YOU and PET cells were maintained in RPMI 1640 medium supplemented with 10% FBS, 50 units/ml penicillin and 50 units/ml streptomycin. For proliferation assay, the cells were seeded at a density of 5×103 cells/well in 96-well plates. After 24 hours incubation, the cells were exposed to the medium-containing drugs (single agents or combination) and subsequently after a lapse of 4 or 48 hours, SRB assay was performed to determine the number of adherent cells. All cytotoxicity experiments were performed three times and their values were averaged.
SRB assay
At the end of the incubation, cells were fixed with ice-cold 10 % trichloroacetic acid for 30 minutes followed by 5 washes with tap water. Cells were then stained with 0.4% sulforhodamine B (SRB) for 15 minutes followed by 5 washes in 1 % acetic acid. The plate was allowed to dry and SRB was dissolved in 10 mM Tris. The plate was then read at 570 nM [13].
Preparation of drugs
Bromelain was dissolved in Tris buffer 10 mg/ml in Tris buffer (pH 7.0) and adjusted to neutral pH with analytical grade sodium hydroxide. This was then sterile filtered and the required quantity was added to fresh media.
NAC solution was prepared by dissolving medical grade NAC in culture media (RPMI), pH adjusted to 7.0 with 0.1 M sodium hydroxide and finally sterile filtered. The required quantity was then added to fresh media.
Cisplatin was dissolved in DMSO and stored at 4 deg Celsius; similarly 5-Fu was also dissolved in DMSO and stored at 4°C. The required quantity was added to fresh media to obtain the desired concentration.
Western blot assays
PET and YOU cells were grown in 250 ml flasks until 50% confluent. They were then treated with bromelain at the required concentration in fresh media. After a period of 48 hours, the media was removed, washed with PBS and then the cells were lysed (lysing buffer containing 10% ant protease and 10% anti- phosphorylase) at 4°C. The lysate was centrifuged at 10000 g (10 mins at 4°C), the protein concentration of the supernatant was determined using the method of Lowry et al. [40].
For western blot, 50ug of protein was resolved on sodium dodecyl sulphate (SDS) polyacrylamide gels followed by electrotransfer onto a PVD membrane. The blots were blocked with 5% non fat dry milk, probed with respective primary antibodies according to manufacturer’s recommendations, and detected by horseradish peroxidise-conjugated antimouse, anti rabbit IgG or Donkey- anti goat antibody, as given in Table 1, using chemiluminescence kit by Millipore and visualized by Versa Doc Imaging System.
Effect of bromelain + NAC on the viability of YOU and PET cells
Bromelain affects cell survival in both the cell lines and cell death was proportional to the concentration of bromelain in the media. The IC50 values for YOU and PET were 55 and 54 ug/ml bromelain, respectively (Figure 1 A and 1C). Conversely, NAC at 10 - 50 mM, has very minimal inhibitory activity on YOU cells with maximum inhibition of 10% at 25 and 50 mM, Figure 1B. Noticeably, NAC had no inhibitory activities on PET cells (Figure 1D).
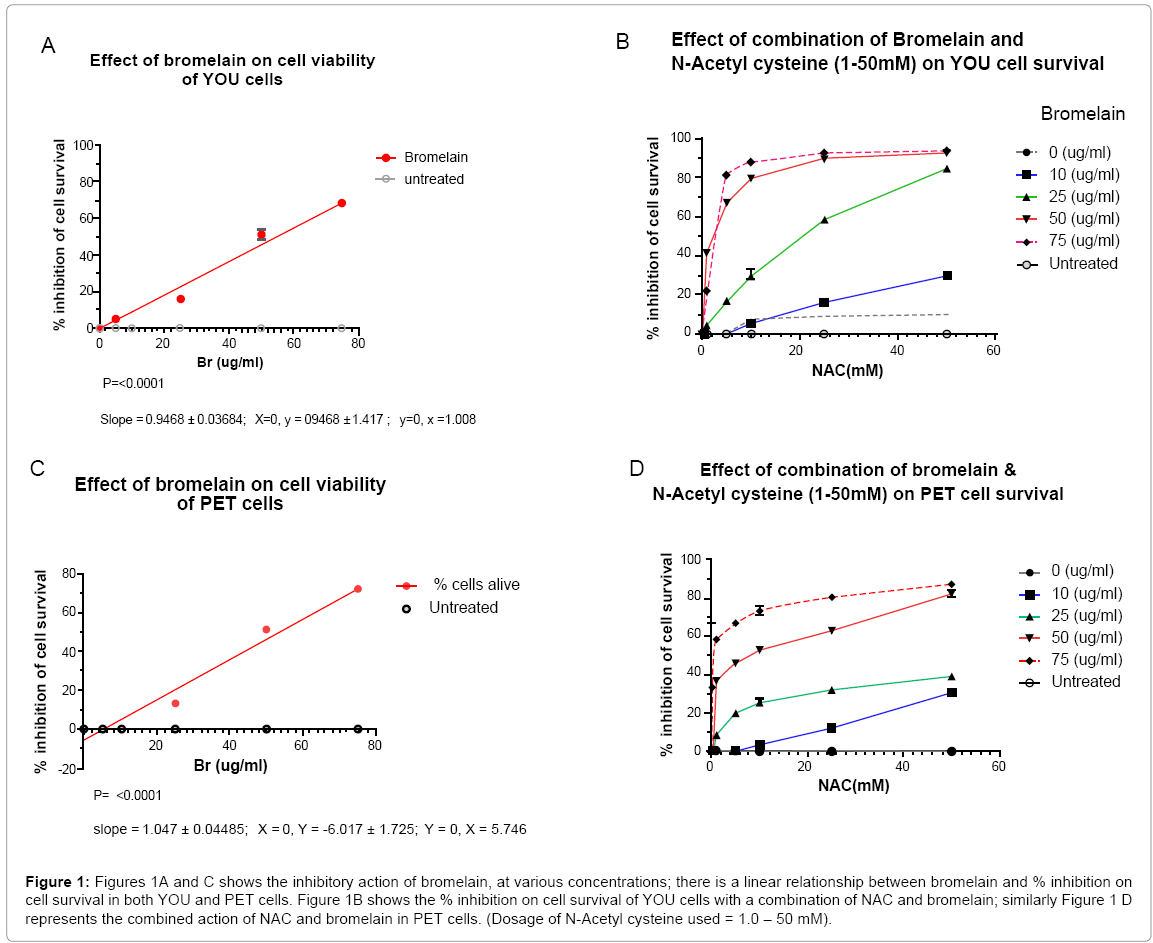
Figure 1: Figures 1A and C shows the inhibitory action of bromelain, at various concentrations; there is a linear relationship between bromelain and % inhibition on cell survival in both YOU and PET cells. Figure 1B shows the % inhibition on cell survival of YOU cells with a combination of NAC and bromelain; similarly Figure 1 D represents the combined action of NAC and bromelain in PET cells. (Dosage of N-Acetyl cysteine used = 1.0 – 50 mM).
The maximum inhibitory activity observed with the addition of 75 ug/ml bromelain to 50 mM NAC indicated a 97% kill for YOU cells as compared to PET of 88%, indicating that PET cells were more resistant to this combination treatment (Table 2). The difference in percentage of inhibitory activities between 50 and 75 ug bromelain when added to 50 mM NAC was very small (4%) for both YOU and PET cells indicating that either of 50 or 75 ug/ml bromelain may be used with 50 uM NAC to achieve the best cell kill ( maximum efficacy).
| YOU CELL LINE +50 mM NAC | PET CELL LINE +50 mM NAC | |||||
| Br (ug/ml) | % I(A) | % I(B) | B - A | % I(C) | % I(D) | D – C |
| 0 | 0 | 10 | 10 | 0 | 0 | 0 |
| 10 | 10 | 30 | 20 | 0 | 30 | 30 |
| 25 | 23 | 85 | 62 | 20 | 37.5 | 17.5 |
| 50 | 45 | 94 | 49 | 46 | 82.5 | 36.5 |
| 75 | 69 | 96 | 27 | 73 | 87.5 | 20.5 |
A = % inhibition of cell viability (CV) by bromelain as a single agent; B = % inhibition of CV with addition of 50 mM NAC to bromelain; C= % inhibition of CV by bromelain as a single agent; D = % inhibition of CV with addition of 50 mM NAC to bromelain; I = inhibition; NAC = N- Acetyl Cysteine. B-A = increase in inhibition of CV with addition of 50 mM NAC
Table 2: Shows the maximum inhibition in cell survival with bromelain as a single agent and when combined with 50 mM N- Acetyl Cysteine (NAC).
The effect of different concentrations of bromelain added to 50 mM NAC as shown in Table 2, indicates that the largest difference in % cell kill (inhibition) is observed at 25 ug/ml bromelain (bromelain by itself = 24 % cell kill, whilst with 50 mM NAC, the % cell kill was 84%, a difference of 60% for YOU cells. Similarly for PET cells, at 50 ug/ml bromelain, gave a % cell kill of 37%. This again indicated that PET cells were more resistant. Further, from this data, it was evident that the addition of NAC to bromelain enhances its cytotoxicity.
On probing further into the activities of adding NAC to bromelain, it is clear that for YOU cells the addition of 10 - 50 mM NAC to 25 ug bromelain and above would achieve sufficient cell kill, although at concentrations of 50 and 75 ug/ml bromelain, the % cell kill approaches 100 % (Figure 2A) A similar scenario exists for PET cells although the % cell kill achieved at higher concentrations of bromelain (50 -75 ug/ ml) is slightly lower than that for YOU cells (Figure 2B), a difference of approximately 10%, which again indicates the resistant nature of PET cells to this treatment (Figure 2C and 2D).
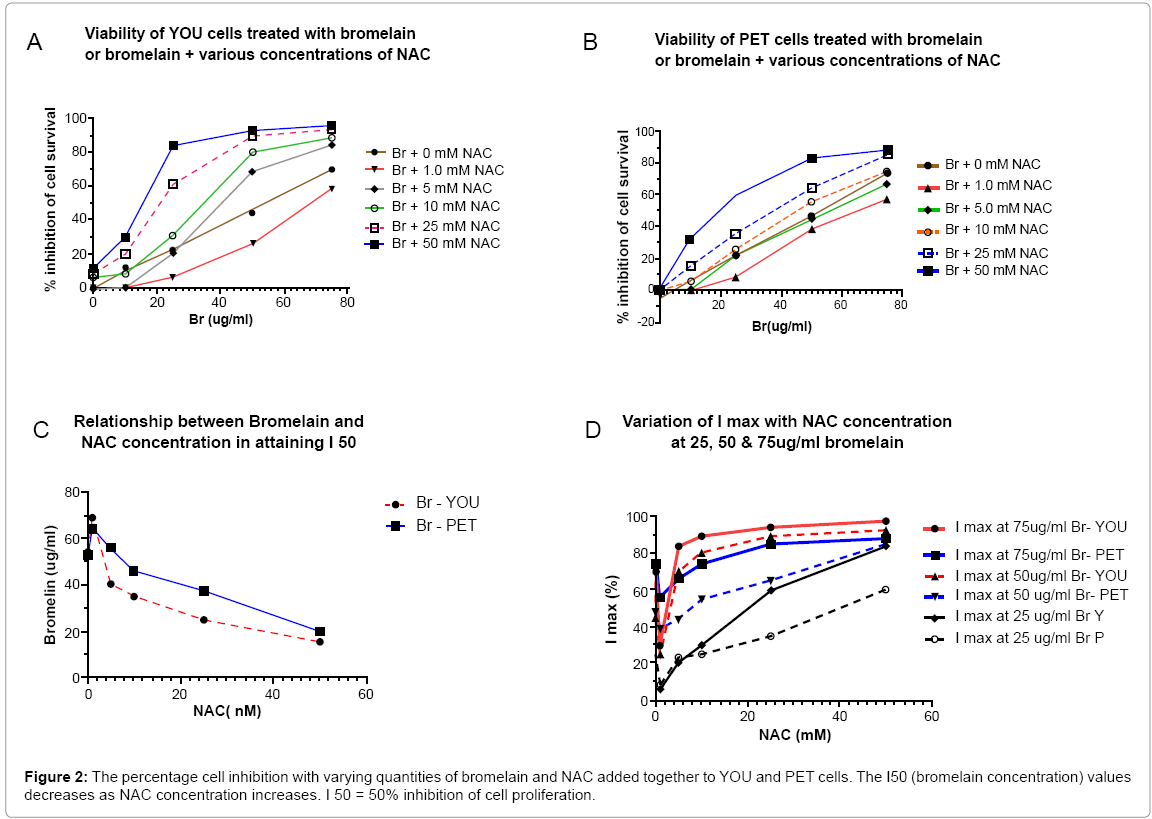
Figure 2: The percentage cell inhibition with varying quantities of bromelain and NAC added together to YOU and PET cells. The I50 (bromelain concentration) values decreases as NAC concentration increases. I 50 = 50% inhibition of cell proliferation.
It appears that the addition of NAC > 5.0 mM to bromelain enhances the effect of bromelain since it reduces the bromelain required to achieve IC50% for YOU cells (Figure 3A). However, for PET cells only >10.0 mM NAC addition to bromelain showed a similar trend, although the effect of adding NAC to bromelain for YOU cells was much greater compared to PET cells.
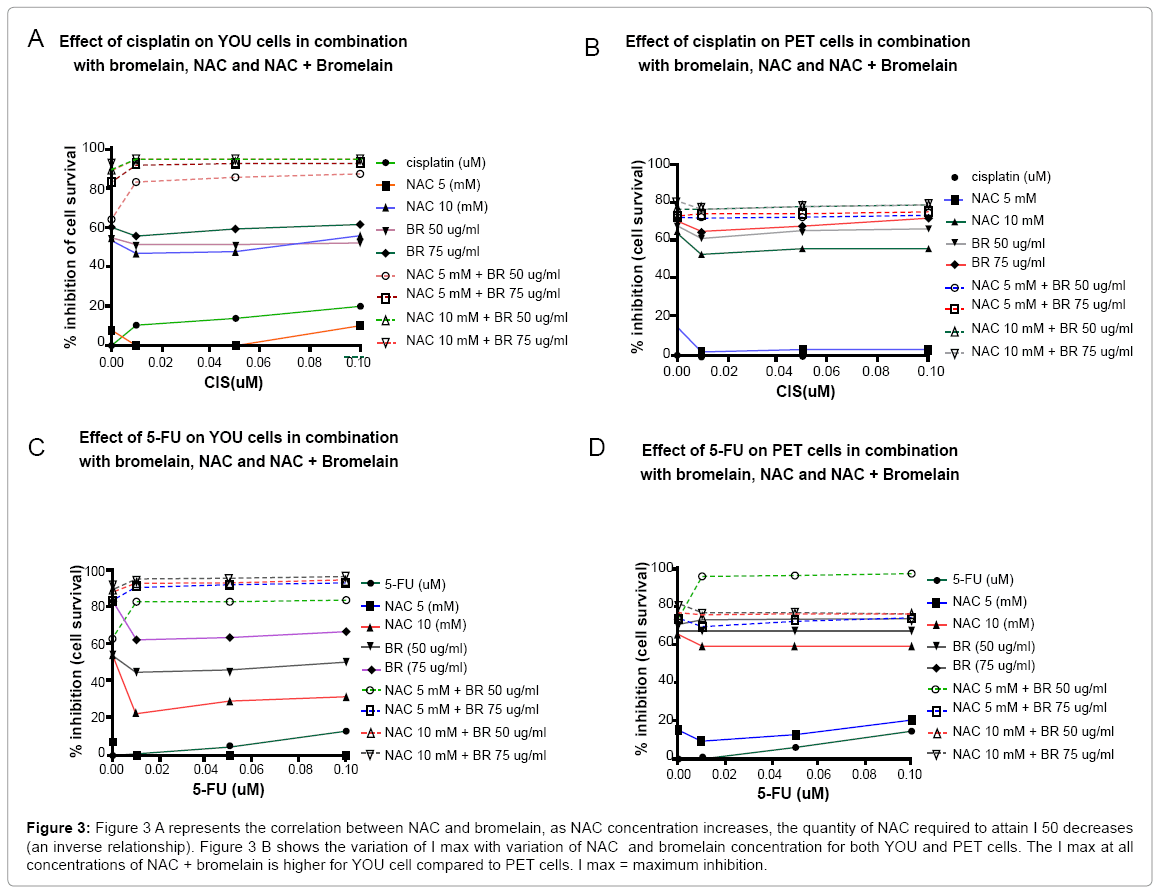
Figure 3: Figure 3 A represents the correlation between NAC and bromelain, as NAC concentration increases, the quantity of NAC required to attain I 50 decreases (an inverse relationship). Figure 3 B shows the variation of I max with variation of NAC and bromelain concentration for both YOU and PET cells. The I max at all concentrations of NAC + bromelain is higher for YOU cell compared to PET cells. I max = maximum inhibition.
Finally Table 3 shows the reduction of bromelain concentration required to achieve 50% Inhibition with the addition of increasing quantities of NAC, reinforcing what has been explained earlier. Further examining the % inhibition (cell kill) observed at different concentrations of bromelain (25, 50, 75 ug/ml) with increasing concentrations of NAC, it is apparent that addition of 10mM NAC to 50 or 75 ug/ml bromelain will achieve at least a 80% cell kill for YOU cells (Figure 2A). On the other hand for PET cells, either a combination of 25 mM NAC + 75 ug/ml bromelain or 50 mM NAC + 50 or 75 ug/ml bromelain will achieve a cell kill greater than 80% (Figure 2B).
| Y O U C E L L | P E T C E L L | |||||||
|---|---|---|---|---|---|---|---|---|
| NAC 50% I at (mM) BR (ug/ml) | Max I (%) at bromelain (ug/ml) | 50% I at | Max I (%) at bromelain (ug/ml) | |||||
| 25 | 50 | 75 | at BR (ug/ml) | 25 | 50 | 75 | ||
| 0 | 54.0 | 24 | 45 | 70 | 53 | 23 | 48 | 74 |
| 1.0 | 70 | 6.0 | 25 | 30 | 66 | 8 | 39 | 56 |
| 5.0 | 40.0 | 20 | 70 | 84 | 56 | 23 | 44 | 66 |
| 10.0 | 34.8 | 30 | 80 | 89 | 46 | 25 | 55 | 74 |
| 25.0 | 25.0 | 60 | 89 | 94 | 37.5 | 35 | 65 | 85 |
| 50.0 | 15.5 | 84 | 93 | 97 | 20 | 60 | 84 | 88 |
I = Inhibition; The values in the table are obtained from graph 1, 2 ; 50% I at bromelain concentration refers to bromelain concentration at which 50% inhibition is seen for each value of NAC concentration; Max I (%) refers to maximum inhibition observed on addition of 25, 50, 75 ug/ml Bromelain; NAC = N- Acetyl Cysteine; BR = Bromelain. Noticeably the addition of 1.0 mM NAC to various concentrations of bromelain inhibits the action of bromelain for both YOU and PET cells, however, it recovers at 10 mM NAC.
Table 3: Shows the 50% inhibition of cellular proliferation and maximum inhibition of cellular proliferation at various NAC concentrations.
A comparison of the maximum inhibition (I max) for the two cell lines (Figure 3B), at various NAC and bromelain concentrations shows that for similar addition of NAC and bromelain, PET cells shows less inhibition compared to YOU. This further confirms the difference between the two cell lines on their reactivity to the agents (Figure 3C and 3D).
Cisplatin cytotoxicity, in combination with bromelain and N-Acetyl-cysteine on YOU and PET cells
YOU cells: Cisplatin as a single agent has a small (<20%) inhibitory effect on YOU cell proliferation. The effect of cisplatin (0.01 - 0.1 mM), in combination with either bromelain (50, 75 ug/ml) or NAC (5, 10 uM), showed a inhibition on cell viability of <60% (Figure 4). Further, the addition of 0.01 uM cisplatin to either bromelain or NAC seems to depress the inhibitory activities, however this depressive effect seems to be lost at higher cisplatin value of 0.1 uM. Remarkably, the combination of three agents such as NAC, bromelain and cisplatin seems to show an inhibitory effect of 85-96%. The addition of cisplatin to bromelain + NAC shows an increase in inhibition (10-24 %) for the lower NAC concentration of 5 uM. However at higher NAC (10uM) concentration + bromelain and cisplatin, the inhibitory effect appears to be almost at par with just NAC + bromelain (NAC + bromelain = 93% vs. the addition of cisplatin = 95%). This indicates that the addition of cisplatin to 10 mM NAC and bromelain does not really have any significance.
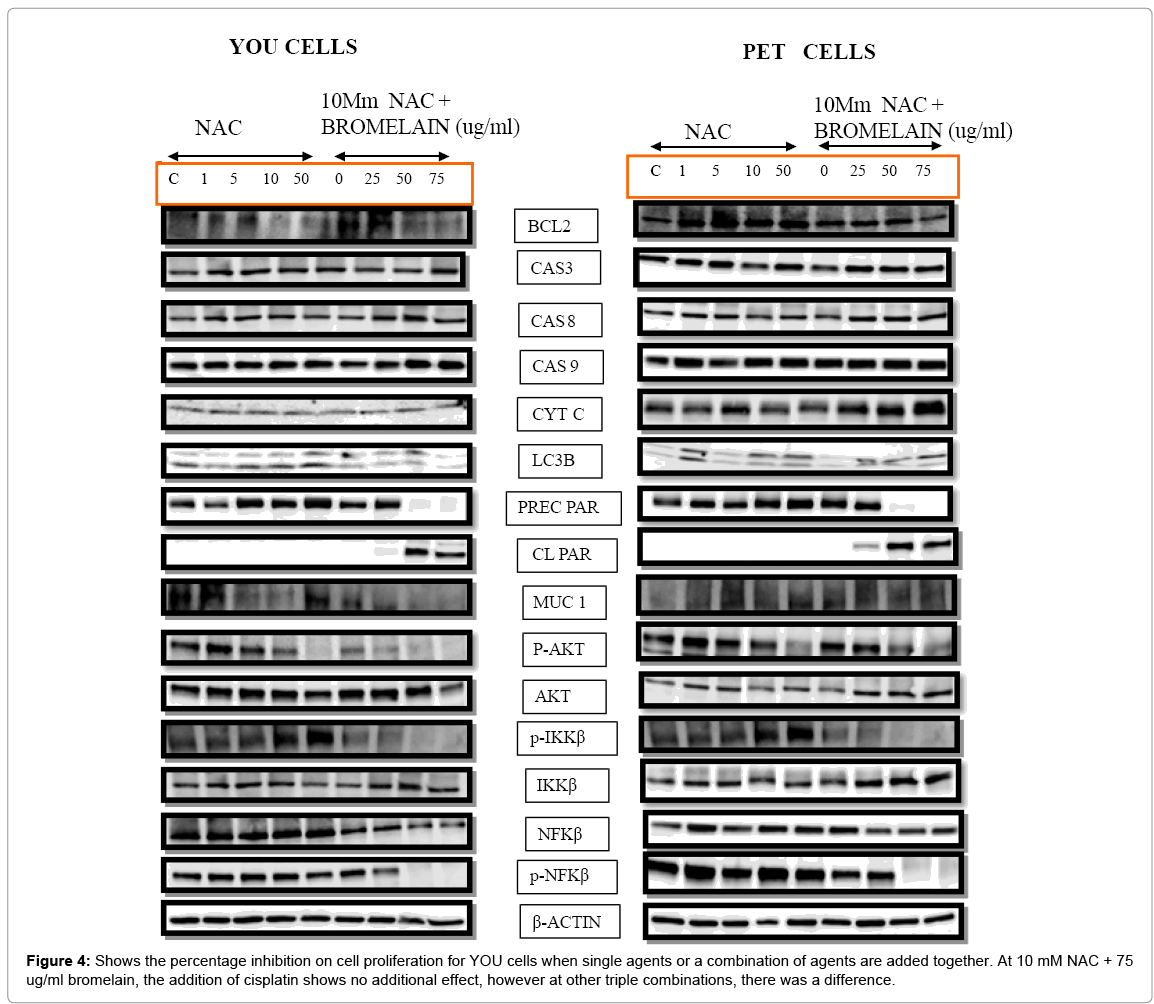
Figure 4: Shows the percentage inhibition on cell proliferation for YOU cells when single agents or a combination of agents are added together. At 10 mM NAC + 75 ug/ml bromelain, the addition of cisplatin shows no additional effect, however at other triple combinations, there was a difference.
PET cells: Cisplatin (0.01 - 0.1 uM) as a single agent has no effect on PET cells. The effect of cisplatin (0.01 - 0.1 uM), in combination with either bromelain (50,75 ug/ml) or NAC (5,10 mM), showed inhibition of cell viability of <70% (Figure 4). The addition of 0.01 uM cisplatin to either bromelain or NAC reduced the inhibitory effect by 10 -18% that is shown by bromelain or NAC as a single agents. Only bromelain at 75 ug/ml is able to attenuate this effect with 0.1 uM cisplatin. Noticeably, the addition of the two agents (bromelain and NAC) to cisplatin did not show any increase in inhibitory effect compared to the NAC and cisplatin as dual agents. Amongst the four different combination concentrations of NAC and bromelain used, NAC 10 mM + 75 ug/ml bromelain showed the highest inhibition of about 80%. Therefore, there is an indication that the addition of cisplatin in the triple combination did not have any therapeutic value for PET cells.
On a comparative basis, PET cells are more resistant to cisplatin treatment since it had zero inhibitory effect at the concentrations investigated. Further using the triple compound treatment, the addition of cisplatin did not make a difference. Finally, the maximum inhibitory effect shown by PET cells is 80% as compared to YOU cells with 95 % inhibition indicating that PET cells are less responsive to this treatment compared to YOU cells.
5-Fu cytotoxicity, in combination with bromelain and NAcetyl -cysteine, on YOU and PET cells YOU cells
5-Fu (0.01 - 0.1 uM) as a single agent had no effect on cell viability of YOU cells. In combination with either bromelain (50, 75 ug/ml) or NAC (5, 10 mM), there was an inhibitory effect on cell viability that ranged from 10 - 65 % (Figure 4). This effect was smaller compared to either bromelain or NAC as individual agents, suggesting that 5-Fu retards the inhibitory action of bromelain or NAC as individual agents by 10-30%. However, when the three agents were used simultaneously, the inhibitory action on cell viability was increased compared to addition of dual agents (bromelain + NAC). At 5 mM NAC, the addition of either 50 or 75 ug/ml bromelain to 5-Fu showed an increase in inhibitory effect (20% increase for 5mM NAC + 50ug/ml bromelain; 8% increase for 5 mM NAC + 75 ug/ml bromelain). With higher concentrations of NAC (10 mM) + bromelain the addition of 5-Fu did not show any increase of inhibitory effect on YOU cells, indicating that at this concentration of NAC and bromelain, the addition of 5-Fu had no pharmaceutical role. A maximum cell inhibitory effect of 95% was shown by 10 mM NAC and 75 ug/ml bromelain.
PET cells
5-Fu as a single agent had a very small inhibitory effect on the cell proliferation of PET cells (maximum of 18% at 0.10 uM 5-Fu). With the addition of either NAC or bromelain to 5-Fu, a maximum inhibitory effect of 18-76% was shown at the various concentrations of NAC and bromelain that was investigated (Figure 4). When the three agents (NAC, bromelain and 5-Fu) were added together, the addition of NAC 5 mM + 50 ug/ml bromelain + 5Fu, produced the largest cell inhibitory effect (95%), compared to the other combinations investigated. Further there was a 20% increase in inhibition on addition of 5-Fu compared to NAC 5 mM + bromelain 50 ug/ml), as dual agents. This indicates that PET cell was very responsive to a specific dosage of NAC (5 mM) and bromelain (50 ug/ml), unlike YOU cells that reacted favourably at high concentrations of combinations of bromelain and NAC.
On a comparative basis, YOU cells showed maximum inhibition to just the addition of 10 mM NAC + 50 ug/ml bromelain or 10 mM NAC + 75 ug/ml bromelain, without requiring 5-Fu, whilst PET cells showed maximum inhibition of cell proliferation at 5 mM NAC + 50 ug/ml bromelain when added to 5-Fu. This difference in behaviour may be related to the higher level of MuC1 expression that is found in PET cells.
Effect of NAC and the combination of NAC with bromelain on cellular proteins in YOU and PET cells YOU cells (NAC as a single agent)
NAC, as a single agent at concentration of 1 -5 mM had no effect on BCL2, however at 10 and 50 mM, BCL 2 is down regulated dramatically (Table 4). Caspase 8, 9 are up regulated on addition of 5 - 50 mM NAC, although we did not detect the elevation of cytochrome C. LC3B an indicator of autophagy is up regulated on addition of 10 - 50 mM NAC. There is also a slight up regulation of precursor PARP between 5 - 50 mM NAC, with total absence of cleaved PARP. There was a slight down regulation of MuC1 on addition of 5, 10 mM NAC that was absent at 50 mM NAC. AKT was up regulated slightly between 5 - 50 mM NAC addition, whilst PKT was considerably down regulated at 50 mM NAC.

Table 4: It is an analysis of the western blot (Figure 4), number of arrows indicating the intensity of the band. The table shows that both the cells are mainly undergoing apoptosis, with a minor component of autophagy, in both the treatment regime. MUC 1 seems to be more affected with combination treatment (NAC + bromelain) in YOU cells compared to PET cells. Further autophagy seems to more highly expressed in PET cells compared to YOU cells.
There was a slight increase in p-Ikkβ at 5 - 50 mM NAC, whilst there was a total absence of effect on Ikkβ at all concentration of NAC used. A slight increase in NFkβ with a decrease in p- NFkβ was observed at 10 -50 mM NAC.
Based on the cellular parameters monitored, NAC as a single agent kills the cells through apoptosis and autophagy, although on a slightly lower scale. MuC 1 is only slightly down regulated by NAC.
YOU cells (NAC + bromelain)
The addition of 50 or 75 ug/ml bromelain to 10mM NAC had a dramatic effect on cellular parameters as shown in Table 4. BCL2, Precursor PARP, MuC1, p-AKT, p-Ikkβ, NFkβ and p-NFkβ were all down regulated, whilst Caspase 8, 9, cleaved PARP and Ikkβ were all upregulated, indicating that apoptosis was the main pathway of cell death that took place at an accelerated phase compared to the effect of NAC alone.
PET cells (NAC as a single agent)
The addition of NAC at 1-50 mM did not have any effect on BCL2 expression. At 1 -5 mM NAC, the expression of caspase 3, 9, LC3B and NFkβ were slightly upregulated, whilst p-AKT and MuC 1 were slightly down regulated. At 10 mM NAC , caspase 3, 9 and NFkβ were slightly up regulated whilst LC3B and precursor PARP was up regulated even more (Table 4). At 10 mM NAC, p-AKT and MuC1 were only slightly down regulated. At 50 mM NAC, LC3B, precursors PARP were considerably up regulated whilst caspase 9 and NFkβ were only slightly up regulated. Noticeably, at 50 mM NAC, p-AKT was considerably down regulated. Hence, the survival protein status indicates that both apoptosis and autophagy was taking place, although at lower level, as indicated by the intensity of expression of the survival proteins.
PET cells (NAC with bromelain)
Although, the addition of 25 ug/ml bromelain to 10mM NAC indicated that both apoptosis and autophagy was taking place, the intensity as indicated by the protein markers, was slow when compared to higher concentrations of bromelain such as 50 or 75 ug/ml. With the addition of 50 ug/ml bromelain to NAC, caspase 3, 8, LC3B, cleaved PARP, AKT, Ikkβ and NFkβ were all down regulated, the intensity of up regulation was indicated by the number of arrows (Table 4). At this concentration of bromelain, down regulation is observed in precursor PARP, p-AKT, p-Ikkβ, p-NFkβ with only slight down regulation of MuC1. When 75 ug/ml bromelain was added to 10 mM NAC, a similar effect to that of addition of 50 ug/ml bromelain took place, although at a higher scale. The expression of cellular protein markers indicate that both autophagy and apoptosis was taking place with higher intensity at 50 and 75 ug/ml bromelain.
Hence, based on the cellular proteins that has been detected, both autophagic and apoptotic cell death was induced by both NAC and the combination of NAC with bromelain. Autophagy seems to be less pronounced in YOU cells compared to PET cells indicating that although YOU cells were more receptive to the treatment, death was mainly by apoptosis. Conversely, PET cells was slightly less receptive to this treatment, however, cell death seems to be taking place by both the pathways with perhaps a little more through autophagy compared to YOU cells.
The anti tumour properties of bromelain has been demonstrated in both in vitro and in vivo studies [28-30,41,42] and recent review by Chobotova et al. [33] has emphasized the proteolytic component of bromelain as being the main agent responsible for anti-cancer properties. On the other hand, N-Acetyl cysteine (NAC) is an agent that had been used as a mucolytic [38,39] with the ability to reduce disulphide bonds in polymerized glycoproteins [40]. Since, chemoresistant MPM cells PET and YOU express high levels of MuC1 (glycoprotein) and its chemoresistivity be linked to MuC1, [9-11] we set out to investigate how the combination of two agents that disintegrate glycoprotein would affect the cell viability of MPM cells. Further, numerous oncogenic cellular proteins are all glycoproteins [43-46] and hence the presence of bromelain and NAC may also affect the tumurogenic performance of these molecules.
Bromelain as a single agent has a dramatic effect on cell viability of both YOU and PET cells. There was a linear relationship between bromelain concentration and cell death in both the cell lines, with an IC50 of 54 ug/ml bromelain. However, the cytotoxicity seems to be enhanced by the addition of NAC to bromelain, as shown by decreasing values in IC50 (50% inhibition of cell viability) with additions of various quantities of NAC, (Table 3). Further, the maximum inhibition of cell viability (I max) took place at the higher end of bromelain and NAC concentrations. (25 mM NAC + 75 ug/ml bromelain; 50 mM NAC + 75 ug/ml bromelain) for YOU cells. Likewise a similar situation existed for PET cells, although the I max was slightly smaller compared to YOU cells (a difference of 97 - 88 = 9%) at the higher end of bromelain and NAC concentration. This difference may be due to the differential expression of MuC1, PET cells having a heavier secretion compared to YOU cells, as detected by immunofluorescence microscopy in previous study (Chapter 9).
On a comparative basis, the addition ≥ 5mM NAC to bromelain accelerated the cytotoxic efficacy of bromelain for YOU cells, whilst ≥ 10 mM NAC was required to show a similar relationship. This again indicates that PET cells were more resistant to this form of treatment. Hence, this investigation seems to suggest that the combination of bromelain and NAC may be used for treating chemoresistant MPM cells. Since YOU cells reacted favourably with 97 % response, a higher bromelain concentration >75 ug/ml in combination with NAC may achieve a 100 cell kill. This needs further investigation. Similarly for PET cells, a similar investigation has to be carried out.
Since, cisplatin and 5-Fu are clinically used for treating MPM, we then investigated to determine whether the addition of bromelain and NAC would enhance the cytotoxicity of these agents. The addition of cisplatin (0 - 0.01 uM) as a single agent showed minimal efficacy on YOU cells (<20%) whilst it had no effect on PET cells.
The addition of cisplatin (0.01 uM) to bromelain or NAC depressed the cell inhibitory effect as compared to individual agents (bromelain or NAC) and on the whole did not show any increase in efficacy. However, the addition of dual agents to cisplatin increased the cytotoxicity at 0.01 uM cisplatin and remained constant at other cisplatin concentrations. At 10 mM NAC + 75 ug/ml bromelain, there was no difference in cytotoxicity by addition of cisplatin. A similar scenario was seen with PET cells, although the maximum inhibition of cell viability (cytotoxicity) was less (<80 %) as compared to YOU cells (95%). Hence, compared to bromelain + NAC as dual agents, the addition of cisplatin seems to depress the cytotoxicity (80 vs. 88 % for PET cells; 97 vs. 95 % for YOU cells).
The anti-tumour effect of cisplatin is initiated at molecular level through cross linking of DNA, binding of cisplatin to macromolecules, nucleic acid and plasma membrane [47]. Further it also depletes the methionine pool within the cells by inhibiting methionine uptake [48]. Hence, agents such as NAC or bromelain with radical scavenging properties may interfere with the cytotoxicity mechanism of cisplatin or vice versa. Therefore, the addition of cisplatin to NAC may have depleted the level of NAC slightly, with attenuated efficacy. This indicated that cisplatin was not compatible with NAC. A similar effect was shown with bromelain, although the depression of inhibition was attenuated at higher concentrations of bromelain (75 ug/ml). In other words, the cytotoxicity of bromelain or NAC may be restored at higher concentrations of both the agents which indicate that cisplatin radicals are quenched but at the same time there was sufficient bromelain and NAC to perform its cytotoxic action. This again indicates that cisplatin had lesser effect in this triple combination therapy and no role at higher bromelain and NAC concentrations.
5-Fu as a single agent has minimal efficacy as a cytotoxic agent at the current concentrations investigated (0 - 0.1 uM). The addition of 5-Fu to either bromelain or NAC seems to depress the inhibitory action of bromelain or NAC in YOU cells, indicating that 5-Fu was not compatible with either of these agents, individually. However, the addition of 5-Fu to dual agents (bromelain + NAC) seems to enhance the cytotoxicity above that was shown by the dual agents, without 5-Fu. The greatest enhancement was seen with the addition of 5 mM NAC with 50 ug/ml bromelain. The enhancement of cytotoxicity persisted at all the other triple combinations, although at a lower degree. At 10 mM NAC + 75 ug/ml bromelain, there was almost no enhancement of cytotoxicity. Hence, at higher concentrations of NAC and bromelain, 5-Fu had no effect.
5-Fluorouracil (5-Fu) was metabolized within the cell to 5-fluorodeoxyuridene monophosphate (FduMP) that can covalently bind to thymidate synthetase (dTMP) in the presence of 5,10-methyltetrahydrofolate (CH2-H4 foliate) and inhibit DNA synthesis, hence depleting the cells of dTMP. 5-Fu can also be metabolized to 5-fluorouridine triphosphate (FuTP) which can be incorporated into RNA. One or both of these metabolites account for the antineoplastic activity of 5-Fu [48]. Since the mechanistic function of 5-Fu depends on charge carrying molecules (reactive radicals) that bind to RNA and DNA, the inhibitory action of 5-Fu on either bromelain or NAC can be accounted on the basis that the level of bromelain and NAC may be depleted during this process i.e., bromelain and NAC may be acting as antioxidant molecules.
A similar scenario existed with PET cells, although at 5 mM NAC + 50 ug/ml bromelain, there was a great boost in cytotoxic efficacy (approx 24%),on addition of 0.01 mM 5-Fu. At all other combinations, of triple agents there was an initial drop in efficacy at 0.01 5-Fu which did not attenuate at higher concentrations of 5-Fu. This indicates that 5-Fu shows increase in efficacy only when added to 5 mM NAC + 50 ug/ml bromelain).
When we examined the cytotoxicity of individual agents with 5-Fu, the addition of 5 mM NAC showed a maximum efficacy of <20 % at 0.1 uM 5-Fu. 50 ug/ml bromelain as a single agent with 5-Fu indicated no increase in cytotoxicity compared to bromelain alone. However, the combination of the two agents seems to work in an additive manner. This behaviour of the agents needs further investigation.
Finally, we examined the status of cellular proteins to determine which pathway of cell death is dominant in the two different treatment regimes (NAC alone and in combination with bromelain). NAC treatment on YOU cells seems to indicate that the main pathway of cell death is through apoptosis with a minor component through autophagy. With the combination of NAC and bromelain, there is an indication that death is mainly through apoptosis. In the case of PET cells, treating with NAC alone seems to suggest that apoptotic cell death is very minimal and probably autophagy is slightly more prominent. In combination treatment of NAC and bromelain, cell death was taking place through both apoptosis and autophagy. There is sufficient literature to indicate that bromelain may induce cell death through autophagy and apoptosis, depending on cell types [31,49]. However, cell death mechanism induced by NAC has not been clearly defined, although our initial work seems to suggest that both autophagy and apoptosis may be taking place at low levels with higher concentrations of NAC (10 - 50 mM). NAC has mainly been used for treating aminoacetophen overdose [37], its mechanistic function works through regeneration of glutathione (GSH) which deactivates reactive quinones and hydroquinones that bind to RNA and DNA of hepatocytes [50]. Indeed, numerous studies exists that support NAC as an agent that protects cells against oxidative injury [51,52] and cell death.
There are also studies indicating that NAC was able to decrease oxidative stress induced by doxorubicin while still preserving doxorubicin efficacy as a chemotherapeutic agent. Other studies seem to indicate that NAC blocks cisplatin induced apoptosis through caspase signalling pathways. By virtue of its antioxidant properties, NAC has the ability to quench reactive radicals. Hence, any anti-cancer agent that works through radical induced injury may be prone to reduced efficacy, when NAC is present. However, in the present case the cytotoxic efficacies of both cisplatin and 5-Fu has been reduced in the presence of NAC or bromelain, when present individually, in both the cell lines, indicating that these agents may be interfering with the radical induced injury to the cells that is inflicted by the chemotherapeutic agents. In the case of NAC, several studies support its protective role against radical induced injury. However, in the case of bromelain, its protective role or its role in the modulation of chemotherapeutic drugs has never been investigated before. Our studies seem to indicate that bromelain may have some antioxidant properties that may interfere with the performance of these two chemotherapeutic drugs.
Although, bromelain and NAC as single agents seems to attenuate the performance of cisplatin and 5-Fu, combination of these two agents with cisplatin or 5-Fu seems to indicate that at lower concentrations of the two agents, in combination, there may be an improvement in efficacy, particularly for YOU cells that have a lower level of MuC1. For PET cells, only a single combination of NAC (5mM) + bromelain (50 ug/ml) seems to show cell inhibition that was almost close to 100%. In conclusion, the combination of bromelain and NAC, particularly at the higher end of concentrations (10 uM NAC + 75 ug/ml), there was no pharmaceutical role for cisplatin or 5-Fu in YOU cells. On the contrary, in PET cells, at all concentrations of bromelain + NAC, except at 5 mM NAC + 50 ug/ml bromelain with 5Fu, there was no pharmaceutical role for either cisplatin or 5-Fu. Further studies are necessary to understand why PET cells that express high levels of MuC1 respond better to a particular concentrations of bromelain + NAC.
For loco-regional treatment of MPM, the combination of NAC and bromelain may provide a new form of therapy, without the use of cytotoxic agents. In combination with cytotoxic agents, it may be an important strategy to use a suitable proportion of bromelain and NAC to derive maximum cytotoxicity. However, this needs further studies.