Journal of Glycobiology
Open Access
ISSN: 2168-958X
ISSN: 2168-958X
Review Article - (2014) Volume 3, Issue 1
Keywords: Proteoglycan; Heparan sulfate; Glycosaminoglycan; Cancer
Proteoglycans (PGs) are a diverse group of glycoconjugates composed of different core proteins covalently linked to linear chains of anionic polysaccharides called glycosaminoglycans (GAGs) which consist of repeating disaccharides. Heparan sulfate proteoglycans (HSPGs) comprise a specific small group of proteins with HS GAG chains. There are three known subfamilies of HSPGs: the glycophosphatidylinositol (GPI)-linked PGs called glypicans (GPCs); the membrane-spanning PGs known as syndecans (SDCs); and the secreted extracellular matrix (ECM) proteoglycans agrin, collagen XVIII and perlecan (PRCAN). Hematopoietic cells also contain a secretory vesicle proteoglycan called serglycin. Additionally, a few more proteins such as betaglycan and CD44v3 may appear as part-time HSPGs. In addition, some HSPGs can also have small proportions of chains of other types of GAGs attached to them [1].
HSPGs are structurally very complex, which means they possess large amounts of biological information, allowing them to participate in many different biological and pathological functions. This information is established by a complex synthetic machinery encoded by around fifty different genes which are able to generate huge volumes of molecular combinations [2]. While the exact nature of most of this biological information remains unclear, HS chains are known to be capable of assuming precise biological roles. As a result many functions have been attributed to HSPGs in both the cell membrane and ECM [3], including cell adhesion and migration, organization of the ECM, regulation of proliferation, differentiation and morphogenesis, cytoskeleton organization, tissue repair, inflammation, vascularization and cancer metastasis[3]. They can bind cytokines, chemokines and growth factors and provide them protection from proteolytic degradation, enhance formation of receptor–ligand signaling complexes and direct ligands into the cell for degradation or recycling [3]. This capacity of HSPGs to interact with both soluble ligands and the matrix architecture defines an exclusive combination of properties that enables normal cells to exert an influence on their microenvironment. Indeed, cancer cells employ various mechanisms to exploit these properties and gain a survival advantage, through tissue invasion and metastatic capacity.
HS biosynthesis is not a random process, depending largely on transcriptional and translational controls that regulate the expression of the enzymes involved [4]. Furthermore, post-translational modifications of HSPGs can occur under the influence of other enzymes, especially extracellular sulfatases, heparanase (HPSE) and certain other sheddases [5] (Figure 1).
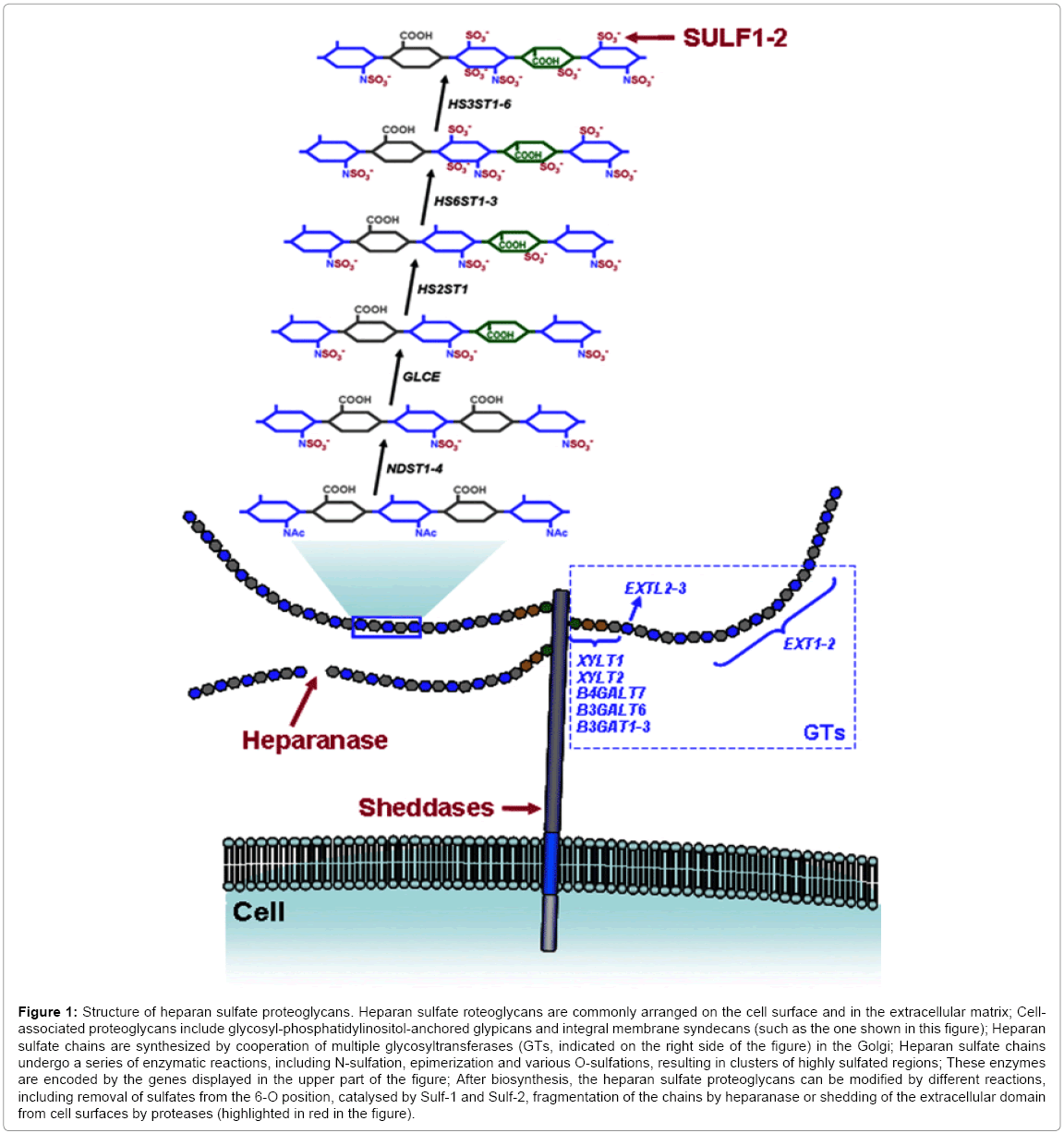
Figure 1: Structure of heparan sulfate proteoglycans. Heparan sulfate roteoglycans are commonly arranged on the cell surface and in the extracellular matrix; Cellassociated proteoglycans include glycosyl phosphatidylinositol-anchored glypicans and integral membrane syndecans (such as the one shown in this figure); Heparan sulfate chains are synthesized by cooperation of multiple glycosyltransferases (GTs, indicated on the right side of the figure) in the Golgi; Heparan sulfate chains undergo a series of enzymatic reactions, including N-sulfation, epimerization and various O-sulfations, resulting in clusters of highly sulfated regions; These enzymes are encoded by the genes displayed in the upper part of the figure; After biosynthesis, the heparan sulfate proteoglycans can be modified by different reactions, including removal of sulfates from the 6-O position, catalysed by Sulf-1 and Sulf-2, fragmentation of the chains by heparanase or shedding of the extracellular domain from cell surfaces by proteases (highlighted in red in the figure).
The aim of this review is to focus on HSPG alterations in cancer from three diverse points of view. We highlight some transcriptional alterations described in different types of cancers for these molecules, followed by a description of some translational aberrations discovered to date, and finally we explain the most frequent post-translations modifications triggered by HSPGs. Every step in the regulation of the synthesis of HSPGs is potentially susceptible to a combinatorial anticancer treatment.
The expression of HSPGs is markedly altered during malignant transformation and tumor progression, affecting both the PG core proteins and the GAG chains [6]. We discuss here three main regulatory points in HSPG formation and modification, all of which are may well be involved in cancer development and progression. We highlight some examples of some of the alterations in the various steps of HSPG synthesis that give rise important changes in cell behavior and malignancy.
Certain well studied genetic or epigenetic modifications are the major cause of protein expression aberrations in cancer. However, some of these alterations can per se induce carcinogenesis while others are simply mere consequences of such malignant processes. The scientific literature reports several examples of gene dysregulations, involving all the enzymatic groups in charge of HSPG biosynthesis (Figure 1). HS chains are synthesized by cooperation of multiple biosynthetic enzymes in the Golgi and attached to specific serine residues at the core protein. Alterations in glycosyltransferases (GTs) involved in HS chain synthesis have been described in some tumors. EXT1and EXT2 encode enzymes involved in HS polymerization, through the consecutive addition of alternating D-glucuronic acid and N-acetyl- D-glucosamine residues; these two genes are well-established tumor suppressors, associated with hereditary multiple exostoses (HME), a disease characterized by the development of benign skeletal tumors in patients [7]. In addition, it has also been suggested that the three EXTlike genes, EXTL1-EXTL3, involved in the transfer of the first N-acetyl- D-glucosamine residue which initiates the polymerization of the HS chains, may each be a tumor suppressor, although none is linked with HME [8].
Although the family of N-deacetylase/N-sulfotransferase enzymes (NDSTs) is essential to generate functional domains in HS chains, descriptions of their altered expression in tumors are scarce, exceptions being the up-regulation of NDST1 and NDST2 in hepatocellular cancer (HCC) [9] and the fact that NDST4 transcripts have been detected in 50% of invasive breast ductal carcinomas [10]. However, a recent study proposed the NDST4 gene as a novel candidate tumor suppressor gene in human cancer, and its loss might be involved in colorectal cancer progression [11].
GLCE encodes a C5-glucuronic acid epimerase, a key enzyme in HS biosynthesis, improving chain flexibility which gives rise to enhanced specific interactions with protein ligands. There are few references concerning alterations of this enzyme in cancer, although it has been pointed out that GLCE may be a potential tumor-suppressor gene, especially in lung cancer [12].
The O-sulphotransferases (HSSTs) group comprises the largest cluster of genes encoding the HS biosynthetic machinery and whose expression is most often deregulated in various tumors, many interesting examples of which have been described. Concerning HS2ST1, studies have demonstrated that it is crucial for cell proliferation and invasion, and its expression correlates with increasing metastatic capacity, especially in prostate cancer [13]. Additionally, overexpression of HS6ST1 in ovarian cancer and HCC, and overexpression of HS6ST2 in colorectal cancer and ovarian cancer has been described [14]. In contrast, HS6STs1-3 are down regulated in glioblastoma [15]. In addition, the largest family of HSSTs corresponds to the 3-O-sulphotransferases, comprising seven different members, all of which are implicated in the formation of specific HS motifs that interact in a selective manner with specific protein ligands. Several studies have described alterations in this group of enzymes in different tumors; overexpression of HS3ST1 in HCC [9] and of HS3ST3A in glioblastoma [15]. On the other hand, many cases of the downregulation of different isoforms have been determined: methylation-associated silencing of HS3ST2 in colon, breast, pancreatic and lung cancers and malignant melanoma17; hypermethylation of HS3ST1, HS3ST2 and HS3ST3A contributes to the invasive phenotype in chondrosarcoma cells [16]. Furthermore, downregulation of isoforms HS3ST4, HS3ST5 and HS3ST6 have been described in invasive breast ductal carcinomas [10].
Genes encoding ECM HS core proteins show dysregulations in some specific tumors, with many important biological consequences. As an example, transcript levels of PRCAN in HCC and lung cancer are practically undetectable, favoring tumor growth and metastatic dissemination [21].
In the last few years, many studies have demonstrated that microRNAs (miRNAs), small noncoding RNAs which regulate fundamental cellular and developmental processes at the posttranscriptional level, can affect the structure and functions of HSPGs, especially in cancer. Increasing numbers of studies have focusing on this issue have definitively explained much of the discord between molecular analysis performed by PCR and tissue analysis performed with immunohistochemistry assays. We describe here some examples where microRNAs can play an important role in HSPG expression in certain cancers. For instance, miR-21 can inhibit HS sulfatase-1 (Sulf- 1) expression in HCC cells, promoting tumor growth [22]. Moreover, as already mentioned, GPC3 is up-regulated in HCC, but in direct association with a downregulation of its repressor miR-1271 [23]. Furthermore, a recent study on preinvasive breast cancer revealed a significant inverse correlation between the expression of SDC1 and the prometastatic miR-10b, suggesting a potential novel mode of posttranscriptional regulation of SDC1 [24]. This finding could explain other results, such as in a recent study on infiltrating ductal adenocarcinomas in breast, where differences between transcript levels of SDC1 quantified by qRT-PCR and immunostaining were determined [10]. In addition, post-transcriptional regulation of syndecan-1 expression in cancer has been indicated, for example in pancreatic cancer [17].
It has been reported that the tissue expression pattern of the NDST isozymes is dependent on their mRNA 5´ untranslated regions (5’ UTRs). These findings demonstrated an important caveat in interpreting levels of enzyme expression from NDST mRNA levels. That is to say that the level of enzyme activity contributed by the various isoforms will depend on both transcription and translation. Furthermore, the same authors also suggested that other enzymes involved in HS formation may also have 5’ UTR-dependent regulation [25].
A recent study of heparanase (HPSE) has identified the existence of the post-transcriptional regulation of the HPSE gene, by a specific 185-bp sequence at the 3’ untranslated region, leading to HPSE downregulation [26]. Furthermore, experiments which deleted this region eliminated the inhibitory effect on HPSE expression.
Once HSPGs are well-configured, they are exported to the cell surface or ECM. Nonetheless, HSPGs can still be modified in the extracellular microenvironment by various reactions that might also be altered in tumoral processes: (1) removal of sulfates from the 6-O position of HS chains by extracellular sulfatases, (2) fragmentation of HS chains by HPSE and (3) shedding of PG extracellular domains from cell surfaces [5] (Figure 1).
Remodeling by extracellular sulfatases
To date, the 6S modification is the only sulfate moiety known to be post-synthetically edited from the HS chains. This reaction is performed by two extracellular isoenzimes that act as endosulfatases (Sulf-1 and Sulf-2) generating specific modifications of the patterns of sulfation on HS chains by removing glucosamine-6S groups from specific regions. Dysregulation of extracellular sulfatases has been described in many tumors [27]. Since 6S modifications are directly linked with the regulation of certain signaling events, the function of these enzymes is to fine-tune the sulfation patterns of HS and thus modulate cell signaling through pathways requiring these coreceptors [2]. Moreover, the Sulfs have been shown to modulate the interaction of a number of protein ligands with HS. Importantly, this property provides a mechanism for Sulf-producing cells to interact with their microenvironment whereby the secreted Sulf causes the release of HSPG-sequestered ligands which can then act back on the cell, and modulate its behavior [27].
Fragmentation of heparan sulfate chains by heparanase
HPSE is an endo-β-D-glucuronidase that is capable of cleaving HS side chains of HSPGs on cell surfaces and the ECM, an activity that is strongly implicated in cancer progression, invasion and tumor metastasis. The HPSE releases fragments that are biologically active [28], influencing the activity and availability of numerous HS-binding factors that regulate cell behavior of both the tumor cells and host cells within the tumor microenvironment [5].
Shedding of proteoglycan extracellular domains
SDCs are, up to the present time, the best studied HSPGs with many cases of increased shedding having been reported in these proteoglycans in cancer. Many sheddases (matrix metalloproteinases) have been described which participate in this process and it can have a dramatic impact on cell behavior, especially on cell adhesion and the formation of growth factor gradients, which is extremely important for cancer progression [29].
Matrix metalloproteinases are able to release intact syndecan ectodomains from the cell surface giving rise to a soluble, shed form of the PG. The shed form of SDC can facilitate the growth, angiogenesis, and metastasis of tumors, playing an active role in the tumoral niche and serum. In fact, studies have demonstrated that high levels of serum SDC1 are an indicator of poor prognosis in both myeloma and lung cancer, and associated with increased growth and metastasis [30]. Moreover, stromal expression of SDC1 in pancreatic cancer is correlated with worse outcomes, as well as being an independent prognostic factor. On the contrary, stromal expression of SDC2 has no significant influence on the prognosis of pancreatic cancer, while epithelial expression is strongly correlated with better outcomes. This fact confirms that in cancer epithelial and stromal SDCs have different functions and that these are related to their locations [31].
Recently it has been discovered that shed SDC1 drives tumor progression by binding to the cell surface and translocating to the nucleus in myeloma cells [32]. Furthermore, new discoveries in HPSE and cancer highlight the fact that tumor expression of HPSE in myeloma and breast cancer cell lines regulates both the location and level of SDC1 within the tumor microenvironment by enhancing its synthesis and subsequent shedding from the tumor cell surface [33]. The HS chains present on syndecan core proteins are by themselves able to suppress the shedding of the proteoglycan [34], such that removing HS chains by HPSE enzymatic activity contributes to increasing levels of the released forms of SDCs. In addition, it is possible that oxidative damage present in the tumoral niche could also contribute significantly to increased levels of shedding in some tumors [34]. Finally, it has been demonstrated that chemotherapy stimulates SDC1 shedding, giving rise to a potentially negative effect of treatment that may promote tumor relapse [35].
HSPGs show a highly modified structure in many cancers, through abnormalities at either the transcriptional, translational or post-translational level. Several examples endorse the fact that these molecules play an important role in carcinogenesis and tumor progression. However, understanding the biosynthetic regulation of HSPGs and decoding the heparanome remain a challenge for scientists in order to fully comprehend the whole process, that is to say, the main functions that HSPGs might actually play in cancer. Once we are able to unravel these as yet unclear roles of HSPGs in cancer, a gate might be opened toward the creation of new anti-cancer drugs.
The University Institute of Oncology of Asturias is supported by Obra Social Cajastur, Asturias, and Spain.