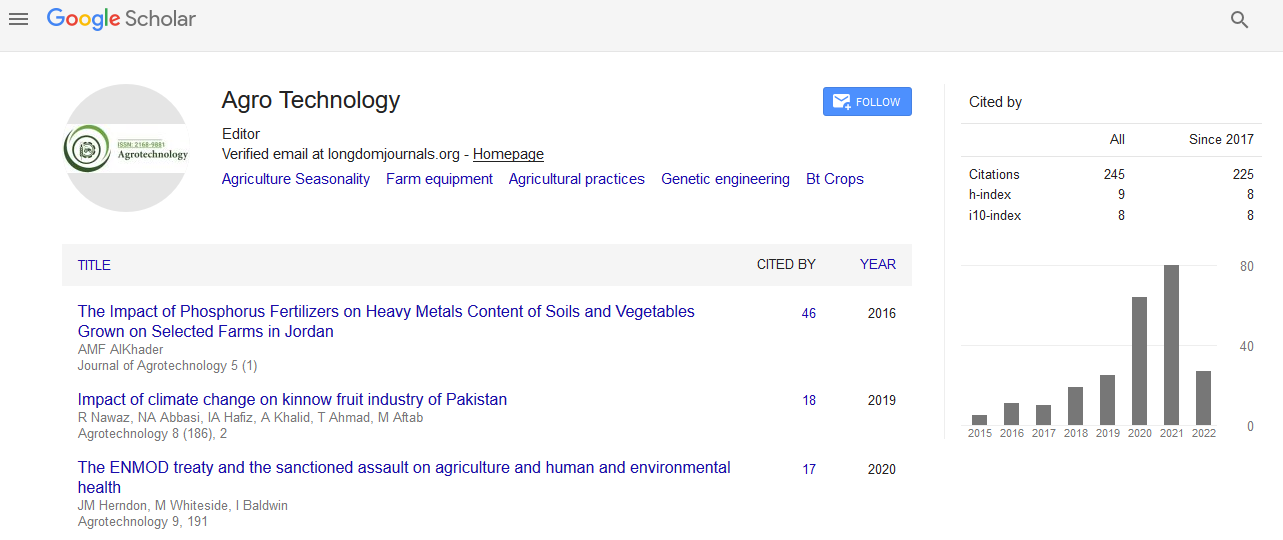

Indexed In
- Open J Gate
- Genamics JournalSeek
- Academic Keys
- ResearchBible
- Cosmos IF
- Access to Global Online Research in Agriculture (AGORA)
- Electronic Journals Library
- RefSeek
- Directory of Research Journal Indexing (DRJI)
- Hamdard University
- EBSCO A-Z
- OCLC- WorldCat
- Scholarsteer
- SWB online catalog
- Virtual Library of Biology (vifabio)
- Publons
- Geneva Foundation for Medical Education and Research
- Euro Pub
- Google Scholar
Useful Links
Share This Page
Journal Flyer

Open Access Journals
- Agri and Aquaculture
- Biochemistry
- Bioinformatics & Systems Biology
- Business & Management
- Chemistry
- Clinical Sciences
- Engineering
- Food & Nutrition
- General Science
- Genetics & Molecular Biology
- Immunology & Microbiology
- Medical Sciences
- Neuroscience & Psychology
- Nursing & Health Care
- Pharmaceutical Sciences
Review Article - (2020) Volume 9, Issue 1
Northern Treelines as Indicators of Climate and Land Use Changes - A Literature Review
Oddvar Skre*Received: 28-Jul-2019 Published: 10-Sep-2019, DOI: 10.35248/2168-9881.19.8.190
Abstract
The alpine and arctic treelines are results of long-term processes involving adaptation to a cold climate with environmental stress, and with low soil temperatures and nutrient uptake rates. Global warming and changed land use as a result of changes in global economy and agricultural policy may interfere with the treeline changes. The global warming is expected to enhance tree growth and seed reproduction, thereby increasing treeline altitudes and latitudes. Expansion of the forested area as a result of reduced logging and grazing would also reduce the albedo and enhance global warming in treeline areas. Feedback effects caused by increased soil temperatures and related output of greenhouse gases is expected to further increase global warming and treeline advance. On the other hand, local disturbance factors like increased risks of insect outbreaks, windthrow, grazing, anthropogenic disturbance and paludification would reduce or interfere with these changes, or even lead to a retreat of treelines. These limitations have to be taken into account when evaluating treelines as climatic indicators.
Keywords
Treelines; Temperature; Land use; Seed reproduction; Feedbacks; Human interactions; Local disturbance factors
Introduction
The circumpolar treelines and corresponding forest lines between the northern boreal forest, or taiga and the treeless tundra is probably the most important vegetation transition on the northern hemisphere. Paleobotanical studies indicate that the subarctic and subalpine tundra and the transition zone between the tundra and boreal forest in the past have been unstable ecosystems that are likely to be strongly affected by future climate and land use changes [1,2]. According to Aas [3] and Aas and Faarlund [4,5] a forest is defined as a stand of more than 30 trees where the distance between individual trees is <15 meter. A tree is defined here as an individual of >2.5 meter height, while e.g. Kullman [6] preferred a minimum tree height in birch of 2 meter, and is supported by Holtmeier [2] and Körner [7]. The forest line or timberline (Figure 1) is mainly a seed maturation limit, where viable seeds may be dispersed from their mother trees from below the forest line and develop into trees further upward and northward, but restricted by the arctic and alpine treelines (Figure 1) where vegetative growth is the limiting factor [8-10]. The definition and relationship between treelines and forest lines is illustrated in Figure 1. According to Kullman [6] tree height is strongly influenced also by the depth of the snow cover, and for this reason a woody plant individual should be defined as a tree already at 2 meter height or less. Mountain and tree-line areas exhibit part of the highest natural (genetic) as well as cultural diversity. Many species are strongly adapted to a harsh environment and seasonal stress, and they are vulnerable to disturbances and fast environmental changes [11]. Specific changes in transition zones (ecotones) therefore depend on the frequency, impact and duration of these environmental changes. However, as a result of the recent global warming and changed land use practise, an increase in circumpolar treeline altitudes and latitudes has already been observed (e.g. from the Canadian Subarctic [12- 15], from Fennoscandia [16,17] and from the Russian Subarctic [18-21]. The latitudinal treelines are usually much more influenced by local factor, e.g. topography and local climate, than altitudinal treelines [15,17,22]. A number of review articles have also been published on this topic, e.g. Grace et al. [23], Skre et al. [1], Payette et al. [24], Holtmeier [25], Holtmeier and Broll [26,27], Bryn and Pothoff [28], Cudlin et al. [29]. Based on these results, it may be hypothesized that changes in treeline position to a certain degree may be used as indicators of global warming on a regional scale [30]. These ecosystems are sensitive to climate changes in many ways, such as higher temperatures and longer growing seasons, moving treelines north- and upwards while fast-growing and competitive species and ecotypes are replacing counterparts adapted to the cold. However, the speed of the treeline advance is to a high degree dependent on the growth rates of seedlings and the length of the juvenile period [31].

Figure 1: Profile through one of the Finnish Treeline Monitoring sites (Riisitunturi) at 66°N with spruce as dominant tree species (cf. Skre and Laine [37]). The profile is extending from the spruce forest (250 m) to the summit (470 m), just above the treeline. The seed maturation limits in terms of mean temperatures (tetratherms) during the four warmest months a year (t4) are shown for mountain birch (Betula pubescens ssp. czerepanovii), Norway spruce (Picea abies) and Scots pine (Pinus sylvestris), according to Kujala (1927), Mork (1933) and Aas [3].
The treeline is also affected by land use changes, and in fact, forest cover in Europe has increased over recent decades as a result of abandonment of farmland, influenced by the complex interactions within the coupled human – environment system. Holtmeier and Broll [32] as well as Ameztegui et al. [33], Bryn [34] and Rössler et al. [35] stressed the importance of reforestation or regrowth after abandonment of grazing and logging as an important driving force of treeline change. The related transitional ecosystems (i.e. low-alpine tundra ecosystem and subalpine forests) would rapidly degrade unless best-management practices, conservation and preventative measures are promoted. Such changes also affect biodiversity and human uses, and understandings of mountain and treeline ecosystems and landscapes. Treelines are also important for biodiversity and ecosystem services, soil formation and slope stability, water retention and quality and as habitats and corridors for many species [36].
Climate and Growth
Reviews of earlier papers concerning limiting processes for growth in northern areas indicates that winter as well as summer temperatures are important controlling factors, through their effects on growth, nutrient uptake rates and seed production [37,38]. Although the alpine tree- and forest lines are considered sensitive indicators of vegetation response to climate change [25,36] and other disturbances, there is not a generally accepted single hypothesis that explains tree- or forest line position and pattern at multiple scales [7,39].
Growth limitation hypothesis
Recently a set of differing hypotheses has been proposed for tree- and forest line explanations [7,30], separating the drivers of tree line formation into two categories: modifying (regional) and fundamental (global). These include the stress hypothesis, the disturbance hypothesis, the reproduction hypothesis, the carbon balance hypothesis, and, finally, the growth hypothesis, and they are mainly driven by climatic changes. The reproduction, stress and growth hypothesis may be combined into three main factors [37].
Reproduction limitation: Low air and soil temperatures, retarded shoot growth and too short growing season for seed and bud development [40,41].
Growth limitation: Low soil temperatures that are lowering the decomposition rates, microbial respiration and root respiration (Figure 2), thereby decreasing root growth and nutrient uptake [42,43].

Figure 2: Root nitrogen uptake rates for mountain birch seedlings grown under experimental conditions for one season in Abisko, northern Sweden. Two fertilizer levels were used, Low (25 uM with respect to N) and High (100 uM). Seedlings grown in organic soil (Org.) were supplied with distilled water only. Redrawn from Karlsson and Nordell [43].
Stress limitation: Frozen soil combined with radiation and strong winds in early spring leading to damages caused by freeze drying, i.e. chlorophyll breakdown [44].
Hustich (1979) [9] combined all these factors into the “climate hazard” coefficient to indicate northernness in the stress to which the treeline forests are subjected. However, comparative studies e.g. by Skre and Laine [37], Karlsson and Nordell (1996) [43] and Körner (2012) [7], see also Chapin [42] indicate that the most important limiting process for vegetative growth at treeline probably is the active uptake of nutrients (P, N) in roots, i.e. the growth hypothesis. This process, as well as the decomposition rates of nutrients in soil by microorganisms, has been shown to be strongly temperaturedependent (Figure 2). Winter stress and seed and bud development rates are modifying factors, where e.g. high seed reproduction rates would increase the chances of seedling survival, and winter conditions would influence on the survival. The mentioned studies therefore seem to support the “climate hazard” hypothesis [1], where all three mentioned factors are involved, but where the growth hypothesis is the most important.
Climatic limitation and adaptation
When looking at the northern (arctic) circumpolar treeline, it may be considered as a result of a long-term adaptation to ecological and environmental factors (drivers). At a large scale, the arctic treeline seems to be mainly a result of the “climate hazard” factor [11] including seed reproduction and freezing stress. Lack of propagules (viable seeds and/or buds) is a probable reason for the abscense or low density of trees at the tundra/taiga boundary. However, individual clones of trees, e.g. Norway spruce (Picea abies) or black spruce (P. mariana) may survive as vegetative “krummholz” individuals, by producing daughter individuals from layerings for several hundred years, and eventually produce viable seeds as soon as summer temperatures are high enough for seed reproduction [24]. These seeds may eventually lead to reforestation of treeline areas (Figure 1) and increased treeline altitudes. In many cases there are more favorable sites for seedling establishment around the treeline, e.g. better light and nutrient conditions [11,45,46] than further down in the forest, therefore the proportion of seedlings relative to the total number of trees are highest at or above treeline [16]. Annual growth decreases with altitude across the treeline (Figure 2) as a result of reduced summer temperature and shorter growing season. However, because of slow growth it may take many years or even decades before new seedlings develop into trees. Trees with long juvenile periods expand their ranges slowly, and community level changes may be delayed for many decades [11,31,47-49].
Feedback effects
The low temperature and short growing season at the treeline is closely correlated with tree performance. However, the presence of trees and shrubs is also in itself creating favorable sites for tree establishment. Treeline studies have documented that both upright and krummholz individuals of trees have large effects on wind dynamics and snow distribution [50-52]. In other words, the effect of trees on their environment may act as a feedback that enhances conditions for other trees, and their own longevity [53]. Because of reduced wind speed inside a tree canopy, there will be less wind-induced needle damage and needle loss when other trees are present [54]. Trees also decrease the albedo and canopy roughness, thereby further increasing needle surface temperatures [55]. However, increased competiton for nutrients may reduce the positive feedbacks from a close canopy of trees, where often also soil temperatures are lower than in adjacent unforested areas outside the canopy [56]. Therefore, the explanations for reduced tree performance across treelines have to consider the often densitydependent influence of trees on their environment.
Another feedback occurs when trees move northward and/or upward as a result of global warming. Ecosystem models [57,58] predict a significant reduction of the tundra biome with large parts being transformed into forests, especially in northern Eurasia. This conclusion has been confirmed by other recent studies, e.g. by Epstein et al. [59] using the BIOME4 and ArcVeg models to predict the consequences of a +2 degrees temperature increase on the circumpolar vegetation limits. They found a 55% increase in the forested area up to 2060 – equivalent to 3 millions square kilometers – and a corresponding 42% decrease in the Arctic tundra, mostly in Eurasia. The models alsi predicted a succession with time lags ranging from 100-150 years for tall shrubs and considerably shorter time for mosses and prostrate shrubs. Other studies based on space-for-time substitutions and past changes of temperature and moisture are pointing in the same direction [60,61]. The lowered albedo as a result of forest expansion, and altered microclimate and hydrology may lead to increased soil temperatures and decomposition rates and increased output of greenhouse gases, which in turn is expected to lead to a feedback effect on global warming [62]. However, thickening of sparse treeline forests are also often resulting in more closed canopies and soil cooling at treeline, with consequences for decomposition rates and nutrient status [63,64]. These processes are also expected to influence strongly on the position of arctic treelines, i.e. the tundra/taiga boundary [65]. The northward tree migration may be hampered by the development of peatlands, high permafrost levels and unfavorable soil conditions. This development has already taken place during late Holocene [1]. The global warming, however, will probably lead to widespread melting of permafrost and release of trace gases that are expected to increase global warming, soil respiration and decomposition rates by feedback effects [66].
Interactions between climate and Land-use
When evaluating treelines as indicators of climate change through temperature limitations, there are a number of modifying factors that has to be taken into account. Temperature changes strongly affect soil decomposition rates and water level, and influence on the radiation balance (albedo) by altering the positon of treelines, i.e. the treeline in itself may have a feedback effect on global warming [65,66]. Higher seed reproduction as a result of increased summer temperatures [16] and lower grazing pressure is expected to lead to an advance of the treeline ecotone [67], but the process may take decades or even centuries [1,68]. According to de Wit et al. [69], who modeled forest expansion for an estimated 2.6 degrees increase in summer temperature in 2100, birch forest cover in south-central Norway in the abscense of land-use, is expected to increase from 12 to 27% during the period from 2000 to 2100, resulting in a 59% increase in biomass carbon storage and a change in albedo from 0.46 to 0.30. The projected modeling study showed that the warming caused by this lowered albedo from expanding forests was 10-17 times stronger than the cooling effect from carbon sequestration, including all emission types. Reduced snow cover further would amplifiy the feedback effect of lower albedo. It may be noticed however, that the current (2000) forest line in this area is about 100 meter lower than its climatic potential due to landuse history. Climatic changes also affect local disturbance factors like insect and fungus attacks, forest fires and windthrows, or a combination of these factors [11,70]. Finally, the human impact through logging, grazing, roadbuilding and mass tourism may interact with climate changes on a local and regional scale.
Human interactions
Recently much attention has been paid to the expected global warming as a result of increased output of greenhouse gases. However, a substantial part of the observed rise in treeline altitudes [4] is probably caused by a combination of global warming and changed land use, e.g. reforestation after abandonment of grazing [71-74]. In fact, recent studies indicate that most of the treeline rise registered is caused by regrowth [33,34,69,75]. The changed land use may partly be a result of changes in global economy and agricultural policy [74]. Mountain ecosystems are a significant reservoir of biodiversity in terrestrial ecosystems, partly because the range of environmental variation and ecological gradients facilitate great diversity of organisms, and human intervention in mountain ecosystems is less intense than in lowlands [76]. The retreat of the tundra/taiga transition zone towards higher altitudes is related to an increased risk of extinction of high alpine species, and a significant soil carbon release [77]. In fragmented landscapes species that have intrinsically low population growth rates and short-range dispersal may be vulnerable to extinction and slowcolonizing species have much lower patch occupancy than do rapid colonizers [78]. Distance to reservoirs of propagules probably plays an important role on the colonization of new areas and on the recovery dynamic and soil factors [79-81]. There is probably also increased chance of introduction of invasive species (animals and plants), which alter the competition, and increased risks of diseases and disturbance. Genes may be transferred between different populations, and give rise to new ecotypes and races that are better adapted to the environment than their origin. On the other hand, global warming may also increase the frequency and intensity of insect outbreaks [70], which could have dramatic effects. In addition to temperature conditions, the recovery potential and re-generation rates of different plant species depend on grazing pressure and plant vitality, in particular on the survival rates of young growth.
In some regions, changes in the economic market environment have evoked changes in the traditional land use of mountain regions. Agricultural expansion into marginal environments, deforestation, recreation and mass tourism can lead to vegetation and soil changes, changes in water balance and quality, erosion, disturbance of animals and reduced visual and aesthetic value of the landscape [82]. On the other hand, advance of subalpine and boreal forests into tundra and alpine regions is also enhanced by the abandonment of summer grazing [34,75], in consequence of common agricultural policies [71]. Natural and traditionally managed ecosystems are expected to be rapidly degraded unless best-management practices are carried out, conservation and preventive measures are enforced. In northern Russia, however, heavy industrialization in the lesotundra transition zone as well as increased logging and grazing pressure during the last decades have caused a retreat of the circumpolar treeline in some areas [82,83]. This retreat is often connected to a combination of massive output of sulfur dioxide from mining industry, cooling as a result of deforestation and solid particle emission, and termokarst formation as a result of water logging caused by permafrost melting. Deforestation from increased logging, mining and oil exploration activities has in turn reduced the reindeer grazing areas and caused overgrazing in the remaining areas [84]. In addition climate-related factors like fire and insect outbreaks (see Chapter 3.2), interact with the land use changes, and the overall consequences are breakdown of the locally-based economy and sustainability, and reduced quality of life [82].
Local disturbance factors
Environmental fragmentation of mountain areas on a local scale may be caused by a number of disturbances, both of natural and anthropogenic origin. Examples of the first category are provided by herbivores like insects or ungulates, and of the latter locally generated by tourism exploitation like ski resort enterprises, by air pollution, forest logging, mineral exploitation and over-grazing by reindeer and sheep. Grazing by sheep and reindeer has formed the tree line as well as the forest line in northern Norway [71,85,86], and animals or insects may have modifying effects on treelines in a variety of environments [39]. Insect herbivory that causes the death of the trees is the most widely reported mode of animal activity, and the effect of the autumnal moth (Epirrita autumnata) on birch (Betula pubescens) has been widely studied in northern Sweden and Finland. The Epirrita eggs will not survive in the winter buds at temperatures below –35˚C [70], hence the global warming may have increased their survival rates. Further, after the Epirrita attack in northern Finland in 1965-66 the birch were not able to recover because of heavy reindeer grazing, as a result a permanent lowering of the treeline has taken place in the Utsjoki area [87-89]. Wind may also be an important disturbance factor locally, by causing abrasion damages on needles and braches of trees above the snow surface, particularly on the windward side of the tree [11,44,90]. In some areas long term wind damages have resulted in ribbon- or wave-shaped forests that gradually change their position in a regular manner [91]. Another example, forest fires or fires in the mountain heaths, may be naturally occurring and/or man-made. In some parts of northern Eurasia, Canada and Alaska, these and other disturbances are considered to be the main drivers in controlling the properties of the forest-mountain or the forest-tundra transition zone.
Conclusions
When evaluating the observed changes in treeline position on a altitudinal and latitudinal scale, the importance of non-climatic drivers like disturbance and land use changes should always be thoroughly analysed, in order to estimate the effects of global warming, and feedback effects due to changes in light, nutrient content, paludification and soil warming, should be investigated. Global warming may influence tree growth and treeline position through increased seed viability and production, as well as increased vegetative growth, bud survival and reproduction. The first indications of these changes are therefore often an increase in seedlings density at or above treeline, as observed by Juntunen and Neuvonen [16] in northern Finland.
However, also the effects of seedling mortality as a result of freezing and drought stress, insect and fungus attacks, should be considered, as well as the effects of grazing pressure [86,88]. The final outcome is dependent on these factors and to the degree of human interaction e.g. by increased or reduced cutting, grazing, or by other disturbance factors. With these limitations taken into account, changes in treeline position may to a certain extent be used as a rough indication of the relative importance of climate vs. land use changes. In these considerations the local as well as global economy and its consequences for the agricultural policy have to be taken into account.
REFERENCES
- Skre O, Baxter R, Crawford RMM, Callaghan TV, Fedorkov A. How will the Tundra-Taiga interface respond to climate change? Ambio Special Report. 2002;12:37-46.
- Holtmeier FK. Mountain timberlines. Ecology, patchiness and dynamics. Berlin, Springer. 2009;436.
- Aas B. Bjørke- og barskoggrenser i Norge. Master Thesis. University of Oslo. 1964.
- Aas B. Climatically raised birch lines in Southeastern Norway 1918-1968. Norsk geogr Tidsskr. 1969;23:119-130.
- Aas B, Faarlund T. The present and the holocene birch belt in Norway. Paleoklimaforschung. 1996;20:18-24.
- Kullman L. A decade of treeline monitoring in the southern Swedish Scandes. UNGI Report. 1987;65:191-202.
- Körner C. Alpine treelines. Functional Ecology of the Global High Elevation Tree Limits. Berlin-Springer, Basel, 2012.
- Dahl E, Mork E. Om sambandet mellom temperatur, ånding og vekst hos gran (Picea abies (L.) Karst Meddr norske SkogforsVes. 1959;16:81-93.
- Hustich I. Ecological concepts and biological zonation in the North; the need for a general accepted terminology. Holarct Ecol 1979;2:208-217.
- Skre O. Physiology of plant survival under cold conditions, with particular reference to dark respiration as a factor limiting growth at timberline. A literature review. Medd Skogforsk. 1991;44:1-34.
- Sveinbjörnsson B, Hofgaard A, Lloyd A. Natural causes of the Tundra-Taiga boundary. Ambio Special Report. 2002;12:23-29.
- Luckman BH, Kavanagh TA. Impact of Climate Fluctuations on Mountain Environments in the Canadian Rockies. Ambio. 2000;29(7):371-380.
- Lescop-Sinclair K, Payette S. Recent advance of the arctic treeline along the eastern coast of Hudson Bay. J Ecol. 1995;83:929-936.
- Gamache I, Payette S. Latitudinal response of subarctic tree lines to recent climate change in eastern Canada. J Biogeogr. 2005;32(5):849-862.
- Payette S, Fortin MJ, Gamache I. The subarctic forest-tundra: the structure of a biome in a changing climate. BioScience. 2001;51:709-718.
- Juntunen V, Neuvonen S. Natural regeneration of Scots pine and Norway spruce close to the timberline in northern Finland. Silva Fennica. 2006;40(3):443-458.
- Hofgaard A, Tømmervik H, Rees G, Hanssen F. Latitudinal forest advance in northernmost Norway since the early 20th century. J Biogeogr. 2013;40:938-949.
- Esper J, Schweingruber FH. Large-scale treeline changes recorded in Siberia. Geophys Res Letters. 2004;31:1-5.
- Kharuk VI, Ranson KJ, Im ST, Vdovin AS. Spatial distribution and temporal dynamics of high-elevation forest stands in southern Siberia. Global ecology and Biogeography. 2010;19:822-830.
- Koshkina NB, Moiseev PA, Goryaeva AV. Reproduction of the Siberian spruce in the timberline ecotone of the Iremel massif. Russian J Ecol. 2008;39(2):83-91.
- Moiseev PA, Bartysh AA, Nagimov Z.Ya. Climate changes and tree stand dynmics at the upper limit of their growth in the North Ural mountains. Russian Journal of Ecology. 2010;41(6):486-497.
- Harsch MA, Bader MY. Treeline form – a potential key to understanding treeline dynamics. Glob Ecol Biogrogr. 2011;20:582-596.
- Grace J, Berninger F, Nagy L. Impacts of Climate Change on the Tree Line. Ann Bot. 2002;90(4):537-544.
- Payette S, Eronen M, Jasinski JJP. The circumboreal Tundra-Taiga interface: Late Pleistocene changes. Ambio Special Report. 2002;12:15-22.
- Holtmeier FK. Waldgrenzstudies im nördlichsten Finnisch-Lapland und angrenzenden Nord-Nordwegen. Rep Kevo Subarct Res Sta. 1971;8:53-62.
- Holtmeier FK, Broll G. Treeline advance – driving processes and adverse factors. Landscape Online. 2007;1:1-32.
- Holtmeier FK, Broll G. Altitudinal and Polar Treelines in the Northern Hemisphere – Causes and Responses to Climate Change. Polarforsch. 2010;79(5):139-153.
- Bryn A, Pothoff K. Elevational treeline and forest line dynamics in Norwegian mountain areas – a review. Landscape Ecology. 2018;33(8):1225-1245.
- Cudlin P, Klopcic M, Tognetti R, Malis F, Alados CL, Bebi P, et al. Drivers of treeline and forest line dynamics in different European mountains. Climate Research. 2017;73:135-150.
- Smith WK, Germino MJ, Johnson DM, Reinhardt K. The altitude of alpine treeline; a bellweather of climate change effects. Bot Rev. 2009;75:163-190.
- Brubaker LB. Response of tree populations to climate change. Plant Ecology. 1986;67(2):119-130.
- Holtmeier FK, Broll G. Sensitivity and response of northern hemisphere altitudinal and polar treelines to environmental change at landscape and local scales. Global Ecol. Biogeogr. 2005;14:395-410.
- Ameztegui A, Coll L, Brotons L, Ninot JM. Land-use legacies rather than climate change are driving the recent upward shift of the mountain tree line in the Pyrennees. Glob Ecol. 2016;25:263-273
- Bryn A. Recent forest limit changes in south-east Noway: Effects of climate change or re-growth after abandoned utilization? Norwegian Journal of Geography. 2008;62(4):251-270.
- Rössler O, Bräuning A, Löffler J. Dynamics and driving forces of treeline fluctuation and regeneration in central Norway during the past decades. Erdkunde. 2008;62:117-128.
- Kullman L. Tree-limits and montane forests in the Swedish Scandes; Sensitive biomonitors of climate change and variability. Ambio. 1998;27:312-321.
- Skre O, Laine K. Limiting processes for growth in northern tree species. In: Taulavuori, K. & Taulavuori, E. (eds.): Physiology of Northern Plants under Changing Environments, Research Signpost, Kerala, India, 2007; pp. 211-227.
- Harsch MA, Hulme PE, McGlone MS, Duncan RP. Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecol Lett. 2009;12:1040-1049.
- Cairns DM, Lafon C, Moen J, Young A. Influence of animal activity on treeline position and pattern: Implication for treeline responses to climatic change. Physical Geography 2007;28:419-433.
- Crawford RMM. Studies in plant survival. Blackwell Sci Publ Oxford. 1989;296.
- Skre O. Growth of mountain birch (Betula pubescens Ehrh.) in response to changing temperatures. In: Alden J, Maastrantonio JL, Ødum S (eds.): Forest Development in Cold Climates, Plenum Press, N.Y. 1993;65-78.
- Chapin FS III. The mineral nutrition of wild plants. Ann Rev Ecol Syst. 1980;11:238-260.
- Karlsson PS, Nordell KO. Effects of soil temperature on nitrogen economy and growth of mountain birch near its presumed low temperature distribution limit. Ecoscience. 1996;3:182-189.
- Tranquillini W. Physiological ecology of the alpine timberline. Tree existence at high altitudes with special reference to the European Alps. Ecological Studies. 1979;137.
- Sullivan PF, Sveinbjörnsson B. Microtopographic control of treeline advance in Noatak National Preserve, northwest Alaska. Ecosystems. 2010;11:275-285.
- Lett S, Nilsson MC, Wardle DA, Dorrepaal E. Bryophye traits explain climate-warming effects on tree seedling establishment. J Ecol. 2016;105:496-506.
- Davis MB. Lags in vegetation response to greenhouse gas emissions. Earth and Environmental Science 1986;15(1-2):75-82.
- Malis F, Kopecky M, Petrik P, Vladovic J, Merganic J, Vida T. Glob Change Biol. 2016;22:1904-1014.
- Rannow S. Do shifting forest limits in south-west Norway keep up with climte change? Scand J For Res. 2013;28:574-580.
- Scott PA, Bentley CV, Fayle DCF, Hansell RI. Crown forms and shoot elongation of white spruce at the treeline. Arctic Alpine Res. 1987;19:175-189.
- Scott PA, Hansell IRC, Ericsson WR. Influence of wind and snow on northern treeline environments at Churchill, Manitoba, Canada. Arctic. 1993;46:316-323.
- Noble IR. A model of responses of ecotones to climate change. Ecol Applic. 1993;26:396-403.
- Kullman L. The geoecological history of Picea abies in northern Sweden and adjacent parts of Norway. A contrarian hypothesis of postglacial tree immigration patterns. Geoekol. 2000;21:141-172.
- Sveinbjörnsson B. North American and European treelines; External forces and internal processes controlling position. Ambio. 2000;29:388-395.
- Chapin FS III, Eugster W, McFadden JP, Lynch AH, Walker DA. Summer differences between Arctic ecosystems in regional climatic forcing. J Climate. 2000;13:2002-2010.
- Karlsson PS, Weih M. Soil temperatures near the distribution limits of the mountain birch; implications for seedling nitrogen economy and survival. Arctic Antarctic Alpine Res. 2001;33:88-92.
- Prentice IC, Cramer WP, Harrison SP, Leemans R, Monserud RA, Solomon AM. A global biome model based on plant physiology and dominance, soil properties and climate. J Biogeogr. 1992;19:17-134.
- Larsen JN, Anisimov OA, Constable A, Hollowed AB, Maynard N, Prestrud P, et al. Polar regions. In: Climate Change; Impacts, Adaptation and Vulnerability, Part B: Regional Aspects, Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel of Climate Change. Cambridge U. Press, Cambridge, N.Y. 2014;1567-1612.
- Epstein HE, Qin Yu, Kaplan JO, Lischke H. Simulating future changes in arctic and subarctic vegetation. Computing in Science and Engineering. 2007;9(4):12-23.
- Björkman A, Myers-Smith IH, Elmendorf SC, Normand S. Plant functional traits across a warming tundra. Nature. 2018;562.
- Beck PSA, Juday GP, Alix C, Barba VA, Winslow SE, Sousa EE, et al. Changes in forest productivity across Alaska consistent with biome shift. Ecology Letters. 2011;14(4):373-379.
- Rydsaa JH, Stordal F, Bryn A, Tallaksen LM. Effects of shrub and tree cover increase on the near-surface atmosphere in northern Fennoscandia. Biogeosciences. 2017;14:4209-4227.
- Haag RW, Bliss LC. Functional effects of vegetation on the radiant energy budget of boreal forests. Can Geotech J. 1974;11:374-379.
- Karlsson PS, Weih M. Soil temperatures near the distribution limits of the mountain birch; implications for seedling nitrogen economy and survival. Arctic Antarctic Alpine Res. 2001;33:88-92.
- Harding R, Kuhry P, Christensen TR, Sykes MT, Dankers R, van der Linden S. Climate feedbacks at the Tundra-Taiga Interface. Ambio Special Report. 2002;12:47-55.
- Crawford RMM, Jeffree CE, Rees WG. Paludification and forest retreat in northern oceanic environments. Ann Botany. 2002;91:213-226.
- Bryn A, Daugstad K. Summer farming in the subalpine birch forest. In: Wielgolaski FE (ed) Nordic mountain birch ecosystems. MAB Series. Parthenon Publ. Group, New York. 2001;307-315.
- Van Bogaert R, Haneca K, Hoogesteger J, Jonasson C, De Dapper M, Callaghan TV. A century of treeline changes in subarctic Sweden shows local and regional variability and only a minor influence of 20th century climate warming. J Biogeogr. 2011;38:907-921.
- De Wit H, Bryn A, Hofgaard A, Karstensen J, Kvalevåg M, Peters G. Climate warming feedback from mountain birch forest expansion; reduced albedo dominates carbon uptake. Global Change Biology 2014;20:2344-2355.
- Tenow O, Bylund H, Nilsen AC, Karlsson PS. Long-term infuence of herbivores on northern birch forests. In: Wielgolaski FE (ed.): Plant Ecology, Herbivory and Human Impact in Nordic Mountain Birch Forests. Ecological Studies. 2005;180:165-182.
- Dalen L, Hofgaard A. Differential regional treeline dynamics in the Scandes mountains. Arctic, Antarctic and Alpine Research. 2005;37:284-296.
- Aune S, Hofgaard EA, Søderstrøm L. Contrasting climate and land-use driven tree encroachment pattern of subarctic tundra in northern Norway and the Kola peninsula. Canadian Journal of Forest Research. 2011;41:437-450.
- Mathisen A, Mikheeva A, Tutubalina OV, Aune S, Hofgaard A. Fifty years of treeline change in the Khi biny Mountains, Russia; advantages of combined remote sensing and dendrochronological approaches. Applied Vegetation Sciences. 2014;17(1):6-16.
- Kyriazopoulos AP, Skre O, Sarkki S, Wielgolaski FE, Abraham EM, Ficko A. Human-environment dynamics in European treeline ecosystems; a synthesis based on the DPSIR framework. Climate Research. 2017;73:17-29.
- Gehrig-Fasel J, Guisan A, Zimmermann NE. Tree line shifts in the Swiss Alps. Climate change or land abandonment? Journal of Vegetation Science. 2007;18(4):571-582.
- Emanuelsson U. Human influence on the vegetation in the Torneträsk area during the last three centuries. Ecol Bull 1987;38:95-111.
- Sørensen MV, Strimbeck R, Nystuen RO, Kapas RE, Enquist B, Jessen B. Draining the pool? Carbon Storage and fluxes in three Alpine plant communities. Ecosystems. 2018;21(2):316-330.
- Collingham YC, Huntley B. Impact of habitat fragmentation and patch size upon migraton rates. Ecol Appl. 2000;10:131-144.
- Kullman L. The changing face of the alpine world. Global Change Newsletter. 2004;57:12-14.
- Kellomäki S, Kolstrom M. Simulation of tree species composition and organic matter accumulation in Finnish boreal forests under changing climatic conditions. Vegetatio 1992;102:47-68.
- Hobbie SE, Chapin FS III. An experimental test of limits to tree establishment in Arctic tundra. J Ecol. 1998;86:449-461.
- Vlassova T. Human Impact in the Tundra-Taiga Zone Dynamics: The Case of the Russian Lesotundra. Ambio Special Report. 2002;12:30-36.
- Orlova MA, Lukina NV, Tutubalina OV, Smirnov VE, Isaeva LG, Hofgard A. Soil nutrient’s spatial variability in forest-tundra ecotones on the Kola Peninsula, Russia. Biogeochemistry. 2013;113:283-305.
- Aksenov D, Karpatchevsky M, Lloyd S, Yaroshenko A. The Last of the Lasts. The Old-growth Forests of Boreal Europe. Taiga rescue network. 1999;67.
- Tømmervik H, Johansen B, Tombre I, Thannheiser D, Høgda KA, Gaare E, et al. Vegetation changes in the Nordic mountain birch forests; the influence of grazing and climate change. Arctic Antarctic Alpine Res. 2004;36:323-332.
- Speed JDM, Austrheim G, Hester AJ, Mysterud A. Experimental evidence for herbivore limitation of the treeline. Ecology. 2010;91:3414-3420.
- Lempa K, Neuvonen S, Tømmervik H. Effects of reindeer grazing on pastures in a mountain birch ecosystem. In: Wielgolaski, F.E. (ed.): Plant Ecology, Herbivory and Human Impact in Nordic Mountain Birch Forests, Ecological Studies, Springer. 2005;157-164.
- Helle T. Mountain birch forests and reindeer husbandry. In: Wielgolaski FE (ed): Nordic mountain birch ecosystems. MAB Series. Parthenon Publ. Group, New York, 2001:279-292.
- Anschlag K, Broll G, Holtmeier FK. Mountain birch seedlings in the treeline ecotone, subarctic Finland: Variation in above- and belowground growth depending on microtopography. Arctic, Antarctic and Alpine Res. 2008;40:609-616.
- Scott PA, Hansell IRC, Ericsson WR. Influence of wind and snow on northern treeline environments at Churchill, Manitoba, Canada. Arctic. 1993;46:316-323.
- Bekker M. Positive feedback between tree establishment and patterns of subalpine forest advancement, Glacier National Park, Montana, USA. Arctic, Antarctic and Alpine Res. 2005;37(1):97-107.
Citation: Skre O (2019) Northern Treelines as Indicators of Climate and Land Use Changes - A Literature Review. Agrotechnology 8:190.
Copyright: Ã?â??Ã?© 2019 Skre O. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.