Biochemistry & Pharmacology: Open Access
Open Access
ISSN: 2167-0501
ISSN: 2167-0501
Review Article - (2015) Volume 4, Issue 2
Significant amounts of heavy metals have been added to the soils globally due to the natural as well as anthropogenic activities. The heavy metal accumulation in crops may cause serious disorders in plants as well as in human beings. Therefore, investigation on heavy metals toxicity is remaining an area of scientific interest. In order to increase crop productivity and minimizing health hazards of heavy metals toxicities it is necessary to understand their mechanisms of toxicity. In this way, osmolytes such as proline is endogenous organic substance which regulates normal growth and development of plants, grown under metal contaminated urban and peri-urban areas. Therefore, application of proline and/or manipulation of gene(s) enhance the activity of enzyme(s) and their biosynthetic pathways have also been found to be much effective in mitigating heavy metal toxicity. In this review, impact of heavy metal toxicity on plants and adaptation strategies of plants with special role of proline have briefly been discussed.
<Keywords: Antioxidants; Heavy metal; Oxidative stress; Plant tolerance; Proline
The ever increasing population, urbanization and industrialization have liberated massive volumes of waste water, which is increasingly utilized as a resource for irrigation in urban and peri-urban agricultural areas. Irrigation through this waste water carried a bulk amount of heavy metals and easily entered into the food chain through soil accumulation and subsequent plant uptake. Heavy metal contamination may severely inhibit the plant growth and productivity, and also increase the risk of damage in animal and human health via the biomagnification process [1-4]. Kidneys, bones and liver are the prone sites where heavy metals get stored due to the non-biodegradable and persistent in nature, leading to the malfunctioning of these vital organs and thus cause numerous serious health disorders [5]. A number of heavy metals such as Zn, Cu, Pb, Mn, Ni, Cr, Cd, As are wandering with the waste water and dispersed in the environment for the long time and cause serious threat to different level of food chains [3]. On the one hand, heavy metals cause different disorder in humans and plants while on the other hand among different heavy metals, some are essential for plant growth and development at their concentrations of acceptable limit [6]. The deleterious effects of heavy metals in plants are associated with the inhibition of basic physiological processes including photosynthesis, mineral nutrition and water relation [6-8] (Table 1) and also by the over production of different reactive oxygen species (ROS) [3,6,9]. In order to alleviate the heavy metals toxicities, chemical application and agronomical crop management practices have been used with a little bit of success. In the similar practices, exogenous application of osmolytes such as proline, glycine betaeins, polyamines etc. has emerged an alternative strategy to induce the capability within plants to face successfully the detrimental situation of heavy metals toxicities. Proline is a multifunctional amino acid, behaving like a signaling molecule and initiates the cascade of signaling processes [10]. It regulates the osmotic pressure inside the cell, prevents denaturation of proteins, membrane integrity, stabilization of enzymes, and quenching the toxic ROS [11] (Table 2 and Figure 1). Evidences showed that exogenous application of proline may improve the health of plants against cadmium [12] and selenium [13] (Table 2). Taking together previous studies, it appears that proline plays an important role in enhancing plant stress tolerance and may also help to reduce adverse effects caused by different heavy metals toxicities. Thus, heavy metals exert the adverse pressure from molecular to whole plant level and seriously limit growth and productivity. Therefore, investigation on heavy metals toxicities is remaining an area of scientific interest and further needed to understand their toxicity mechanisms as well as how to minimize health hazards by regulating their entry into crop plants. In this short review, we discuss adaptation strategies of plants with special reference of proline in combating heavy metals toxicities. We also examine recent advances in putative mechanisms whereby proline helps plants to deal with such environmental fluctuations.
| Heavy metal | Plant | Heavy metal induced disturbance in plants | Reference |
|---|---|---|---|
| Arsenic | Carrot, Lettuce, Spinach and Sun flower | Decline in growth, photosynthetic pigments, increased production of stress biomarkers | [7,75] |
| Cadmium | Tobacco | An accumulation of Cd, increased lipid peroxidation and decreased superoxide dismutase and catalase activity | [12] |
| Chromium | Radish | Disrupt the metabolic activity and translocations of nutrients | [16] |
| Nickel | Triticumaestivum | Disturbed the growth and nitrogen metabolism | [76] |
| Lead | Brassica species | Decline in growth due to hyper accumulation | [77] |
Table 1: Effect of heavy metal on plants
| Osmolyte | Plant | Work | Against heavy metal | Reference |
|---|---|---|---|---|
| Proline | Microalgae, Tobacco, Chickpea, Brinjal | Protect membranes, enhance growth, reduce oxidative stress through antioxidant mechanism, provide molecular mechanisms of proline-mediated tolerance | Cadmium | [12, 60, 9, 46] |
| Bean | signaling molecule, redox balancing reduces phytotoxic effects by minimising oxidative stress | Selenium | [13] | |
| Sunflower | Protection by scavenging hydroxyl radical and reducing metal uptake As treatment increased endogenous proline | Arsenic | [7] | |
| Indian senna | involved in stress-resistance mechanisms | Lead | [78] | |
| Ocimumtenuiflorum | Detoxify by mechanism of improve antioxidant system | chromium | [79] | |
| Rice | proline lower water potential and involved in the Hg2+ tolerance | Mercury | [80] | |
| Wheat | Play osmoprotective role and protect plants from Ni stress | Nickel | [81] |
Table 2: Significance of proline in plants under heavy metal stress
A huge amount of heavy metals have been inserted into the environment through both natural as well as anthropogenic activities including the geogenic process, weathering from the parental rocks, frequent irrigation, improper agricultural practices, industrial effluents, refuse burning, organic wastes, transport, combustion of fuels and power generation [14] (Table 3). Environmental contamination by heavy metals is a matter of global concern, which is directly related to anthropogenic actions (Table 3). Industrialization and urbanization have amplified the anthropogenic contribution of heavy metals pollution [2] and their increased level in the biosphere (Table 4). An improper disposal technique of municipal solid waste is well thoughtout one of the major sources of heavy metals in soil [15]. Similar to the municipal solid waste, industrial discharge may also carry a huge amount of heavy metals that exerts pressure on aquatic as well as terrestrial ecosystems including the agricultural fields in adjacent regions of industrial complexes [16].
| Heavy metal | Sources | Reference |
|---|---|---|
| Arsenic | Volcanic eruptions, smelting operations, fossil-fuel combustion and uses of pesticides and herbicides | [3,61] |
| Cadmium | by-product of zinc (occasionally lead) refining | [2 ,62] |
| Nickel | atmospheric fall out, biological cycles, dissolution of rocks and soil, industrial processes, diesel oil and fuel oil, dental or orthopaedic implants, stainless steel kitchen utensils | [63,64] |
| Lead | Industrial application, fuel additive, paint, food, beverages, mining and smelting activities, automobile exhausts | [2,62,65] |
| Chromium | mining and smelting, battery manufacturing, chrome plating, pigments, tanning of animal skins, dyes and wood | [66,67] |
Table 3: Sources of heavy metal in environment
| Heavy metal | Soil | Fresh water | Sea water | Reference |
|---|---|---|---|---|
| Arsenic | 50-15000 (mg kg-1) | 1-10 μg L−1. | > 2 μg L−1 | [68,69,70] |
| Cadmium | 3-6 (mg kg-1) | 0.01 mg L−1 | _ | [71] |
| Nickel | 3-1000 (mg kg-1) | > 10 µg L−1 | 0.1- 0.5 µg L−1 | [63,64,72] |
| Lead | 250-500 (mg kg-1) | 0.1 mg L−1 | _ | [71] |
| Cromium | 0.05-125 (g kg−1) | 0.1-117 μg L−1 | 0.2-0.5 μg L−1 | [73,74] |
Table 4: Concentration of heavy metals in soil and water
Anthropogenic activities may seriously influence the accessibility of heavy metals in the environment. Heavy metals may abruptly damage the vital physiological process of the cells including the gaseous exchange, CO2 fixation, respiration and nutrient absorption potential. A number of biotic as well as abiotic factors such as temperature, soil pH, soil aeration, moisture, type of plants their size and root systems, competition between the plants and the accessibility of elements in soil highly influence metals uptake rates in plants [2,17].
The acidification of rhizosphere and exudation of carboxylates are thought to be potential targets for enhancing metal accumulation in plants. In the process of heavy metal mobilization, primarily a metal has to be captured by root cells and got bound to the outer cell wall. Secondarily, various transporter proteins with their higher affinity to different binding sites help metals uptake across the plasma membrane. Furthermore, the secondary transporters such as channel proteins and/or H+ coupled carrier proteins accelerate the metals uptake and their accumulation. These secondary transporters get energy from the fluctuations of membrane potential of the plasma membrane and root epidermal cells [18]. It has been shown that metal transporters acting as foremost role for the maintaining of intracellular metal homeostasis [19]. At the same time, transporter proteins may also involved in metal detoxification via the regulation of metals uptake from root cells and their translocation from the cytosol to the vacuolar compartment [20] (Figure 1). Moreover, these proteins may also boost the antioxidant system, osmolytes concentration, genetic or ion homeostasis. Similarly, Verbruggen et al. [21] have also reported the influx of arsenate is driven by phosphate transporters while arsenite is taken up by the aquaporin nodulin 26-like intrinsic proteins. From the another point of view, heavy metals such as copper, manganese and zinc etc. act as essential micronutrients for a range of plant physiological processes via the action of Cu, Mn and Zn dependent enzymes. Thus plants have evolved a suite of mechanisms that control and respond to the uptake and accumulation of both essential and non-essential heavy metals [22].
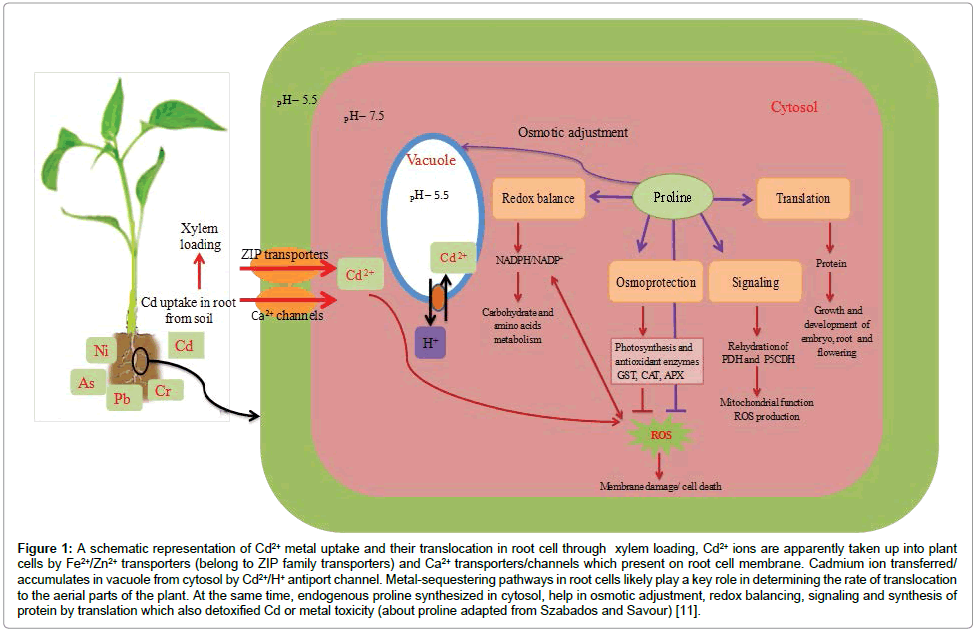
Figure 1: A schematic representation of Cd2+ metal uptake and their translocation in root cell through xylem loading, Cd2+ ions are apparently taken up into plant cells by Fe2+/Zn2+ transporters (belong to ZIP family transporters) and Ca2+ transporters/channels which present on root cell membrane. Cadmium ion transferred/ accumulates in vacuole from cytosol by Cd2+/H+ antiport channel. Metal-sequestering pathways in root cells likely play a key role in determining the rate of translocation to the aerial parts of the plant. At the same time, endogenous proline synthesized in cytosol, help in osmotic adjustment, redox balancing, signaling and synthesis of protein by translation which also detoxified Cd or metal toxicity (about proline adapted from Szabados and Savour) [11].
Heavy metal toxicity is considered one of the major abiotic stresses leading to reduced crop yield and productivity. The higher accumulation of these heavy metals such as As, Cd, Pb, Cr, Zn, Cu and Ni in cells have resulted in different alterations at the physiological, biochemical and cellular level leading to the severe damage to plant [6,9,12,23,24] (Table 1). Panda et al. [25] have reported Cr induced chlorosis in young leaves of wheat, damage to root cells, impaired photosynthesis, altered enzymatic function, stunted growth, and consequently plant death. Similarly, Sangwan et al. [26] observed Cr induced decline in growth of cluster bean due to arrest in activities of nitrogenase, nitrate reductase, nitrite reductase, glutamine synthetase, and glutamate dehydrogenase enzymes. In another finding Yadav et al. [7] have observed the replacement of central atom (Mg) of chlorophyll molecule with As leading to the dismantle of chlorophyll resulting in the breakdown of photosynthesis and growth of the sunflower seedlings under As stress. It is also reported that, arsenic has great potential to block various metabolic processes in cell, and may interact with sulphydryl groups of the proteins, and also replace the phosphate group of ATP. Similar to As, Hg also interacts with the -SH groups and form the S-Hg-S bridge, disrupting the stability of the group and consequently affects seed’s germination and embryo’s growth [27].
Further, study showed that Cr stress declined biomass accumulation in barley plants by causing ultrastructure disorders in leaves such as uneven thickening and swelling of chloroplast, increased amount of plastoglobuli and disintegrated thylakoid membrane which resulted into decline in net stomatal conductance, cellular CO2 concentration, transpiration rate, photochemical efficiency and net photosynthetic rate [8]. In another finding, Hg reduces the plant growth due to its higher affinity to bind with-SH groups of proteins and causes uneven modification in protein structure [23]. It has been observed that brinjal plant when exposed to Cd showed restricted growth, leaf area, photosynthetic pigment and chlorophyll fluorescence [9]. Similarly, Shahid et al. [28] have reported that the growth and photosynthetic pigments of Vicia faba got inhibited under Pb stress. The decrease in growth was associated with the loss of photosynthetic pigments due to modification of chloroplast ultrastructure and higher affinity of Pb for S- and N-ligands of protein, restriction of plastoquinone and carotenoids synthesis pathway, impairment in electron transport chain and membrane permeability, insufficient supply of CO2 and inactivation of numerous enzymes [28,29].
The fate of cells functioning depends on the production of reactive oxygen species (ROS). Over production of ROS than acceptable limit in different compartment of cells and their accumulation may lead damage to various essential macromolecules including peroxidation of lipids and proteins, DNA damage and several other vital constituents of the plants cell (Table 1). Evidence shows that, ROS directly formed through Haber-Weiss reaction lead to the oxidative stress during heavy metal stress [30]. The foremost generation sites of ROS are the chloroplasts, mitochondrion and plasma membrane which are interconnected to electron transport system hence ROS are produced as by products during oxygen metabolism [31]. Keeping the biological significance of heavy metals in mind, they have been classified in two groups namely redox active and redox inactive element [32]. On the one hand, redox active elements may directly participate in redox reaction processes in cells and consequently lead to the formation of ROS such as superoxide radical (O2•¯ ), hydrogen peroxide (H2O2) and after this most lethal hydroxyl radicals (•OH) via the combined Haber-Weiss and Fenton reactions [31] while on the other hand, redox inactive elements may disrupt the electron transport chains and also accelerated the ROS generation inside the cells [32,33]. Extensive studies have been performed on different heavy metals interaction with a wide variety of plants on oxidative stress biomarkers such as Cr in rice seedlings [26], Hg in alfalfa seedlings [23], Zn in wheat plants [6], Cd in brinjal seedlings [9], As in sunflower seedlings [7] and Ni in soybean seedlings [33] (Table 1).
In order to acquire vitamins, minerals and fibers, human beings are dependent on vegetables and fruits as they are the rich sources for them. At the same time, these plants also carry a good amount of toxic metals. It is well established that plants easily take up these metals by absorbing them from contaminated soils as well as from deposits on aerial parts of the plants exposed to the ambient air from polluted environments. Hyper accumulation of heavy metals in plants may cause serious complications in human beings especially of the cardiovascular, renal, nervous, skeletal systems and some other like carcinogenesis, mutagenesis and teratogenesis [34]. Moreover, it has been reported that inorganic form of arsenic (As) is most lethal as it is known as epigenetic carcinogen metalloid. In this way, trivalent arsenite (As+3) has more
properties than the pentavalent arsenate (As+5). In humans, trivalent arsenite (As+3) induces the carcinogenesis in skin, lung, bladder, and kidney tissues by disrupting the multiple signaling pathway [35]. Similarly, the ionic forms of Ni2+ taken up by the cell have great affinity to bind with proteins and amino acids induce carcinogenesis through several processes including DNA hyper methylation (H3K9 mono and dimethylation), DNMT inhibition, DNA mutation, ROS generation, inhibiting histone H2A, H2B, H3 and H4 acetylation, modifying the tumor inhibitor genes to the heterochromatin, and substantial increase of the ubiquitination of H2A and H2B. Moreover, nickel ion also swaps essential metal ions that are generally required for the proper functioning of many enzymes, leading to their dysfunction [36].
Proline has been considered as one of the important osmoticum found in the cellular system exposed to water stress, saline stress, metal stress etc. In recent years, the role of proline has also been characterized as scavenger of ROS, generated during stress condition [37]. Proline synthesization takes place with glutamate involving two consecutive reductions catalyzed by pyrroline-5-carboxylate synthase (P5CS) and pyrroline-5-carboxylate reductase (P5CR), respectively. Another alternative precursor for proline biosynthesis is ornithine (Orn), which can be transaminated to P5C by Orn-d-aminotransferase (OAT), enzyme to be found in mitochondrion however, in animals proline can be synthesized from either arginine or glutamate [38]. The relative contributions of these two precursors to proline synthesis in various tissues have not been explored much and needs further investigation. Moreover, proline may also acts as molecular chaperon stabilizing the structures of protein and regulates the cytosolic pH which helps to balance the redox status of cell. In addition to its role as an osmolyte, proline also prevents the disruption of proteins and membranes by forming clusters with H2O molecules and stabilized their structures [37,39]. Furthermore, in Arabidopsis, the overexpression of an antisense proline dehydrogenase cDNA has been shown to accelerate the proline accumulation and that improves the resistance potential of plant against the freezing and hyper salinity condition [40]. Similarly, Su and Wu [41] showed that the over expression of P5CS gene from mothbean enhanced the accumulation of P5CS mRNA and proline level in transgenic rice which enhances the tolerance capacity of the plant against drought and salt stress. However, in case of human cells proline catabolism is implicated in mitochondria-dependent signaling that regulates the programmed cell death and apoptosis. Evidences showed that, proline application may efficiently remove the toxic ROS in fungi and yeast, thus preventing programmed cell death [42] and also protects human’s cell from carcinogenic oxidative stress [43].
Antioxidant defense system
Plants develop a number of strategies to overcome with the adverse impacts imposed by heavy metals. In plants, heavy metal toxicity may lead to the over production of ROS, resulting in peroxidation of many vital constituents of the cell. In this way, plants have an efficient defense system comprising a set of enzymatic as well as non-enzymatic antioxidants. A wide variety of enzymatic antioxidants consisting of superoxide dismutase (SOD), peroxidase (POD), catalase (CAT) and glutathione-s-transferase (GST) which may efficiently convert the superoxide radicals into hydrogen peroxide and subsequently water and oxygen whereas low molecular weight non-enzymatic antioxidants consisting the proline, ascorbic acid and glutathione may directly detoxify the ROS [7,9,44]. These two groups of antioxidants may professionally quench a wide range of toxic oxygen derivatives and prevent the cells from oxidative stress. Depending upon their localization at the different compartments of the cell their quenching mechanism also differs. In this way, SODs are a group of metaloenzymes that accelerate the conversion of superoxide radical (SOR, O2 •¯) into hydrogen peroxide (H2O2) whereas CAT, guaiacol peroxidase (GPX), and a variety of general PODs catalyze the breakdown of H2O2 [31]. Similarly, GST is capable to catalyze the conjugation of different electrophilic substrates with reduced glutathione. Moreover, several studies report that under stress condition proline acts as an osmolyte and may increase antioxidant enzymes to minimize the adverse effect of oxidative stress [37,45] (Table 2 and Figure 1). In another finding, Islam et al. [12] reported that proline behaves like a plant growth regulators and maintains the osmotic adjustments, protects cells against ROS accumulation under Cd stress. In the similar way, exogenous application of proline reduces phytotoxic effects of selenium by minimizing oxidative stress and also improves growth in Phaseolus vulgaris L. Seedlings [13]. Moreover, Hayat et al. [46] showed that the exogenous application of proline alleviates the damaging effects of Cd in plants and thereby enhances the growth and photosynthesis. At the same time, heavy metal stress cause in a down regulating of leaf water potential on the other side it get recovered by the exogenous application of proline and enhanced the leaf water potential thereby protecting the membranes from metal induced injuries [46,47].
Cellular homeostasis
Proline may accumulate in cytosol under various biotic and abiotic stress conditions [37,45,48]. Hayat et al. [46] observed that the exogenous application of proline may enhance the endogenous proline level under heavy metal stress conditions which helps to maintain intracellular redox homeostasis potential [49] (Figure 1) protects enzymes [12], 3-D structure of proteins [50] and vital organelles including the cell membranes and also reducing the risk of peroxidation of lipids and proteins [47]. Proline may enhance the tolerance potential of plants by chelating heavy metals in the cytoplasm [51], regulating the water potential which is often impaired by heavy metals [52] maintain osmotic adjustment through cellular homeostasis and reduce metal uptake [53]. Wu et al. [54] also observed the minimum efflux of K ions in Anacystis nidulans after the application of proline under Cu stress suggesting the protective role of proline which protects the plasma membrane from Cu toxicity.
Role of genes in metal uptake and their transportation
a number of genes may express under the heavy metal stress which activate the specific enzymes to overcome with negative response. It has been reported that the genetically modified tobacco callus showed more resistance to methyl mercury (CH3Hg+) than the wild-type one. The merB gene encoded the MerB enzyme that dissociates the CH3Hg+ to Hg2+ which is less toxic and accumulated in the form of Hg-polyP complex in tobacco cells [55]. Moreover, the over expression of AtPCS1 and CePCS genes also enhanced the detoxification potential of tobacco plants under As and Cd stress with the increase in phytochelatins (PC) level [56,57]. In another study, Clemens [58] showed that Cd ions are apparently entered into the plant cell by Fe2+/ Zn2+ transporters of ZIP family and possibly by Ca2+ transporters or channels (Figure 1). Furthermore, there are also some genes which expressed in hyper accumulator phenotypes such as HMA4 gene under heavy metal stress. At the same time, some orthologous genes are also consistently over-expressed in Cd and Zn hypertolerant accessions in non-hyper accumulator metallophytes, e.g. SvHMA4 and SpHMA4 in Silene vulgaris and S. paradoxa (Arnetoli and Schat unpublished data). Likewise, MT2b-orthologous genes are over-expressed in hyper accumulators, particularly in metallicolous accessions [59]. In this way, proline has recently been demonstrated to enhance the tolerance capacity of transgenic alga Chlamydomonas reinhardtii under hyper dose of cadmium [60]. In this study, a gene encoding moth bean D1-pyrroline-5-carboxylate synthetase (P5CS), initiates the proline synthesis, and introduced into the nuclear genome of green microalga Chlamydomonas reinhardtii. The transgenic alga produced 80% higher proline levels than the wild-type cells and grew more rapidly under higher Cd concentrations [60].
The above study concludes that ROS are formed as by products during normal metabolic processes in aerobic life and involved in regulation of many physiological processes, however, their enhanced levels have been responsible for many complications in plants. This situation is further get more worsened with the increasing metal pollution due to the enhanced anthropogenic activities. The direct effect of heavy metal may be appeared as over generation of ROS in stressed plant which subsequently cause damage to essential macromolecules such as lipids, proteins, and nucleic acids, and thus, constrain crop productivity. Therefore, to engineer more metal tolerant plants, it is important to find out the key components of the plant metal tolerance network. These components include various genes, transcription factors, proteins and metabolites (osmolytes, phytohormones, lipids, etc.) which may be used to engineer plants for their increased metal tolerance. Despite the availability of ample literature, the understanding of relationship between metal stress and proline interaction at the physiological, biochemical as well as molecular level are still an area of scientific thrust.
Authors are grateful to the UGC for providing financial support. One of the authors Jitendra Kumar is also thankful to UGC for providing financial assistance as JRF under the Rajiv Gandhi National Fellowship scheme (RGNF-2012-13-SCUTT- 33185)