PMC/PubMed Indexed Articles
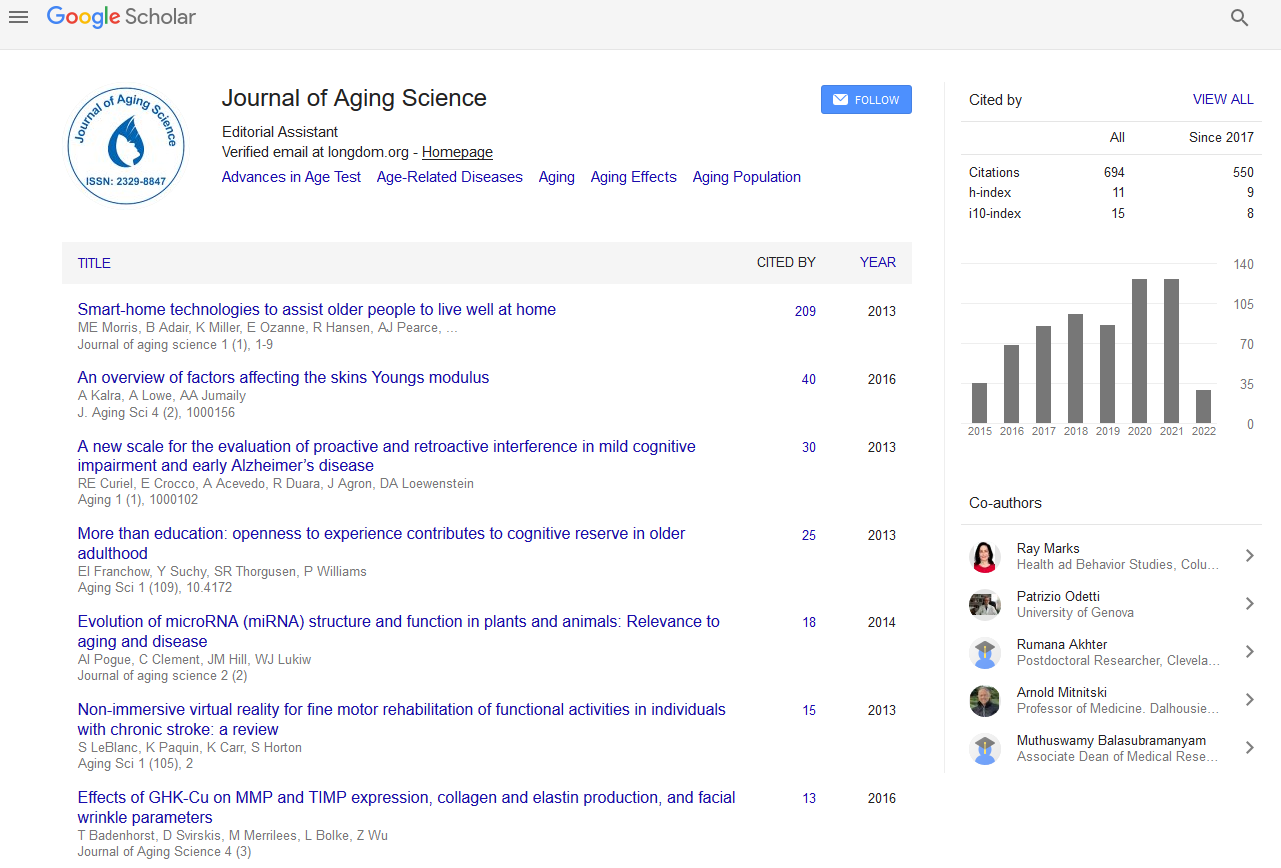

Indexed In
- Open J Gate
- Academic Keys
- JournalTOCs
- ResearchBible
- RefSeek
- Hamdard University
- EBSCO A-Z
- OCLC- WorldCat
- Publons
- Geneva Foundation for Medical Education and Research
- Euro Pub
- Google Scholar
Useful Links
Share This Page
Journal Flyer

Open Access Journals
- Agri and Aquaculture
- Biochemistry
- Bioinformatics & Systems Biology
- Business & Management
- Chemistry
- Clinical Sciences
- Engineering
- Food & Nutrition
- General Science
- Genetics & Molecular Biology
- Immunology & Microbiology
- Medical Sciences
- Neuroscience & Psychology
- Nursing & Health Care
- Pharmaceutical Sciences
Review - (2019) Volume 7, Issue 3
Vulnerability of Human Erythrocytes to Persistent High Glycemic Index Diets: Implications for Ageing and Neurodegeneration: Possible Amelioration by Carnosine
Alan R Hipkiss*Received: 05-Nov-2019 Published: 05-Dec-2019
Abstract
It is proposed that excessive consumption of high carbohydrate and high glycemic index (GI) diets, typical of the socalled Western diet, converts human erythrocytes into systemic sources of methylglyoxal (MG) and glycated protein, including alpha-synuclein. This is due to activity-induced deamidation of asparagine residues in the glycolytic enzyme triosephosphate isomerase (TPI), which can result in loss of enzyme activity and accumulation of the MG precursor dihydroxyacetone-phosphate. Under such circumstances, erythrocytic MG could provoke protein glycation in the tissues, including the brain, and may be responsible for much age-associated macromolecular modification. The naturally-occurring and pluripotent dipeptide carnosine (beta-alanyl-L-histidine) is enriched in erythrocytes (10-fold compared to sera). Carnosine could help to ameliorate MG generation and reactivity, due to its ability to (i) partially inhibit glycolysis and suppress MG generation and (ii) prevent MG-induced protein glycation, It is concluded that persistent consumption of high GI diets should be avoided, and carnosine, administered orally or intra-nasally to enhance access to the brain, could be explored with respect to age-related conditions including type-2 diabetes and neurodegeneration.
Keywords
Age-related conditions; Neurodegeneration; Human erythrocytes; Carnosine
Introduction
It is clear that changes in cellular energy metabolism play important and even causal roles in onset and progression of aging; excessive glycolysis appears to accelerate aging onset, whereas upregulation of mitochondrial ATP generation appears to be somewhat protective towards age-related dysfunction [1-3]. The fact that human erythrocytes are solely glycolytic raises questions concerning their possible causal role towards aging generally and neurodegeneration in particular. A typical characteristic of cellular aging and associated dysfunction is the accumulation of altered protein, obvious pathological examples being amyloid peptide plaques and Lewy bodies in Alzheimer’s disease (AD) and Parkinson’s disease (PD), respectively. Indeed, this suggestion is reinforced by the recent finding that erythrocytes obtained from Alzheimer’s disease patients appear to possess lower levels of certain protective activities (20S proteasome and glyoxalase-1), functions that are usually regarded as exerting anti-aging roles [4]. That erythrocytes may influence aging onset or development is supported by two more recent papers which discuss (i) the causal effects of glycemic index (GI) and load with respect to type-2 diabetes in humans [5], and (ii) age-related changes in mouse erythrocyte metabolism following parabiosis [6]. Two other papers have recently explored the potential efficacy of the endogenous dipeptide carnosine (betaalanyl- L-histidine) towards brain disorders [7] and the effects of carnosine supplementation on cognitive impairment in human subjects [8].
The objectives of the present communication are threefold. First, to suggest that high GI diets in humans induce erythrocytes to perform almost continuous glycolysis, a likely consequence of which is enhanced production of the reactive bicarbonyl methylglyoxal (MG); MG has been suggested to play a causal role in much age-related dysfunction including type-2 diabetes and cognitive dysfunction [9,10] via glycation of proteins, aminolipids and nucleic acid [11,12]. Secondly, to suggest that high GI diets may convert erythrocytes into systemic sources of MG and macromolecular modification throughout the body. Thirdly, to propose that carnosine’s pluripotent properties towards energy metabolism and proteostasis [13] can exert ameliorative activity with respect to MG generation and/or its reactivity, and, under certain circumstances, may help to suppress age-related dysfunction.
Glycolysis And Age-Related Macromolecular Modification
Much age-related modifications of proteins, lipids and nucleic acids mostly occur following generation of highly-reactive products and intermediates of normal metabolism. While the productions of reactive oxygen species (ROS) has been regarded as a consequence of mitochondrial metabolism and have long been regarded as causal to much age-related dysfunction, it is important to note that glycolysis can directly generate dihydroxyacetone-phosphate (DHAP) and glyceraldehyde-3- phosphate (G3P), highly reactive intermediates that can directly modify (glycate) proteins. Furthermore, both these compounds can spontaneously decompose into methylglyoxal (MG) which is even more reactive [10]. Indeed glycolytically–generated MG is regarded as a major source of protein glycation associated with not only aging but much of the protein modifications (secondary complications) which accompany type-2 diabetes [11]. It is also important to note that many of the processes or agents which delay aging onset have a common characteristic, namely partial suppression of carbohydrate catabolism which decreases glycolytic flux [1-3,12,13].
In contrast, two recent studies have shown that the age-related neurodegenerative disease, AD, is associated with lowered glycolytic function [14,15], a conclusion which appears to contradict the notion that age-related dysfunction is a consequence of glycolytic excess. It is, however, possible to reconcile this apparent dilemma by suggesting that dietaryinduced excessive glycolysis decreases the activity of the glycolytic enzyme triosephosphate isomerase (TPI) in either or both erythrocytes and astrocytes. Because TPI catalyses the conversion of DHAP into G3P, loss or decline in its activity will cause DHAP to accumulate and enhance MG formation and consequential protein glycation [16]; indeed much evidence shows a strong associated between protein glycation and AD pathology [17,18].
Glycolysis in erythrocytes: Triose-phosphate isomerase inactivation
The process of glycolysis involves the release of energy, as ATP, mediated by the step-wise conversion of glucose to lactic acid. One of the steps is carried out by the enzyme triose-phosphate isomerase (TPI). TPI catalyses the conversion of DHAP into G3P, both of which are generated by the action of aldolase on fructose-1.6-diphosphate. However TPI is not a true catalyst because its structure can become altered as result of its catalytic activity via a process which has been termed “molecular wear and tear” [19]. Some 30 years ago it was found by Gracy et al. [20] that, as a consequence of its catalytic action, certain asparagine residues (15 and 71) in TPI spontaneously deamidate into aspartic acid residues. It was concluded that “ the probability of deamidation of an individual TPI molecule is a function of the number of times that it is used as a catalyst” [21]. A consequence of such deamidation is the disassociated of the dimeric enzyme into monomers and their subsequent proteolytic destruction. As erythrocytes are unable to synthesize proteins, due to lack of nuclei and ribosomes, it is obvious that should TPI activity become rate-limiting, then MG generation from the accumulated DHAP would accelerate [22]. However, TPI activity in erythrocytes is reported to be more than 3-times that of any other glycolytic enzyme [23], which suggests an evolutionary adaptation to prevent TPI becoming rate-limiting during the normal (limited) lifespan of the human erythrocyte.
It is reasonable to suggest that the human diet has radically changed from that of the “hunter-gatherer” during which much of humankind’s evolution occurred. The modern “Western” diet contains a much higher amount of carbohydrate than that of preceding generations, and almost certainly is very different indeed from that of the hunter-gatherer in terms of carbohydrate intake. Consequently, it is suggested that the high GI and load of the current “ Western ” diet can induce a decline in erythrocytic TPI (as outlined above) which in turn provokes an increase in MG formation. Thus it is suggested that erythrocyte TPI maybe a metabolic “Achilles Heel” when presented with the current high carbohydrate “ Western ” diet. Although the presence of erythrocyte glyoxalase activity would be expected to facilitate MG detoxification, this enzyme can become inactivated by excess MG and furthermore has been found to decline in aged erythrocytes [24].
Erythrocytes and methylglyoxal
As outlined above, it is likely that diets of high GI and almost continuous carbohydrate load can provoke MG accumulation within erythrocytes in the modern human consuming an overabundance of carbohydrate. One possible consequence could be erythrocyte-mediated MG distribution throughout the body; indeed as the erythrocyte membrane is permeable to MG then it likely that the highly glycolytic erythrocyte is a “Trojan Horse” supplying the deleterious glycating agent to many tissues including the brain [16]. Furthermore, MG can glycate proteins within the erythrocyte and thus contribute to the glycotoxin burden, in addition to the glycated proteins in the diet [25] and those generated within the gut [26]. Erythrocytes contain the protein alpha-synuclein (Asyn) which is very readily glycated by MG. Aggregated forms of Asyn (called Lewy bodies) are found in the PD brain [27]. As MG can induce eryptosis (red cell lysis) [28], one wonders whether erythrocyte Asyn could contribute to Lewy body formation, especially as there is a strong association between type-2 diabetes and neurodegenerative conditions e.g. AD and PD [29,30]. It is also interesting to note that the neurotoxin ADTIQ (1-acetyl-6,7-dihydroxyl-1,2,3,4- tetrahydroisoquinoline) has been detected in the brains of type-2 diabetics and PD patients [31]: ADTIQ is the spontaneous product of the reaction between dopamine and MG [32], an observation which reinforces the notion that increased levels of MG contribute to both these pathologies.
During construction of this review, new evidence emerged which supports the proposal that changes in erythrocytic metabolism accompany neurodegenerative condition; Lv et al. reported that erythrocytes from AD patients possessed decreased glyoxalase activity, whilst papers by Graham et al. and Tian et al. showed that erythrocytes from dementia subjects contained increased levels of aggregated alpha-synuclein, observations also possibly related to the deceased proteasome activity also reported by Lv et al. [4,33,34]. Furthermore, MG has been recently reported to be causal towards brain senescence in an aging mouse model [35].
Triose-phosphate isomerase, aging and the brain
As noted above, studies have shown that AD is accompanied by glycolytic dysfunction [14,15] coupled with increased MG generation [35-39]. Furthermore it has recently been reported that brain senescence in a mouse aging model is caused by elevated levels of MG [40]. Theoretically, both these observations may derive from either erythrocytic MG [4,33,34,39] or/and, the inactivation of astrocyte TPI and glyceraldehyde-3-phosphate dehydrogenase activities, perhaps driven by not only astrocytic excessive glycolysis but also due to the effects of MG released from erythrocytes.
It is very likely that TPI is present in all cells which undertake either glycolysis or gluconeogenesis. Neurons are not particularly glycolytic as they receive their energy via input of lactate supplied by astrocytes in which glycolysis readily occurs; altered forms of TPI have been detected in aged cells and tissues [40-42], including the brain. There is evidence for the occurrence of asparagine deamidation in human brains [43] and dysfunctional TPI has been detected in the brains of AD patients [44]. Although there are no reports on whether TPI deamidation occurs in the aged human brain, there is evidence suggesting that upregulation of TPI enhances longevity in mice [45]; TPI activity in a senescence-accelerate mouse strain (SAMP3) brain and liver was found to be substantially lower than in the “normal” SAMR mice, while acupuncture improved organism longevity and enhanced tissue TPI activity [45]. Furthermore, treatment of senescent human fibroblasts with gammatocotrienol not only increased in vitro proliferation but also increased TPI expression [46]. While explanation of the beneficial effects resulting from enhanced TPI expression may reside in the possibility outlined above (i.e. decreased MG generation), it has been pointed out that TPI may perform undefined non-metabolic “ moonlighting ” activities [47]. For example, it is possible that TPI could be involved in cell cycle regulation [48], thereby enhancing organism longevity and stress responses, least in Drosphila [49]. As cell cycle dysregulation has been described in Alzheimer ’ s disease [50], it cannot be necessarily assumed that the consequences of TPI decline in the brain can all be explained in terms of MG accumulation and glycation-related phenomena [51]. Nevertheless there is substantial evidence from animal and human studies showing that TPI dysfunction provokes neurological dysfunction, possibly related in part to enhanced MG-induced protein glycation [52,53]. It has, however, also been suggested that the phenotypic effects of lowered TPI might involve a response to protein miss-folding and its consequences [47]. Additionally it should also be noted that enhancement of MG formation has been shown to promote a hormetic response, thereby improving cellular ability to deal with toxic dicarbonyls. [54,55].
Enhanced protein glycation is frequently associated with agerelated neurodegeneration such as AD and PD [9-12,17,18], and the two major proteins which accumulate in the AD brain, betaamyloid and aggregated Tau, can profoundly affect TPI function. Beta-amyloid induces nitration of TPI and decreases the enzyme’s activity and thereby provokes MG accumulation [56]. In contrast, the Tau protein is normally protective towards TPI, but this activity is lost following Tau hyperphosphorylation and aggregation [57]. Both observations seem to point to a pivotal role for TPI in neurodegenerative conditions.
It has recently been reported that a phenylpyrrole fungicide, frequently used on crops worldwide, induces MG production via the agent’s inhibitory effects on TPI [58]. Such a finding may help to explain the apparent association between pesticide exposure and the increased occurrence of neurodegenerative conditions such as PD, and obviously has serious implications for the food industry.
It is likely that enhanced protein glycation plays a major role in PD. The protein Asyn is present in most cells including erythrocytes, and in aggregated form it is a major component of Lewy bodies in the PD brain, and it has recently been shown that glycation of Asyn provokes its binding to and inhibition of glyceraldehyde-3-phosphate dehydrogenase, thereby provoking G3P accumulation and increasing the potential for enhanced MG formation [59].
Carnosine, aging and age-related dysfunction
Carnosine (beta-alanyl-L-histidine) is synthesized by many animals (mammals, birds and fish) and is mainly located in muscle tissue and the brain, especially in the olfactory lobe of mammals [7,60-62]. It is likely that carnosine is present in most carnivorous diets; there is only one report of carnosine occurring in plant tissue (gourd skin) [63]. Originally discovered over 100 years ago, many properties have been ascribed to the dipeptide. These included wound healing agent [64,65], immune-stimulant [66,67] and anti-aging agent [68,69], while at the biochemical level, pH buffering [70,71], anti-oxidant [72,73] and anti-glycating [74-77] activities have been detected. More recently, beneficial effects of the dipeptide on lung disease [78,79], various brain disorders [80-82] and a variety of conditions related to diabetes such as diabetic kidney disease [83-85], cardiovascular disease [86,87] and stroke [88,89] have been observed. Anti-viral activity towards Denge and Zika viruses has also been detected. [90]. Toxicity towards tumour cell growth was first observed some decades ago [91,92] and whilst it appears that the dipeptide’s effect is associated with decreased glycolytic ATP generation [93,94], there is no agreement on the precise mechanisms responsible [95-97]. Another recent finding is that carnosine induces intestinal cells to release exosomes that activate neuronal cells [98].
As outlined above, it is likely that MG formation and detoxification can strongly influence age-related macromolecular modification. Although model systems have demonstrated that carnosine can react with MG [99], there is little in vivo evidence for occurrence of the putative reaction products i.e. carnosine- MG adducts. In fact, the major carnosine-adduct found in human urine is that formed between the dipeptide and acrolein [100-102]. However, acrolein is usually thought to be a product of lipid oxidative damage [103]. Given that carnosine seems to protect against the consequence of excessive carbohydrate catabolism, the lipid origin of acrolein in the carnosine-acrolein adduct may therefore be questionable. As the structures of acrolein and MG are quite similar, the theoretical conversion of MG into acrolein could occur in two steps, first a reduction generating lactaldehyde followed by a dehydration to generate acrolein. Hence one has to consider whether MG is a source of acrolein in vivo. In support of this proposition, a recent study using glyoxalase-knockout mouse cells has demonstrated the presence of an additional/alternative route by which MG is detoxified to lactaldehyde using uncharacterized aldehyde/keto reductases [104]. The conversion of lactaldehyde to acrolein has yet to be demonstrated, however. Nevertheless should this route of MG detoxification be established, it would have the advantage of ensuring the elimination of two molecules of MG per carnosine, as the carnosine-acrolein adduct is a cyclic structure formed by two acrolein molecules per carnosine [102].
It has been shown that there is a strong association between schizophrenia and elevated levels of plasma protein glycation (i.e. carbonyl stress) mediated by triose-phosphates [105]. There are reports showing that dietary supplementation with carnosine can exert beneficial effects towards human schizophrenics [106,107]. Thus, one is tempted to speculate that carnosine’s apparent beneficial effects towards schizophrenia may reside at least in part from the dipeptide’s anti-glycating activity, especially as it has recently been reported that schizophrenia is associated with glial cell glycolytic dysfunction, which produces an advanced aging phenotype and accelerated aging [108].
Carnosine and energy metabolism
The many beneficial effects of carnosine (some of which outlined above) suggest that the dipeptide is pluripotent in its actions. It not only can protect cells against deleterious agents such MG, acrolein and malondialdehyde, but it may directly interfere with glycolysis. However, the dipeptide’s effects can vary depending on cell metabolism. For example, carnosine’s effect on yeast cells varied according to the growth conditions; the dipeptide was somewhat inhibitory to cells growing on glucose as sole carbon source i.e. where glycolysis was the major ATP generating pathway, whereas in cells growing aerobically on glycerol, i.e. where mitochondria provided ATP generation, the dipeptide stimulated growth and exerted no inhibitory effects on cell proliferation [109]. Thus it appears from these yeast studies that carnosine can exert suppressive activity towards glycolysis.
Studies using transformed cells also show that carnosine exerts inhibitory effects upon glycolytic ATP synthesis [93,96]. However, the mechanism(s) by which these effects are mediated remains uncertain: there are at least three possibilities. First, it is possibility that carnosine stimulates the enzyme fructose-1,6- bisphosphatase (FBP). FBP is regarded as a gluconeogenic as it converts fructose-1,6-bisphosphate into fructose-6-phosphate (F6P), but does not generate ATP. Many years ago, it was shown that carnosine and histidine could activate FBP by binding to zinc ions which otherwise inhibit FBP activity [110]. Thus upon FBP activation in yeast or tumor cells, the presence of the dipeptide would provoke a futile ATP-consuming cycle in which fructose-6-phosphate is phosphorylated using ATP, yielding fructose-1,6-diphosphate which then dephosphorylated by FBP back to F6P and inorganic phosphate. A second possibility is that carnosine exerts effects upon the regulatory complex mTOR. mTOR activity is suppressed by the immune-suppressor rapamycin which results in decreased glycolytic activity and upregulation of mitochondrial ATP generation [111]. Rapamycin [112] and carnosine [113] have both been shown to induce similar anti-aging effects such as delaying senescence in cultured human fibroblasts and stimulating proteolysis of long-lived proteins in replicative senescent cells, observations suggesting that carnosine may possess rapamycin-like properties [111]. However, carnosine’s putative rapamycin-like activity has been questioned in studies using various glioblastoma cells which are unresponsive to rapamycin but whose growth is still inhibited by carnosine [114]. The third possible mechanistic route of carnosine’s inhibitory effects of glycolysis involves upregulation of the enzyme pyruvate dehydrogenase kinase-4 (PDK4) [115], whose activity decreases glycolysis. It is interesting to note that aging in cultured mouse oocytes is delayed by the polyamine putrecine which also upregulates PDK4 expression [116]; carnosine has been shown to delay aging in cultured human fibroblasts [117], although whether this is also accompanied by increased PDK4 has yet to be explored. Thus one may conclude that carnosine’s anti-aging activity might explained by its antiglycolytic effects mediated by upregulation of PDK4. However, the various effects on glycolysis induced by the presence of carnosine may not be necessarily mutually exclusive; hence it is suggested that, as carnosine appears to be pluripotent in terms of its effects on glycolysis, any one or more of the possible mechanisms outlined above could be responsible for its suppressive effects on glycolysis.
Carnosine and erythrocytes
There has only been one report of the occurrence of carnosine in human erythrocytes [118]; interestingly, carnosine was shown to be enriched (up to ten-fold) within erythrocytes compared to serum, whereas acetyl-carnosine (unsusceptible to carnosinase attack) was found predominantly in serum. Although these findings have yet to be confirmed, it was also found that when erythrocytes and sera from young (29 years old ± 4) and elderly (81 years ± 7) human subjects were compared, both carnosine and acetyl-carnosine levels were substantially lower in the elderly individuals. The origin of red cell carnosine has yet to be studied; presumably erythrocytic carnosine is synthesized during red cell development in the bone marrow. Hence it is conceivable that raised levels of erythrocyte carnosine could be achieved by dietary supplementation with beta-alanine (or carnosine as a source of beta-alanine), although full benefit might be not be achievable until complete red cell replacement is achieved (i.e. around 120 days, the maximum lifespan of human erythrocytes.). It would also be interesting to determine if high GI diets affect erythrocyte carnosine levels. Whether carnosine-loaded erythrocytes supply the dipeptide to the tissues, including the brain, has yet to be investigated.
Carnosine and neurodegeneration
Theoretically carnosine could exert beneficial effects in brain tissue by (i) decreasing glycolytic flux in astrocytes and glia, (ii) directly facilitating elimination of glycating agents such as MG, or (iii) indirectly, by decreasing erythrocyte-generated MG and/or glycated proteins such as Asyn. Carnosine has been previously proposed as potentially therapeutic towards both AD and PD [119,120], especially if administered nasally to avoid serum carnosinase activity [79]. Recent findings support these suggestions [121,122], including the potential efficacy of the nasal route [123]. It is also interesting to note that the olfactory lobe is enriched in carnosine and loss of a sense of smell is a frequent early symptom of neurodegeneration [124], which, at least superficially, supports the notion that carnosine might play a role in controlling aspects of neurological function[125-127].
Discussion and Conclusion
It is clear that aging onset can be delayed if excessive glycolytic activity is suppressed to a minimum. Biochemically, under such a circumstance, generation of deleterious agents such as MG and its reaction products (AGEs) would be reduced to a minimum. It is likely that the current “Western” diet (almost permanent high GI carbohydrate input) may turn erythrocytes into a major source of systemic of MG and protein-AGEs, thereby accelerating aging onset and age-related dysfunction, partly as a result of the activity-induced change in the primary structure of red cell TPI. Indeed recent studies support the notion that enhanced erythrocytic MG generation plays a role in certain age-related neurodegenerative conditions.
Carnosine’s ability to suppress glycolysis may help to explain its apparent beneficial effects towards cellular aging and tumor cell growth. It is possible that carnosine also exerts anti-glycating activity by scavenging reactive carbonyls such as MG. Indeed carnosine-acrolein adducts can be detected in human urine, which suggest that the dipeptide could be beneficial towards human aging where AGE formation occurs, especially as carnosine is far less toxic than other anti-glycating agents such as aminoguanidine. Thus, increasing synthesis or dietary input of carnosine could be beneficial by suppressing glycation-related, age-related, phenomena.
Green vegetables are, of course, another source of naturallyoccurring anti-glycating agents, due to the synthesis of MG precursors during photosynthesis, against which plants must protect themselves. Indeed there are multiple claims in the literature suggesting that plant preparations possess antiglycating activity. It would be interesting to determine whether increased intake of vegetable-derived anti-glycating agents affects erythrocyte carnosine levels and the excretion of carnosineacrolein adducts.
REFERENCES
- Ingram DK, Roth GS. Glycolytic inhibition as a strategy for developing calorie restriction mimetics. Exp Gerontol. 2011;46(2): 148-154.
- Priebe S, Menzel U, Zarse K, Groth M, Platzer M, et al. Extension of life span by impaired glucose metabolism in Caenorhabditis elegans is accompanied by structural rearrangements of the transcriptomic network. PLoS One. 2013; 8(10): e77776.
- Hipkiss AR. On the mechanisms of ageing suppression by dietary restriction-is persistent glycolysis the problem? Mech Ageing Dev. 2006;127(1): 8-15.
- Lv H, Wei GY, Guo CS, Deng YF, Jiang YM, Gao C, et al. 20S proteasome and glyoxalase 1 activities decrease in erythrocytes derived from Alzheimer's disease patients. Neural Regen Res. 2020;15(1): 178-183.
- Ojo O, Ojo OO, Wang XH, Adegboye ARA. The Effects of a Low GI Diet on Cardiometabolic and Inflammatory Parameters in Patients with Type 2 and Gestational Diabetes: A Systematic Review and Meta-Analysis of Randomised Controlled Trials Nutrients. 2019;11(7): E1584.
- Morrison EJ, Champagne DP, Dzieciatkowska M, Nemkov T, Zimring JC, Hansen KC, et al. Parabiosis incompletely reverses aging-induced metabolic changes and oxidant stress in mouse red blood cells. Nutrients. 2019;11(6): E1337.
- Schön M, Mousa A, Berk M, Chia WL, Ukropec J, Majid A, Ukropcová B, et al. The potential of carnosine in brain-related disorders: A comprehensive review of current evidence. Nutrients. 2019;11(6): E1196.
- Masuoka N, Yoshimine C, Hori M, Tanaka M, Asada T, Abe K, et al. Effects of Anserine/Carnosine Supplementation on Mild Cognitive Impairment with APOE4. Nutrients. 2019;11(7): E1626.
- Beeri MS, Moshier E, Schmeidler J, Godbold J, Uribarri J, Reddy S, et al. Serum concentration of an inflammatory glycotoxin, methylglyoxal, is associated with increased cognitive decline in elderly individuals. Mech Ageing Dev. 2011;132(11): 583-587.
- Huang X, Wang F, Chen W, Chen Y, Wang N, Von Maltzan K. Possible link between the cognitive dysfunction associated with diabetes mellitus and the neurotoxicity of methylglyoxal. Brain Res. 2012;1469: 82-91.
- Nigro C, Leone A, Fiory F, Prevenzano I, Nicolò A, Mirra P, et al. Dicarbonyl Stress at the Crossroads of Healthy and Unhealthy Aging. Cells. 2019;8(7): E749.
- Allaman I, Bélanger M, Magistretti PJ. Methylglyoxal, the dark side of glycolysis. Front Neurosci. 2015;9: 23.
- Moraru A, Wiederstein J, Pfaff D, Fleming T, Miller AK, Nawroth P, et al. Elevated Levels of the Reactive Metabolite Methylglyoxal Recapitulate Progression of Type 2 Diabetes. Cell Metab. 2018:27(4): 926-934.
- Butterfield DA, Halliwell B. Oxidative stress, dysfunctional glucose metabolism and Alzheimer disease. Nat Rev Neurosci. 2019;20: 148-160.
- Theurey P, Connolly NMC, Fortunati I, Basso E, Lauwen S, Ferrante C, et al. Systems biology identifies preserved integrity but impaired metabolism of mitochondria due to a glycolytic defect in Alzheimer's disease neurons. Aging Cell. 2019;18(3): e12924.
- Hipkiss AR. The human erythrocyte can become both a metabolic “Achilles’ Heel” and a “Trojan Horse”: Likely consequences of persistent excessive glycolysis. Integr Food Nutr Metab. 2019;6: 1-2.
- Vicente Miranda H, El-Agnaf OM, Outeiro TF. (2016). Glycation in Parkinson's disease and Alzheimer's disease. Mov Disord. 2016;31(6): 782-790.
- Srikanth V, Westcott B, Forbes J, Phan TG, Beare R, Venn A, et al. Methylglyoxal, cognitive function and cerebral atrophy in older people. J Gerontol A Biol Sci Med Sci. 2013;68(1): 68-73.
- Gracy RW, Talent JM, Zvaigzne AI (1998) Molecular wear and tear leads to terminal marking and the unstable isoforms of aging. J Exp Zool. 1998;282(1): 18-27.
- Sun AQ, Yüksel KU, Gracy RW. Relationship between the catalytic center and the primary degradation site of triosephosphate isomerase: Effects of active site modification and deamidation. Arch Biochem Biophys. 1992;293(2): 382-389.
- Robinson NE, Robinson AB. Triosephosphate Isomerase. In: Molecular clocks: Deamidation of asparaginyl and glutaminyl residues in peptides and proteins. Althouse press, Oregon, USA, 2004;pp:231.
- Hipkiss AR. Energy metabolism and ageing regulation: Metabolically driven deamidation of triosephosphate isomerase may contribute to proteostatic dysfunction. Ageing Res Rev. 10(4): 498-502.
- Newsholme EA, Start C. Regulation in Metabolism. In: Biochemical education. John wiley and sons, London, UK, 1973; pp: 98-99.
- McLellan AC, Thornalley PJ. Glyoxalase activity in human red blood cells fractioned by age. Mech Ageing Dev. 1989;48(1): 63-71.
- Cai W, Uribarri J, Zhu L, Chen X, Swamy S, Zhao Z, et al. Oral glycotoxins are a modifiable cause of dementia and the metabolic syndrome in mice and humans. Proc Natl Acad Sci USA. 2014;111(13): 4940-4905.
- Snelson M, Coughlan MT. Dietary advanced glycation end products: Digestion, metabolism and modulation of gut microbial ecology. Nutrients. 2019;11(2): E215.
- Miranda HV, Szego ÉM, Oliveira LMA, Breda C, Darendelioglu E, de Oliveira RM, et al. Glycation potentiates α-synuclein-associated neurodegeneration in synucleinopathies. Brain. 140(5): 1399-1419.
- Xu W, Peng F, Deng Y, Fan X, Li N. The emerging roles of eryptosis in liver diseases. Transfus Clin Biol. 2019; pii: S1246-7820(19)30062-X.
- Caberlotto L, Nguyen TP, Lauria M, Priami C, Rimondini R, Maioli S, et al. Cross-disease analysis of Alzheimer's disease and type-2 Diabetes highlights the role of autophagy in the pathophysiology of two highly comorbid diseases. Sci Rep. 2019;9: 3965.
- Renaud J, Simola N, Martinoli MG. The sweet road to Parkinson's disease. Aging. 2019;11(3): 853-854.
- Xie B, Lin F, Ullah K, Peng L, Ding W, Dai R, et al. A newly discovered neurotoxin ADTIQ associated with hyperglycemia and Parkinson's disease. Biochem Biophys Res Commun. 2015;459(3): 361-366.
- Szent-Györgyi A, McLaughlin JA. Interaction of glyoxal and methylglyoxal with biogenic amines. Proc Natl Acad Sci USA. 1975;72(4): 1610-1611.
- Graham C, Santiago-Mugica E, Abdel-All Z, Li M, McNally R, Kalaria RN, et al. Erythrocytes as Biomarkers for Dementia: Analysis of Protein Content and Alpha-Synuclein. J Alzheimers Dis. 12. 2019;1(1): 569-580.
- Tian C, Liu G, Gao L, Soltys D, Pan C , Stewart T, et al. Erythrocytic a-synuclein as a potential biomarker for Parkinsons’s disease. Transl Neurodegen. 2019;8: 1-12.
- Li H, Zheng L, Chen C, Liu X, Zhang W. Brain senescence caused by elevated levels of reactive metabolite methylglyoxal on D-galactose–induced aging mice. Front Neurosci. 2019;13: 1004.
- Chou PS, Wu MN, Yang CC, Shen CT Yang YH. Effect of advanced glycation end products on the Progression of Alzheimer's Disease. J Alzheimers Dis. 2019;72(1): 191-197.
- Rorbach-Doplata A, Piwowar A. Neurometabolic Evidence Supporting the Hypothesis of Increased Incidence of Type 3 Diabetes Mellitus in the 21st Century. Biomed Res Int. 2019;21: 1435276.
- Haddad M, Perrotte M, Khedher MRB, Demongin C, Lepage A, Fülöp T, et al. Methylglyoxal and Glyoxal as Potential Peripheral Markers for MCI Diagnosis and their effects on the expression of neurotrophic, inflammatory and neurodegenerative factors in neurons and in neuronal derived-extracellular vesicles. Int J Mol Sci. 2019;20(19): E4906.
- Hayden MR. Type 2 Diabetes Mellitus Increases The Risk of Late-Onset Alzheimer's Disease: Ultrastructural Remodeling of the Neurovascular Unit and Diabetic Gliopathy. Brain Sci. 2019;9(10): E262.
- Prestes AS, Dos Santos MM, Ecker A, Zanini D, Schetinger MR, Rosemberg DB, et al. Evaluation of methylglyoxal toxicity in human erythrocytes, leukocytes and platelets. Toxicol Mech Methods. 2017;27(4): 307-317.
- Dan A, Takahashi M, Masuda-Suzukake M, Kametani F, Nonaka T, Kondo H, et al. Extensive deamidation at asparagine residue 279 accounts for weak immunoreactivity of tau with RD4 antibody in Alzheimer's disease brain. Acta Neuropathol Commun. 2013;1: 54.
- Truscott RJ, Friedrich MG. The etiology of human age-related cataract. Proteins don't last forever. Biochim Biophys Acta. 2016;1860(1 Pt B): 192-198.
- Reissner KJ, Aswad DW. Deamidation and isoaspartate formation in proteins: Unwanted alterations or surreptitious signals? Cell Mol Life Sci. 2003;60(7): 1281-1295.
- Zhao L, Jia Y, Yan D, Zhou C, Han J, Yu J. Aging-related changes of triose phosphate isomerase in hippocampus of senescence accelerated mouse and the intervention of acupuncture. Neurosci Lett. 2013;542: 59-64.
- Tajes M, Guivernau B, Ramos-Fernández E, Bosch-Morató M, Palomer E, Guix FX, et al. The pathophysiology of triose phosphate isomerase dysfunction in Alzheimer's disease. Histol Histopathol. 2013;28(1): 43-51.
- Tan JK, Jaafar F, Makpol S. Proteomic profiling of senescent human diploid fibroblasts treated with gamma-tocotrienol. BMC Complement Altern Med. 2018;18(1): 314.
- Rodríguez-Bolaños M, Perez-Montfort R. Medical and Veterinary Importance of the Moonlighting Functions of Triosephosphate Isomerase. Curr Protein Pept Sci. 2019;20(4): 304-315.
- Lincet H, Icard P. How do glycolytic enzymes favour cancer cell proliferation by nonmetabolic functions? Oncogene. 2015;34(29): 3751-3759.
- Zhou D, Xue J, Lai JC, Schork NJ, White KP, Haddad GG. Mechanisms underlying hypoxia tolerance in Drosophila melanogaster: Hairy as a metabolic switch. PLoS Genet. 2008;4(10): e1000221.
- Majd S, Power J, Majd Z. Alzheimer's disease and cancer: When two monsters cannot be together. Front Neurosci. 2019;13: 155.
- Segal J, Mülleder M, Krüger A, Adler T, Scholze-Wittler M, Becker L, et al. Low catalytic activity is insufficient to induce disease pathology in triosephosphate isomerase deficiency. J Inherit Metab Dis. 2019;42(5): 839-849.
- Roland BP, Richards KR, Hrizo SL, Eicher S, Barile ZJ, Chang TC, et al. Missense variant in TPI1 (Arg189Gln) causes neurologic deficits through structural changes in the triosephosphate isomerase catalytic site and reduced enzyme levels in vivo. Biochim Biophys Acta Mol Basis Dis. 2019;1865(9): 2257-2266.
- Cabrera N, Torres-Larios A, García-Torres I, Enríquez-Flores S, Perez-Montfort R. Differential effects on enzyme stability and kinetic parameters of mutants related to human triosephosphate isomerase deficiency. Biochim Biophys Acta Gen Subj. 2018;1862(6): 1401-1409.
- Ravichandran M, Priebe S, Grigolon G, Rozanov L, Groth M, Laube B, et al. Impairing L-Threonine Catabolism Promotes Healthspan through Methylglyoxal-Mediated Proteohormesis. Cell Metab. 2018;27(4): 914-925.
- Nokin MJ, Durieux F, Bellier J, Peulen O, Uchida K, Spiegel DA, et al. Hormetic potential of methylglyoxal, a side-product of glycolysis, in switching tumours from growth to death. Sci Rep. 2017;7(1): 11722.
- Tajes M, Eraso-Pichot A, Rubio-Moscardó F, Guivernau B, Ramos-Fernández E, Bosch-Morató M, et al. Methylglyoxal produced by amyloid-β peptide-induced nitrotyrosination of triosephosphate isomerase triggers neuronal death in Alzheimer's disease. J Alzheimers Dis. 2014;41(1): 273-288.
- Park SA, Park HW, Kim NH, Kim YH, Kwak MJ, Shin JS, et al. Effects of Tau on the activity of triose phosphate isomerase (TPI) in brain cells. Neurochem Int. 2010;56(8): 886-892.
- Brandhorst TT, Kean IRL, Lawry SM, Wiesner DL, Klein BS. Phenylpyrrole fungicides act on triosephosphate isomerase to induce methylglyoxal stress and alter hybrid histidine kinase activity. Sci Rep. 2019;9(1): 5047.
- Semenyuk P, Barinova K, Muronetz V. Glycation of a-synuclein amplifies the binding with glyceraldehyde-3-phosphate dehydrogenase. Int J Biol. Macromol. 2019;127: 278-285.
- Artioli GG, Sale C, Jones RL (2019). Carnosine in health and disease. Eur J Sport Sci. 2019;19(1): 30-39.
- Boldyrev AA, Aldini G, Derave W. Physiology and pathophysiology of carnosine. Physiol Rev. 2013;93(4): 1803-1845.
- Hipkiss AR, Baye E, de Courten B. Carnosine and the processes of ageing. Maturitas. 2016;93: 28-33.
- Swetha MP, Muthukumar SP. Characterization of nutrients, amino acids, polyphenols and antioxidant activity of Ridge gourd (Luffa acutangula) peel. J Food Sci Technol. 2016;53(7): 3122-3128.
- Mirzakhani N, Farshid AA, Tamaddonfard E, Imani M, Erfanparast A, Noroozinia F. Carnosine improves functional recovery and structural regeneration after sciatic nerve crush injury in rats. Life Sci. 2018;215: 22-30.
- Ansurudeen I, Sunkari VG, Grünler J, Peters V, Schmitt CP, Catrina SB, et al. Carnosine enhances diabetic wound healing in the db/db mouse model of type 2 diabetes. Amino Acids. 2012;43(1): 127-134.
- Hyland P, Duggan O, Hipkiss A, Barnett C, Barnett Y. The effects of carnosine on oxidative DNA damage levels and in vitro lifespan in human peripheral blood derived CD4+T cell clones. Mech Ageing Dev. 2000;121(1): 203-215.
- Li YF, He RR, Tsoi B, Li XD, Li WX, Abe K, et al. Anti-stress effects of carnosine on restraint-evoked immuno-compromise in mice through spleen lymphocyte number maintenance. PLoS One. 2012;7(4): e33190.
- Hipkiss AR. Carnosine, a protective, anti-ageing peptide? Int J Biochem Cell Biol. 1998;30(8): 863-868.
- Boldyrev AA, Gallant SC, Sukhich GT. Carnosine, the protective, anti-aging peptide. Biosci Rep. 1999;19(6): 581-587.
- Matthews JJ, Artioli GG, Turner MD, Sale C. The Physiological Roles of Carnosine and β-Alanine in Exercising Human Skeletal Muscle. Med Sci Sports Exerc. 2019;51(10): 2098-2108.
- Blancquaert L, Everaert I, Derave W. Beta-alanine supplementation, muscle carnosine and exercise performance. Curr Opin Clin Nutr Metab Care. 2015;18(1): 63-70.
- Boldyrev AA. Does carnosine possess direct antioxidant activity? Int J Biochem. 1993;25(8): 1101-1107.
- Ansari FA, Khan AA, Mahmood R. Protective effect of carnosine and N-acetylcysteine against sodium nitrite-induced oxidative stress and DNA damage in rat intestine. Environ Sci Pollut Res Int. 2018;25(20): 19380-19392.
- Yan H, Harding JJ. Carnosine protects against the inactivation of esterase induced by glycation and a steroid. Biochim Biophys Acta. 2005;1741(1): 120-126.
- Narda M, Peno-Mazzarino L, Krutmann J, Trullas C, Granger C. Novel Facial Cream Containing Carnosine Inhibits Formation of Advanced Glycation End-Products in Human Skin. Skin Pharmacol Physiol. 2018;31(6): 324-331.
- Ghodsi R, Kheirouri S. Carnosine and advanced glycation end products: A systematic review. Amino Acids. 20189;50(9): 1177-1186.
- Tanaka KI, Kawahara M. Carnosine and Lung Disease. Curr Med Chem. 2019.
- Sun C, Wu Q, Zhang X, He Q, Zhao H. Mechanistic evaluation of the protective effect of carnosine on acute lung injury in sepsis rats. Pharmacology. 2017;100(5): 292-300.
- Caruso G, Caraci F, Jolivet RB. Pivotal role of carnosine in the modulation of brain cells activity: Multimodal mechanism of action and therapeutic potential in neurodegenerative disorders. Prog Neurobiol. 2019;175: 35-53.
- Berezhnoy DS, Stvolinsky SL, Lopachev AV, Devyatov AA, Lopacheva OM, Kulikova OI, et al. Carnosine as an effective neuro-protector in brain pathology and potential neuromodulator in normal conditions. Amino Acids. 2019;51(1): 139-150.
- Bermúdez ML, Skelton MR, Genter MB. Intranasal carnosine attenuates transcriptomic alterations and improves mitochondrial function in the Thy1-aSyn mouse model of Parkinson's disease. Mol Genet Metab. 2018;125(3): 305-313.
- Corona C, Frazzini V, Silvestri E, Lattanzio R, La Sorda R, Piantelli M, et al. PLoS One. 2011;6(3): e17971.
- Kilis-Pstrusinska K. Carnosine and Kidney Diseases: What We Currently Know? Curr Med Chem. 2019.
- Albrecht T, Schilperoort M, Zhang S, Braun JD, Qiu J, Rodriguez A, et al. Carnosine Attenuates the Development of both Type 2 Diabetes and Diabetic Nephropathy in BTBR ob/ob Mice. Sci Rep. 2017;7: 44492.
- Peters V, Klessens CQ, Baelde HJ, Singler B, Veraar KA, Zutinic A, et al. Intrinsic carnosine metabolism in the human kidney. Amino Acids. 2015;47(12): 2541-2550.
- Zhao J, Posa DK, Kumar V, Hoetker D, Kumar A, Ganesan S, et al. Carnosine protects cardiac myocytes against lipid peroxidation products. Amino Acids. 2019;51(1): 123-138.
- de Courten B, Kurdiova T, de Courten MP, Belan V, Everaert I, Vician M, et al. Muscle carnosine is associated with cardiometabolic risk factors in humans. PLoS One. 2015;10(10): e0138707.
- Davis CK, Laud PJ, Bahor Z, Rajanikant GK, Majid A. Systematic review and stratified meta-analysis of the efficacy of carnosine in animal models of ischemic stroke. J Cereb Blood Flow Metab. 2016;36(10): 1686-1694.
- Baek SH, Noh AR, Kim KA, Akram M, Shin YJ, Kim ES, et al. Modulation of mitochondrial function and autophagy mediates carnosine neuroprotection against ischemic brain damage. Stroke. 2014;45(8): 2438-2443.
- Rothan HA, Abdulrahman AY, Khazali AS, Nor Rashid N, Chong TT, Yusof R. Carnosine exhibits significant antiviral activity against Dengue and Zika virus. J Pept Sci. 2019;25(8): e3196.
- Nagai K, Suda T. Antineoplastic effects of carnosine and beta-alanine--physiological considerations of its antineoplastic effects. Nihon Seirigaku Zasshi. 1986;48(11): 741-747.
- Holliday R, McFarland GA. Inhibition of the growth of transformed and neoplastic cells by the dipeptide carnosine. Br J Cancer. 1996;73(8): 966-967.
- Renner C, Asperger A, Seyffarth A, Meixensberger J, Gebhardt R, Gaunitz F. Carnosine inhibits ATP production in cells from malignant glioma. Neurol Res. 2010;32(1): 101-105.
- Shen Y, Yang J, Li J, Shi X, Ouyang L, Tian Y, et al. Carnosine inhibits the proliferation of human gastric cancer SGC-7901 cells through both of the mitochondrial respiration and glycolysis pathways. PLoS One. 2014;9(8): e104632.
- Hipkiss AR, Gaunitz F. Inhibition of tumour cell growth by carnosine: Some possible mechanisms. Amino Acids. 2014;46(2): 327-337.
- Iovine B, Iannella ML, Nocella F, Pricolo MR, Bevilacqua MA. Carnosine inhibits KRAS-mediated HCT116 proliferation by affecting ATP and ROS production. Cancer Lett. 2012;315(2): 122-128.
- Cheng JY, Yang JB, Liu Y, Xu M, Huang YY, Zhang JJ, et al. Profiling and targeting of cellular mitochondrial bioenergetics: inhibition of human gastric cancer cell growth by carnosine. Acta Pharmacol Sin. 2019;40(7): 938-948.
- Sugihara Y, Onoue S, Tashiro K, Sato M, Hasegawa T, Katakura Y. Carnosine induces intestinal cells to secrete exosomes that activate neuronal cells. PLoS One. 2019;14(5): e0217394.
- Hipkiss AR, Chana H. Carnosine protects proteins against methylglyoxal-mediated modifications. Biochem Biophys Res Commun. 1998;248(1): 28-32.
- da Silva Bispo V, Di Mascio P, Medeiros M. Quantification of Carnosine-Aldehyde Adducts in Human Urine. Free Radic Biol Med. 2014;75 Suppl 1: S27.
- Carvalho VH, Oliveira AHS, de Oliveira LF, da Silva RP, Di Mascio P, Gualano B, et al. Medeiros MHG Exercise and β-alanine supplementation on carnosine-acrolein adduct in skeletal muscle. Redox Biol. 2018;18: 222-228.
- Baba SP, Hoetker JD, Merchant M, Klein JB, Cai J, Barski OA, et al. Role of aldose reductase in the metabolism and detoxification of carnosine-acrolein conjugates. J Biol Chem. 2013;288(39): 28163-28179.
- Gęgotek A, Skrzydlewska E. Biological effect of protein modifications by lipid peroxidation products. Chem Phys Lipids. 2019;221: 46-52.
- Schumacher D, Morgenstern J, Oguchi Y, Volk N, Kopf S, Groener JB, et al. Compensatory mechanisms for methylglyoxal detoxification in experimental & clinical diabetes. Mol Metab. 2018;18: 143-152.
- Ohnuma T, Nishimon S, Takeda M, Sannohe T, Katsuta N, Arai H. Carbonyl Stress and Microinflammation-Related Molecules as Potential Biomarkers in Schizophrenia. Front Psychiatry. 2018;9: 82.
- Chengappa KN, Turkin SR, DeSanti S, Bowie CR, Brar JS, Schlicht PJ, et al. A preliminary, randomized, double-blind, placebo-controlled trial of L-carnosine to improve cognition in schizophrenia. Schizophr Res. 2012;142(1): 145-152.
- Ghajar A, Khoaie-Ardakani MR, Shahmoradi Z, Alavi AR, Afarideh M, Shalbafan MR, et al. L-carnosine as an add-on to risperidone for treatment of negative symptoms in patients with stable schizophrenia: A double-blind, randomized placebo-controlled trial. Psychiatry Res. 2018;262: 94-101.
- Zuccoli GS, Guest PC, Martins-de-Souza D. Effects on Glial Cell Glycolysis in Schizophrenia: An Advanced Aging Phenotype? Adv Exp Med Biol.2019;1178: 25-38.
- Cartwright SP, Bill RM, Hipkiss AR. L-carnosine affects the growth of Saccharomyces cerevisiae in a metabolism-dependent manner. PLoS One. 2012;7(9): e45006.
- Ikeda T, Kimura K, Morioka S, Tamaki N. Inhibitory effects of Zn2+ on muscle glycolysis and their reversal by histidine. J Nutr Sci Vitaminol (Tokyo). 1980;26(4): 357-366.
- Zhang Z, Miao L, Wu X, Liu G, Peng Y, Xin X, et al. Carnosine inhibits the proliferation of human gastric carcinoma cells by retarding Akt/mTOR/p70S6K Signaling. J Cancer. 2014;5(5): 382-389.
- Zhao J, Zhai B, Gygi SP, Goldberg AL. mTOR inhibition activates overall protein degradation by the ubiquitin proteasome system as well as by autophagy. Proc Natl Acad Sci USA. 2015;112(52): 15790-15797.
- Hipkiss AR, Brownson C, Bertani MF, Ruiz E, Ferro A. Reaction of carnosine with aged proteins: Another protective process? Ann N Y Acad Sci. 2002;959: 285-294.
- Oppermann H, Faust H, Yamanishi U, Meixensberger J, Gaunitz F. Carnosine inhibits glioblastoma growth independent from PI3K/Akt/mTOR signaling. PLoS One. 2019;14(6): e0218972.
- Oppermann H, Alvanos A, Seidel C, Meixensberger J, Gaunitz F. Carnosine influences transcription via epigenetic regulation as demonstrated by enhanced histone acetylation of the pyruvate dehydrogenase kinase 4 promoter in glioblastoma cells. Amino Acids. 20119;51(1): 61-71.
- Xu W, Li L, Sun J, Zhu S, Yan Z, Gao L, et al. Putrescine delays postovulatory aging of mouse oocytes by upregulating PDK4 expression and improving mitochondrial activity. Aging (Albany NY). 2018;10(12): 4093-4106.
- McFarland GA, Holliday R. Retardation of the senescence of cultured human diploid fibroblasts by carnosine. Exp Cell Res. 1994;212(2): 167-175.
- Chaleckis R, Murakami I, Takada J, Kondoh H, Yanagida M. Individual variability in human blood metabolites identifies age-related differences. Proc Natl Acad Sci USA. 2016;113(16): 4252-4259.
- Hipkiss AR. Could carnosine or related structures suppress Alzheimer's disease? J Alzheimers Dis. 2007;1(2): 229-240.
- Boldyrev A, Fedorova T, Stepanova M, Dobrotvorskaya I, Kozlova E, Boldanova N, et al. Carnosine [corrected] increases efficiency of DOPA therapy of Parkinson's disease: a pilot study. Rejuvenation Res. 2008;11(4): 821-827.
- Bermúdez ML, Seroogy KB, Genter MB. Evaluation of Carnosine Intervention in the Thy1-aSyn Mouse Model of Parkinson's Disease. Neuroscience. 2019;411: 270-278.
- Hipkiss AR. Glycotoxins: Dietary and Metabolic Origins; Possible Amelioration of Neurotoxicity by Carnosine, with Special Reference to Parkinson's Disease. Neurotox Res. 2018;34(1): 164-172.
- Zhao J, Shi L, Zhang LR. Neuroprotective effect of carnosine against salsolinol-induced Parkinson's disease. Exp Ther Med. 2017;14(1): 664-670.
- Hipkiss AR. Could carnosine suppress zinc-mediated proteasome inhibition and neurodegeneration? Therapeutic potential of a non-toxic but non-patentable dipeptide. Biogerontology. 2005;6(2): 147-149.
- Bermúdez ML, Skelton MR, Genter MB. Intranasal carnosine attenuates transcriptomic alterations and improves mitochondrial function in the Thy1-aSyn mouse model of Parkinson's disease. Mol Genet Metab. 20198;125(3): 305-313.
- Yu Q, Guo P, Li D, Zuo L, Lian T, Yu S, Hu Y, et al. Olfactory dysfunction and its relationship with clinical symptoms of Alzheimer Disease. Aging Dis. 2018;9(6): 1084-1095.
- Qin Z, Dimitrijevic A, Aswad DW. Accelerated protein damage in brains of PIMT+/-mice; a possible model for the variability of cognitive decline in human aging. Neurobiol Aging. 2015;36(2): 1029-1036.
Citation: Hipkiss AR (2019) Vulnerability of Human Erythrocytes to Persistent High Glycemic Index Diets: Implications for Ageing and Neurodegeneration: Possible Amelioration by Carnosine. J Aging Sci. 7:214. DOI:10.35248/2329-8847.19.07.214
Copyright: © 2019 Hipkiss AR. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.